
Abstract
Introduction
The cellular mechanisms underlying the visible-light–tissue interaction have been extensively studied. 8,19,20 It is commonly believed that following illumination, reactive oxygen species (ROS) are formed as a consequence of light absorption by endogenous cellular photosensitizers. 19,21,22 Low fluxes of ROS serve as cellular signaling messengers that induce processes such as transcription factor activation, gene expression, muscle contraction, and cell growth. 23 –26
In order to be influenced by light, the cells must possess suitable photosensitizers. Potential endogenous photosensitizers that absorb light in the visible range include porphyrins,
27
–29
cytochromes
30
(especially the cytochrome c oxidase, as suggested by Karu,
19
) pyridine cofactors, nicotinamide adenine dinucleotide (NADH) and nicotinamide adenine dinucleotide phosphate (NADPH),
31
Fe-S clusters,
32
and flavins.
33
–36
The plasma membrane contains the NADPH oxidase system (a transmembrane complex containing both heme molecules and flavins), which is also a potential photosensitizer. This system produces superoxide radicals (
In order to investigate the involvement of the plasma membrane in the interaction of LEVL with the cell, we used the electron spin resonance (ESR) technique to measure ROS formation in irradiated isolated membranes. Sperm cells were chosen as the source of plasma membranes because of our previous finding that the capacitation process, which is a necessary stage in fertilization, is an ROS-dependent process occurring in the sperm plasma membrane. 12 To determine whether the light-induced ROS production is a result of a photo-accelerated electron transfer in the enzyme-catalyzed reaction with oxygen in the plasma membrane, or resulted from a photochemical reaction of the chromophores alone with oxygen, denatured membranes were irradiated as well.
Materials and Methods
Sperm-plasma membrane
Ram sperm-plasma membranes were prepared as previously described. 38 The membrane fraction was deposited on top of a 1.5 M sucrose layer and was removed and diluted using phosphate-buffered saline (PBS). The protein concentration in the membrane preparation was determined using the Lowry method. 39 Concentrations of 4 mg/mL and 18 μg/mL were obtained from fresh and frozen sperms, respectively. To denature membrane proteins, a suspension of 1.8 μg/mL membranes was heated in a 100°C water bath for 5 min.
Electron paramagnetic resonance (EPR) measurements using spin trapping
The EPR spin-trapping technique was used for the detection of oxyradicals. The spin trap 5,5-dimethyl-1-pyrroline-N-oxide (DMPO), purchased from Sigma, was used for ROS detection. DMPO is a common spin probe that reacts with •OH to produce a stable radical spin adduct, DMPO-OH (Eq. 1). When DMPO-OH is present in the solution, a quartet signal is observed in the EPR spectrum. DMPO also reacts with
The DMPO solution was purified in the dark using activated charcoal in PBS, pH 7.4. After ∼30 min, the solution was filtered, and its concentration was determined spectrophotometrically, ɛ227nm=8.0 mM−1×cm−1. 41 The solution was stored at −20°C for no longer than 2 weeks.
Samples containing 0.4 mg or 1.8 μg membrane proteins/mL (from fresh and frozen sperms, respectively) and 0.04 M DMPO were drawn by a syringe into a gas-permeable Teflon capillary (Zeus Industries, Raritan, NJ) and inserted into a narrow quartz tube open at both ends, as described previously. 42 The tube was placed within the EPR cavity and the spectra were recorded on a Bruker EPR 100d X-band spectrometer during or after illumination of samples. The EPR measurement parameters were as follows: frequency: 9.75 GHz; microwave power, 20 mW; scan width, 85 Gauss; resolution, 1024; receiver gain, 2×105; conversion time, 82 ms; time constant, 655 ms; sweep time, 168 sec; number of scans, 1 (unless stated otherwise). For the quantification of spin concentration, double integration of the signal was performed using Bruker software WIN-EPR, version 2.11 software, and calibrated with the stable radical, 2,2,6,6-tetramethyl piperidine-N-oxyl (TEMPO) (purchased from Sigma): a calibration curve displays different concentrations of TEMPO versus the double integration of the EPR triplet spectra of TEMPO. 43 The concentration of the DMPO-OH adduct was calculated by comparing the average double-integration area of the two DMPO-OH central peaks to the TEMPO calibration.
Illumination
A conventional source of visible light (400–800 nm) was used to illuminate the samples in the EPR cavity at a power density of 40 or 130 mW/cm2. The incident light produced no detectable heating. For measuring ROS formation as a function of blue and red light, dichroic filters were used. For red light, a filter transferring 600–800 nm was used, whereas a 400–505 nm filter was used for blue light. The filters were attached to the EPR cavity during illumination, resulting in power densities of 30 and 10 mW/cm2 for red and blue light, respectively.
