
Abstract
Introduction
Recently, we showed that irradiation of EGCG-exposed HeLa cells with 670-nm laser light is instrumental in forcing the cells to rapidly uptake high amounts of EGCG and other cytostatic compounds. 11 The laser light was applied in a linear scanning mode at a frequency of 1 Hz with the basic irradiation parameters irradiance and fluence adjusted to the biostimularory windows 12 communicated by us earlier: 1·103 Wm2 and 1·104 Jm2, respectively. At 45 μM EGCG 1 min intermittent irradiation following an incubation period of 3 h resulted in a reduction in cell numbers by two-thirds compared to a reduction by one-third with EGCG alone. EGCG is described to kill tumor cells by telomere fragmentation. 13 In other words, EGCG, acting here as an anticancer compound, performed its function intracellularly. Interestingly, the anticancer effect is not restricted to the pure compound EGCG. As we previously demonstrated, even sterile-filtered green tea had a profound cytostatic effect on HeLa cells. 14 The function of the intermittent irradiation mode is to facilitate the transport of the active compound into cells via transmembrane convection, a mechanism based on the interaction of 670-nm light with nanoscopic interfacial water layers on surfaces. 15,16 The cause of the convective flow across the plasma membrane is the simultaneous modulation of the density and viscosity of the nanoscopic interfacial water layers prevailing between intracellular surfaces (macromolecules and organelles). The effect is expected to be most pronounced in cells that are rich in mitochondria. 17 Previously, we reported that the combination of topically applied EGCG and moderate levels of 670-nm light has beneficial effects in vivo, in particular in skin rejuvenation, where we exploited the capability of EGCG to act as a scavenger of reactive oxygen species (ROS). 18 We anticipated that irradiation with 670-nm light is possibly also useful in forcing brain cells to uptake EGCG, on the one hand, and to increase ATP levels in Aβ-stressed brain cells, on the other hand—both effects supposed to be beneficial in improving the condition of patients with AD. For clarification, we exposed human neuroblastoma (SH-EP) cells loaded with Aβ42 to 670-nm laser light and EGCG.
Material and Methods
Synthetic Aβ42 was labeled with the fluorescent dye Alexa 633-NHS (Invitrogen) overnight at 4°C at a 1:1 M ratio under the conditions specified by the manufacturer. Labeled peptide was purified by gel filtration in 10 mM NH4OH and filtered through a 30 kD membrane filter (Millipore) to remove aggregated material. SH-EP cells (25.000 cells per well) that express the green fluorescent protein (GFP)-tagged nuclear histone protein H2B, were cultured in DMEM supplemented with 4.5 g/L
To test the effects of EGCG treatment and laser irradiation on the clearance of aggregated intracellular Aβ42, we added fluorescently labelled Aβ42 to the culture medium containing the SH-EP cells. 3 Cells were then incubated for 24 h, before the culture medium was exchanged to remove Aβ42 from outside the cells. The cells were divided in six groups: (1) controls, (2) supplemented with 10 μM EGCG, (3) supplemented with 40 μM EGCG, (4) treated with Aβ42, (5) treated with Aβ42 and 10 μM EGCG, (6) treated with Aβ42 and 40 μM EGCG. Four wells of each group were subjected to laser treatment, whereas four wells remained untreated. Laser treatment (power 33 mW, beam geometry 2 mm x 15.5 mm, local irradiance 1·103 Wm2 and mean fluence 1·104 Jm2) was performed manually for 1 min per well (area 1.9 cm2) in a linear scanning mode at a frequency of 1 Hz (controlled by a metronome), according to the previously communicated protocol. 11 Cells were then incubated further for 24 h.
Next, the well plates were placed into a high-content screening (HCS) cell analysis system (Arrayscan VTI, Thermo Scientific). In groups 1 to 6 the analysis comprised cell proliferation and quantification of the Aβ42 aggregate amounts within the cells. The working principle of the HCS analysis is described in detail elsewhere. 19 Briefly, the compartmental analysis module of the ArrayScan VTI software was used to identify nuclei by GFP fluorescence, to fit cell dimensions and to identify Aβ42 aggregates by Alexa 633 fluorescence (cf. Fig. 1). Non-aggregate fluorescence background was discarded from the analysis and fluorescence signals were normalized to cell numbers. Four independent wells with ∼4500 cells per well were analyzed. Statistical significance was evaluated by Student's t test. * Marks p<0.05. Higher significance levels are marked by ** as indicated. All results are represented as mean values; error bars signify standard deviations. Furthermore, we checked for possible effects of the 670-nm laser light on the bleaching of the labeled Aβ42 aggregates. For this, we irradiated half of the previously nonirradiated wells using the protocol mentioned above, and quantified Aβ-Alexa 633 fluorescence signals directly after irradiation. Importantly, a bleaching effect could not be observed.