Results
Production of oxyradicals in illuminated membranes
ROS generation in visible light–illuminated isolated sperm membranes was studied. To this end, we measured the DMPO-OH signal in illuminated sperm membranes. A 5-min illumination resulted in an increase of the DMPO-OH adduct signal showing its characteristic quartet signal with an intensity profile of 1:2:2:1 (see Fig. 1b). This signal monitors •OH and/or
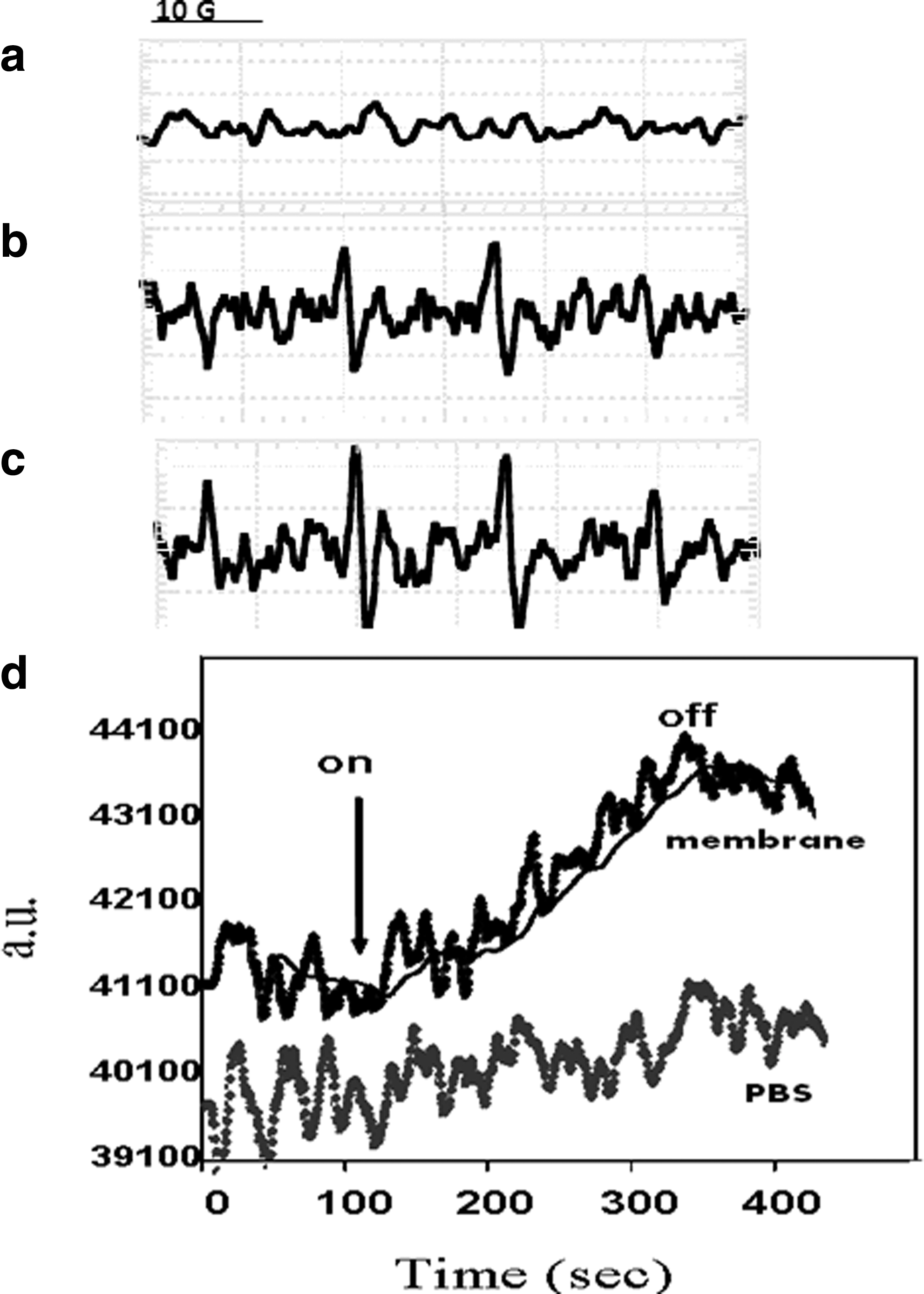
Spin adduct signal of 0.4 mg/mL sperm membrane with 0.033 M DMPO upon illumination :
The kinetics of DMPO-OH formation was further investigated using static field measurements. The intensity of the low-field DMPO-OH peak was measured as a function of time, as presented in Figure 1d. As DMPO-OH has a limited lifetime, each point on the graph reflects both the rates of adduct formation and its decomposition. It can be seen that DMPO-OH production began immediately upon the onset of illumination and increased during the ∼200 sec of illumination. The illumination was then turned off. When the same illumination protocol was performed with PBS containing DMPO but no membranes, the signal remained at the baseline (Fig. 1d, bottom line). These results indicate that isolated sperm membranes produce oxygen radicals simultaneously to light activating.