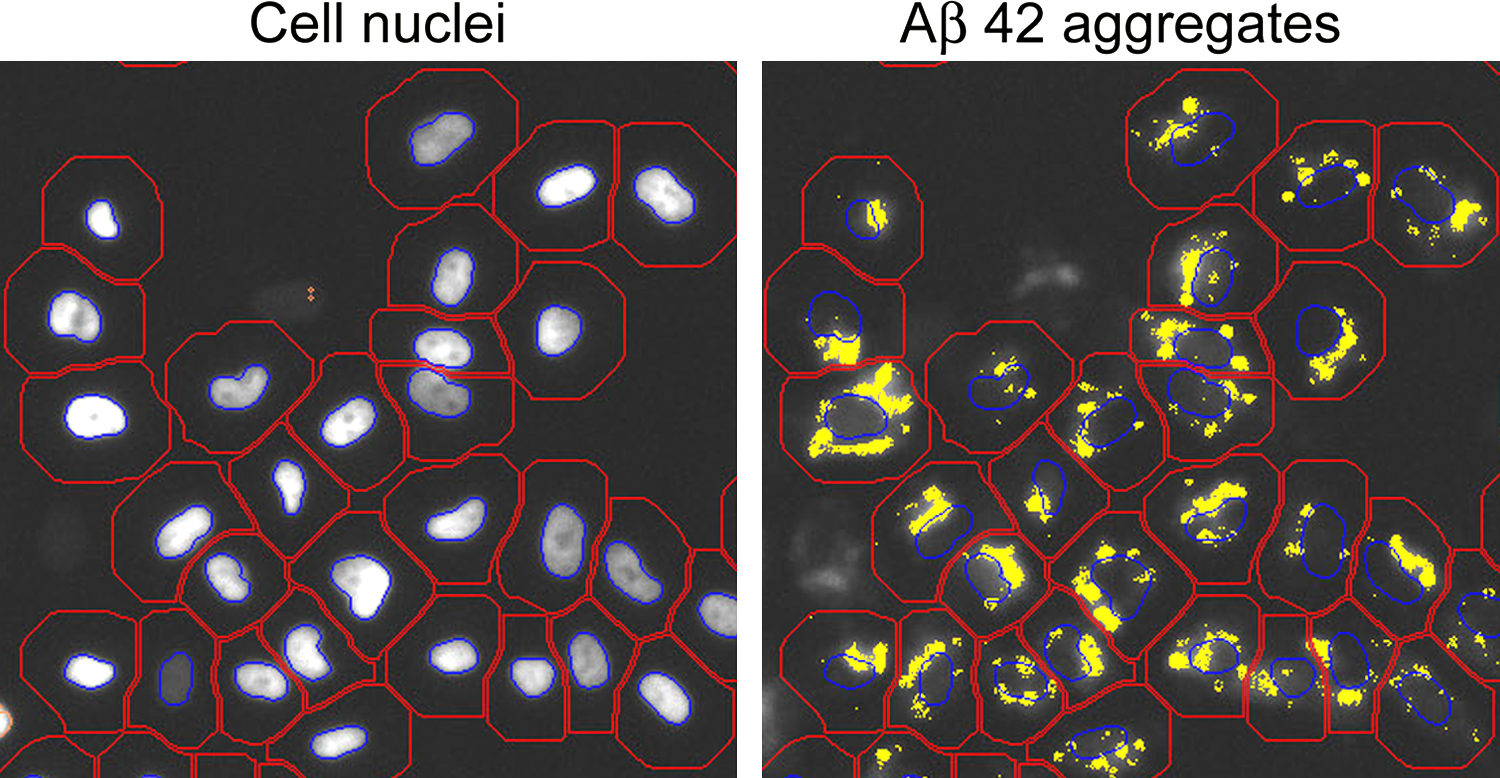
Fluorescence microscopy images of SH-EP cells with Aβ42 aggregates. Left: nuclear staining, right: Aβ42-Alexa 633 signal. Cell bodies (red contour lines), nuclei (blue contour lines), and Aβ42 aggregates (yellow) were identified by the image analysis software.
In order to test the effect of the laser light on ATP levels, SH-EP cells were seeded in 24-well plates and divided into two groups, controls and irradiated, conforming to the aforementioned parameters. We also incubated rat pheochromocytoma (PC12) cells with aggregated Aβ42 5 for 24 h, and subjected them to the same laser and EGCG treatment regime as described above. After irradiation, cells were incubated for further 48 h and cellular ATP content was quantified. Cellular ATP content was determined after lysing the cells (Tris-HCl 200 mM pH 7.7, NaCl 2 M, EDTA 20 mM, Triton X-100 0.2%) for 1 h at 4°C. Supernatants were cleared by centrifugation (14 krpm, 15 min, 4°C). Cellular ATP content was quantified using a luciferin/luciferase luminescence kit (A 22066, Invitrogen) and compared to standard ATP dilution series.
Results
A representative fluorescence microscopy image of Aβ42 aggregates in SH-EP cells is shown in Fig. 1. Figure 2 shows the results of the analysis for the Aβ42 aggregate amounts in groups 1 to 6. EGCG treatment reduced the intracellular Aβ42 aggregate amounts in SH-EP cells by up to 50% in a concentration-dependent manner. A similar reduction had been observed earlier in Chinese hamster ovary cells that overexpressed Aβ (7PA2 cells). 6 Coincidentally, laser irradiation significantly reduced intracellular Aβ42 aggregate amounts, when compared to nonirradiated cells (Fig. 2). We started with the working hypothesis that laser irradiation would increase the uptake of EGCG and thereby further reduce intracellular Aβ aggregates. Surprisingly, the reduction was observed even in laser-irradiated cells that were not treated with EGCG. We therefore conclude that Aβ aggregate reduction by laser irradiation does not require prior treatment by EGCG. Rather, both processes appear to affect the removal of Aβ aggregates complementarily.
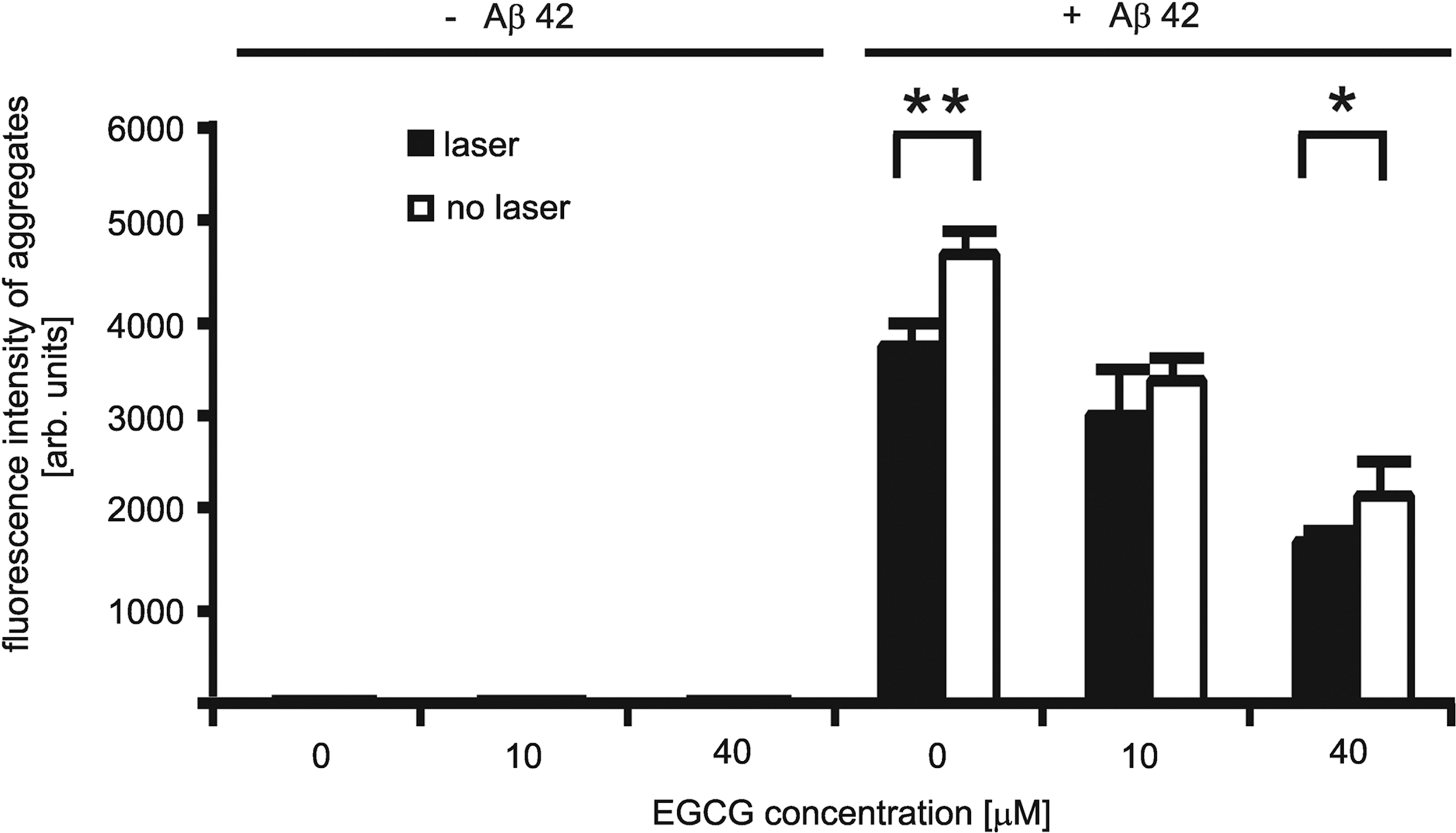
Quantification of the Aβ42 aggregate fluorescence signals. Fluorescence intensity is proportional to the Aβ42 aggregate amounts. Fluorescence is restricted to the wells containing SH-EP cells laden with Aβ42. Cells incubated with Aβ42 (+Aβ42) showed fluorescence signals. Cells incubated without Aβ42 (-Aβ42) showed no fluorescence. Laser irradiation significantly reduced intracellular aggregate load. Analysis of four independent wells per sample with ∼4500 cells per well: **p<0.002, *p<0.05.
Figure 3 illustrates the effect of the laser light on the proliferation of the SH-EP cells. We quantified the proliferative effect in the presence and absence of Aβ42 aggregates by counting cell numbers 24 h after irradiation. Similar to what has been observed in other model systems, 12 irradiation at 670 nm significantly increased cell proliferation, in our case by ∼20%. Concomitant treatment with EGCG did not affect cell proliferation in cells that did not contain Aβ42 aggregates. Remarkably, the proliferative effect was most pronounced in Aβ42-free cells. Interestingly, the proliferative effect of the light depended on the specific load of the cells. Relative to groups 1 to 3 (starting from the left in Fig. 3) the proliferation in group 4 (cells laden with Aβ42) was lower. And in groups 5 and 6 (cells double-burdened with EGCG and Aβ42) a proliferative effect was totally absent. This finding might have clinical relevance (c.f. Discussion section).
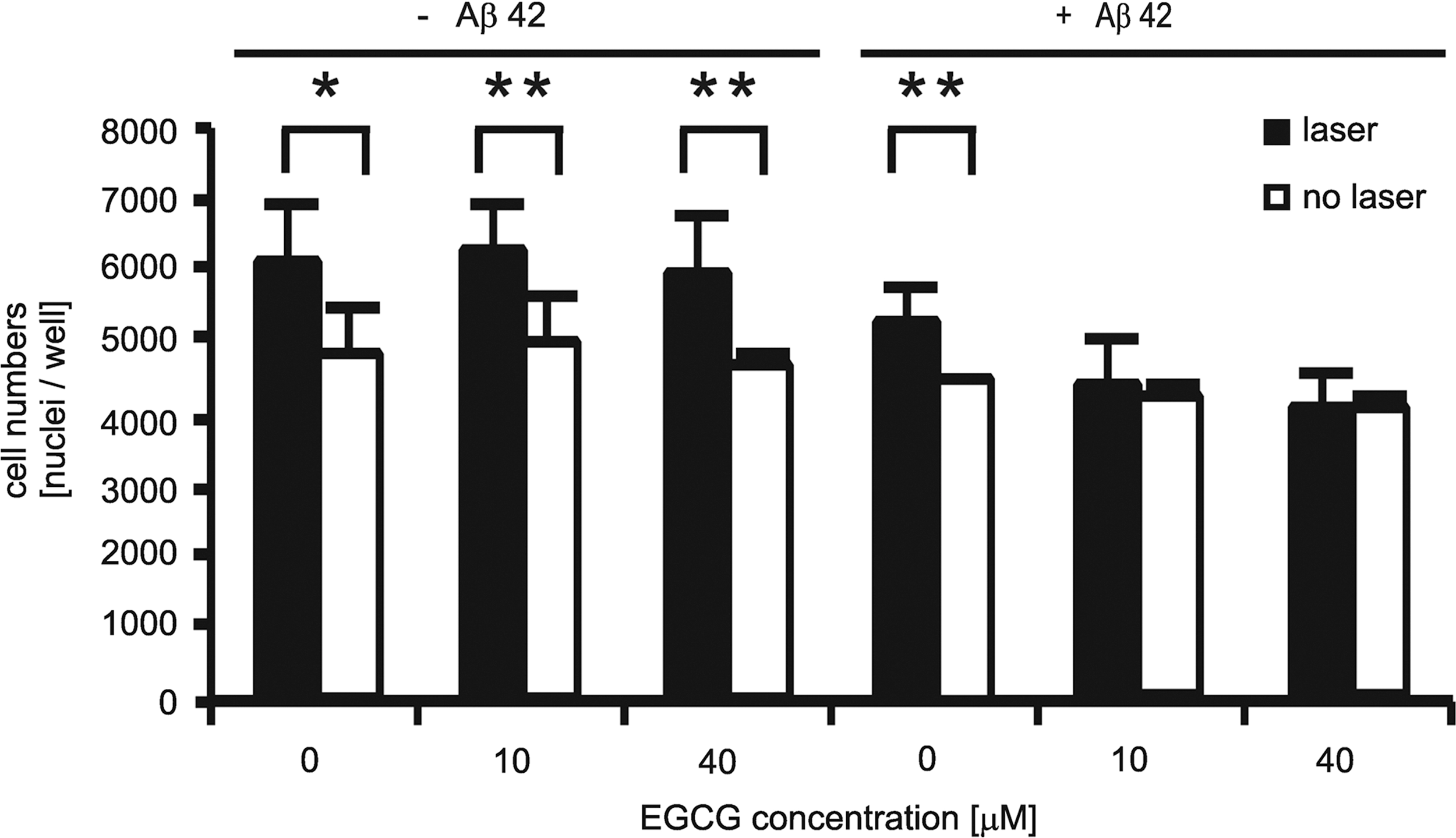
670-nm laser irradiation accelerates cell proliferation. Proliferation is reduced in Aβ42− and EGCG-treated SH-EP cells. Analysis of four independent wells: **p<0.02, *p<0.05.
We therefore conclude that clearance of intracellular Aβ aggregates is performed at the expense of cell proliferation—either the Aβ aggregates inhibit cellular metabolism or clearance of the aggregates redirects metabolic resources away from proliferation. To explore whether the process of Aβ clearance inhibited cellular metabolism, we quantified the ATP content of the cells under the different treatment conditions. For this, we irradiated two neuronal model cell lines, SH-EP and PC12 in the absence of Aβ42, using the previously established protocol. 11 In SH-EP cells we found a 20% increase in ATP levels (Fig. 4a), corresponding to the increase in cell proliferation (Fig. 3). PC12 cells presented a small increase in ATP levels, in the absence of further treatment (Fig. 4a). However, PC12 cells that were challenged with Aβ42 aggregates and then treated with a combination of laser irradiation and EGCG showed a significant reduction in ATP levels when compared to Aβ42-challenged but untreated cells (Fig. 4b).
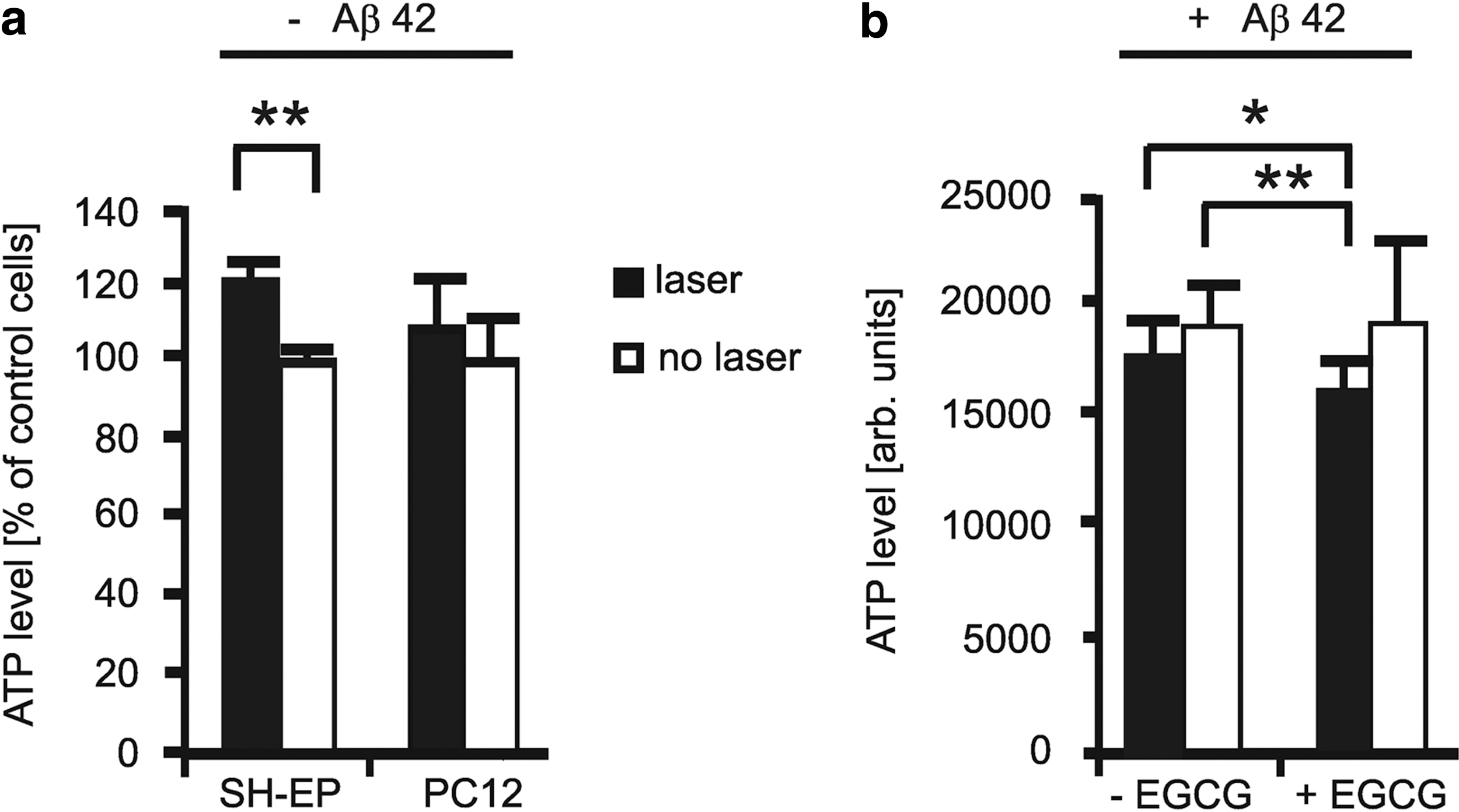
ATP quantification in irradiated and nonirradiated SH-EP and PC12 cells:
Discussion
The principal result of the present study is that 670-nm laser light and EGCG complementarily reduce intracellular Aβ42 aggregate amounts (Fig. 2). In interpreting this result, it is instructive to establish a connection with the result of Fig. 3 (increase in cell proliferation). It is well documented that exposure to 670-nm light at irradiance and fluence ranges described by the biostimulatory windows 12 stimulates cell proliferation and mitochondrial metabolism, which was also observed in our model system (Fig. 4a). According to current photobiology models, cell proliferation is primarily caused by a boost in ATP levels in the mitochondria, with cytochrome c oxidase as a receptor for 670-nm light, as suggested by several reports. 20 –25 Moreover, intermittent laser treatment with irradiation parameters adjusted to the biostimulatory windows 12 was found to facilitate the uptake of drugs such as EGCG by HeLa cells. 11
A dominance of the latter effect in our model system would manifest itself in potentiating the EGCG effect. Instead, we observed that both treatments affect Aβ load complementarily. Consequently, it is tempting to assume that EGCG transport into SH-EP cells by intermittent laser irradiation is less pronounced than for HeLa cells. It is now easy to see the reason for the apparent inefficiency of the intermittent irradiation mode. Inspection of Fig. 3 (groups 1 to 3) reveals that EGCG had only a minimal effect on the proliferation of nonirradiated SH-EP cells, as opposed to a massive inhibition on that of nonirradiated HeLa cells. 11 From Fig. 2, it is clear that EGCG has crossed the plasma membrane (otherwise it could not reduce intracellular Aβ42 levels). Comparison with the impact of EGCG on HeLa cells 11 displays differences in the action spectrum of the compound in general, and in its cytostatic potency in particular. In general, the efficacy of transmembrane convection depends upon both the cell's permeability to water and the fraction of intracellular water prevalent as nanoscopic interfacial water layers on hydrophilic surfaces. Notably, mitochondria contain considerable amounts of nanoscopic interfacial water layers. 17 Inspection of Fig. 1 reveals that internalized Aβ42 tends to condensate into a shell around the nucleus. A precondition for the establishment of the shell is passage across the plasma membrane and radial diffusion toward the nucleus. En route to the nucleus, the hydrophobic Aβ42, known to accumulate in intracellular vesicular structures, 2,3 might compete for proximal surfaces and contribute to a depletion of hydrophilic intracellular surfaces—the reservoirs of nanoscopic interfacial water layers.
From considering the stimulation of mitochondrial metabolism by the laser irradiation (Fig. 4) it is clear that the laser light plays a strong role in reducing Aβ aggregate load. We observed that cellular ATP levels were increased by laser irradiation, but that this increase was diminished in the presence of Aβ aggregates. Thus, light-induced removal of Aβ aggregates seems to lower ATP levels (Fig. 4). Of particular interest in this context is the absence of a proliferative effect of the laser light in double-burdened cells (Fig. 3, groups 5 and 6). Therefore, in the course of therapy, EGCG treatment prior to light treatment could be counterproductive. EGCG treatment of cells in the absence of Aβ had no significant impact on either cell proliferation or ATP levels (Figs. 3 and 4b). In contrast, ATP and cell proliferation were lowered when Aβ aggregates were removed by EGCG/irradiation treatment. Consequently, we feel justified in assuming that the anti-Aβ effect of the laser irradiation was performed at the expense of the cell's own currency: ATP.
However, there are uncertainties in our apparently straightforward experiments, which should be clearly mentioned. In other models, proliferative effects are related to a moderate increase in ROS in the mitochondria. 26,27 However, an increase in ROS levels provoked by the applied laser light seems implausible. EGCG is a potent ROS scavenger, 28 so an increase in ROS levels should oxidize EGCG and result in a decrease in EGCG activity. The ROS scavenging capacity of EGCG was recently exploited in preventing Aβ-induced ROS accumulation in primary cortical neurons. 29 Cell proliferation might even be controlled by an externally imposed metabolism (nutrient uptake) via intermittent irradiation, 11 possibly also observed in HEp-2 cells. 30 Of fundamental interest is the nature of the mechanism by which Aβ is cleared in response to 670-nm light irradiation—presumably, a cellular program fueled by the light. Our experiments cannot determine whether the light stimulates known pathways of protein homeostasis such as chaperone activity, ubiquitin-proteasome system response or autophagocytosis. 31 All of these pathways could account for ATP depletion but we cannot fully exclude an unknown photochemical effect involving mitochondrial ROS activated by 670-nm light.
Figure 5 illustrates the synergistic interplay between the chemical and physical mechanism of photobiomodulation. In both the irradiation parameters coincide with those found by the Mester group to accelerate wound healing. 32 In principle, it should be possible to combine ATP and/or ROS-related metabolic stimulation with the external modulation of nanoscopic interfacial water layers by optimizing irradiation parameters, an attractive challenge in photomedicine, which reflects the intense discussion on the subject. 33
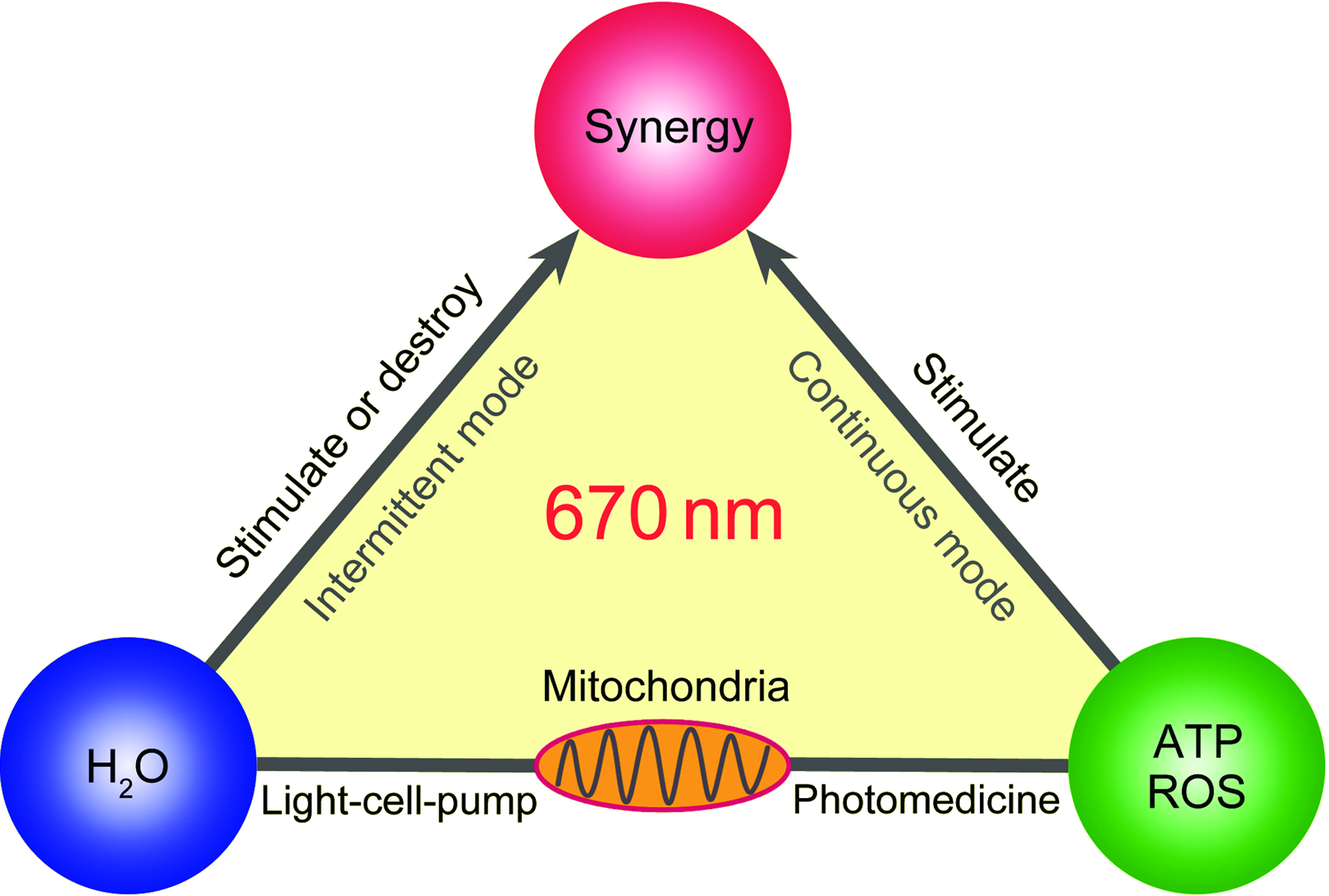
Visual synopsis of basic principles of photobiomodulation. Reproducibility in photobiomodulation started with the use of lasers operating in the 1–4·104 Jm−2 window. 32 The fluence window was found to be generally valid for red to NIR light both in vitro and in vivo. Central targets for the light activation are mitochondria; they respond to the light stimulation by producing ATP and/or ROS. At irradiances on the order of 1·103 Wm−2 most cells respond to stimulation with 1–4·104 Jm−2 by proliferating. Complementary metabolic processes are induced in cells with a reasonable water permeability by intermittent irradiation (same irradiation parameters). In the design of advanced irradiation protocols the image may serve as a diagnostic tool for optimization of cellular metabolic processes.
Our laboratory experiment receives possible therapeutic relevance from two sides: Animal experiments indicate the penetration of EGCG across the BBB. 9,10 Coincidentally, moderate irradiances (∼kWm2) of red to NIR light, including 670 nm, penetrate several centimeters of tissue, including the cranial bone, delivering therapeutically relevant irradiances. For example, in the case of extreme cardiac dysrhythmia with characteristic symptoms of angina pectoris, 30 min transcostal cardiac stimulation with a battery of seven 670-nm lasers, each 50 mW, pulse frequency 1 Hz, resulted in an instant normalization of the heart rhythm and total disappearance of the angina symptoms (A.P. Sommer, personal communication). Three minutes' irradiation of the heart of rats and dogs with NIR laser light (local irradiance and fluence at the myocardial level 60 Wm2, 1.08·104 Jm2, respectively) was already reported to reduce scar tissue formation after myocardial infarction. 34 The transcranial transmission of red to NIR light delivered by an array of light-emitting diodes has been recently illustrated in the treatment of chronic, traumatic brain injury. 35 A representative review of the literature on the use of moderate irradiances of red to NIR light in the treatment of acute stroke both in humans and animal models can be found in the literature. 36 Therefore, amyloid deposits in internal organs, including brain may be accessible to the combination of irradiation and drug treatment that reduced Aβ aggregate load in our cellular model. However, in vivo transfer of the irradiance level found to be effective in vitro at 670 nm needs careful adjustment. In view of the documented clinical evidence for the deep tissue penetration of 670-nm light, 37 and the reported clinical success in transcranial application of combinations of red and NIR light, it appears reasonable to extend the current tests and investigate the clearance of Aβ by combinations of wavelengths, in particular 670 nm and NIR. Expectedly, the functional interplay and complementary effects of laser irradiation and drug treatment are not restricted to the drug EGCG, but might also be an interesting combination for other therapeutic compounds. In view of the encouraging results, it is clear that further study is now necessary.
Conclusions
Irradiation with moderate levels of 670-nm light and EGCG supplementation complementarily reduces Aβ aggregate amounts in human neuroblastoma cells. Combination therapy is most effective when components are combined that have complementary mechanisms of action. It can be expected that EGCG is not the only compound which, in combination with 670-nm laser light, reduces Aβ. Generalization of the complementarity principle between light and drug may open totally new perspectives for the treatment of AD.
Footnotes
Acknowledgments
We gratefully acknowledge technical assistance by G. Grelle and S. Kostka. The SH-EP cell line was a kind gift from R. König, F Westermann and M. Schwab, German Cancer Research Center (DKFZ). This research was partly financially supported by the Helmholtz Alliance for Mental Health in an Ageing Society (HelMA), the German Research Foundation (DFG, BI 1409/2), and the German Federal Ministry of Education (BMBF, NGFN-Plus 01GS08132).
Author Disclosure Statement
No competing financial interests exist.