Confirmation of •OH generation
The formation of the DMPO-OH quartet, as presented in Fig. 1, can be attributed to the formation of both superoxide and •OH. In order to determine whether the signal detected is the result of the production of superoxide and/or •OH, we used DMSO, which is known to scavenge •OH. In Fig. 2, the intensity of the first low field peak of DMPO-OH as a function of illumination time with and without DMSO, is presented. As can be seen in Fig. 2, illumination of the membranes in the presence of DMPO and DMSO almost abolished the DMPO-OH low field-peak signal. It can therefore be concluded that the DMPO-OH signal results mainly from the direct trapping of •OH.
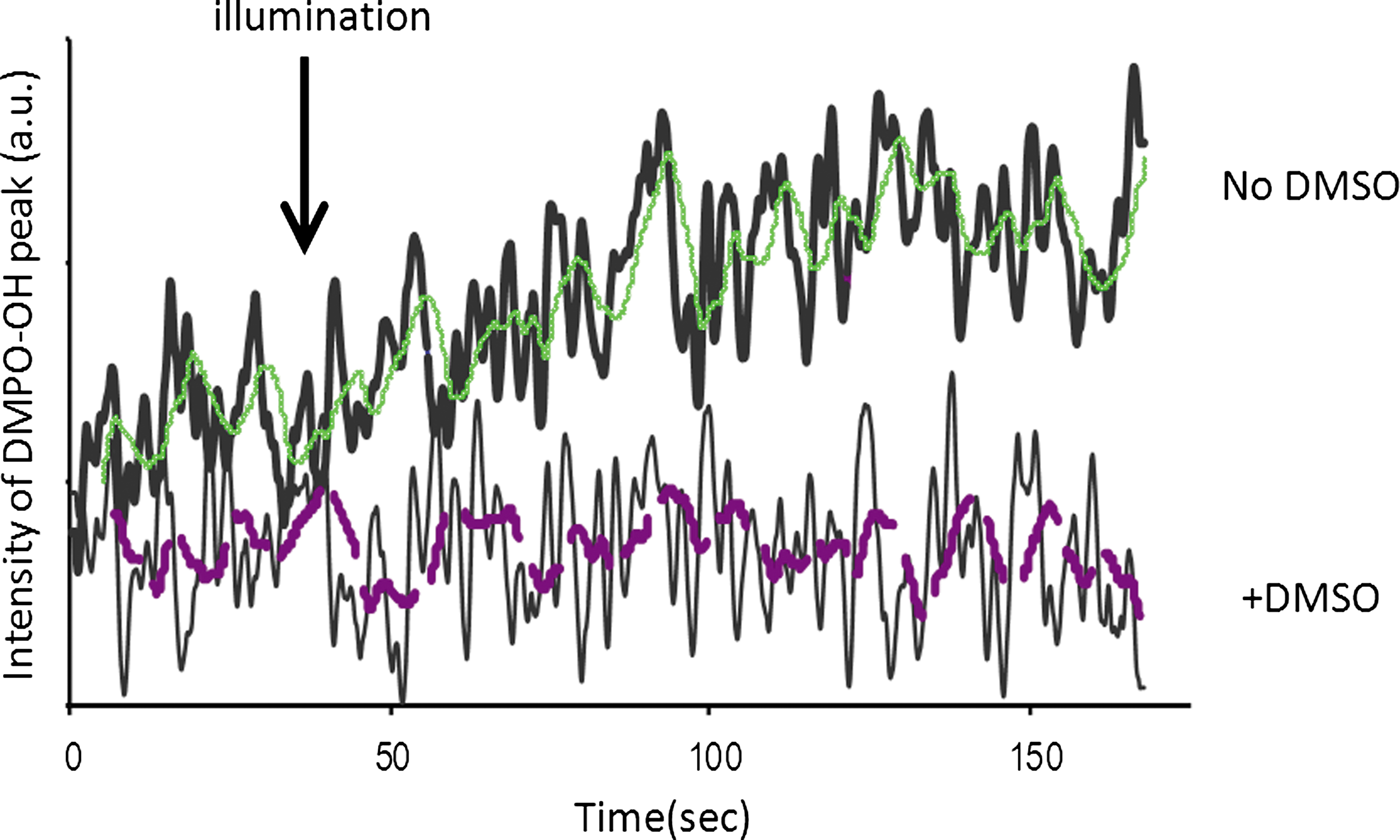
The effect of DMSO on the DMPO-OH signal. The graph presents the intensity of the first low field peak of DMPO-OH plus membranes (0.4 mg/mL) upon illumination in the presence or absence of DMSO. Experimental parameters: scan width, 100 G; sweep time, 84 sec; # of scans, 2. The fine line is the moving average curve of the original (bold) signal.
Membranal oxyradicals as a function of the illuminated wavelength
To study wavelength dependence of ROS generation by plasma membranes, we compared the ability of red (600–800 nm) and blue (400–505 nm) light to induce oxyradical formation in illuminated membranes. The amount of ROS generated by red and blue light was normalized according to the intensity of the light: 30 and 10 mW/cm2, respectively. Figure 3 presents a histogram of the normalized double-integrated DMPO-OH spin adduct signal, which monitors ROS levels following 170 sec illumination of sperm membrane with red or blue light. The result strongly suggests that the blue light is much more effective than the red light.
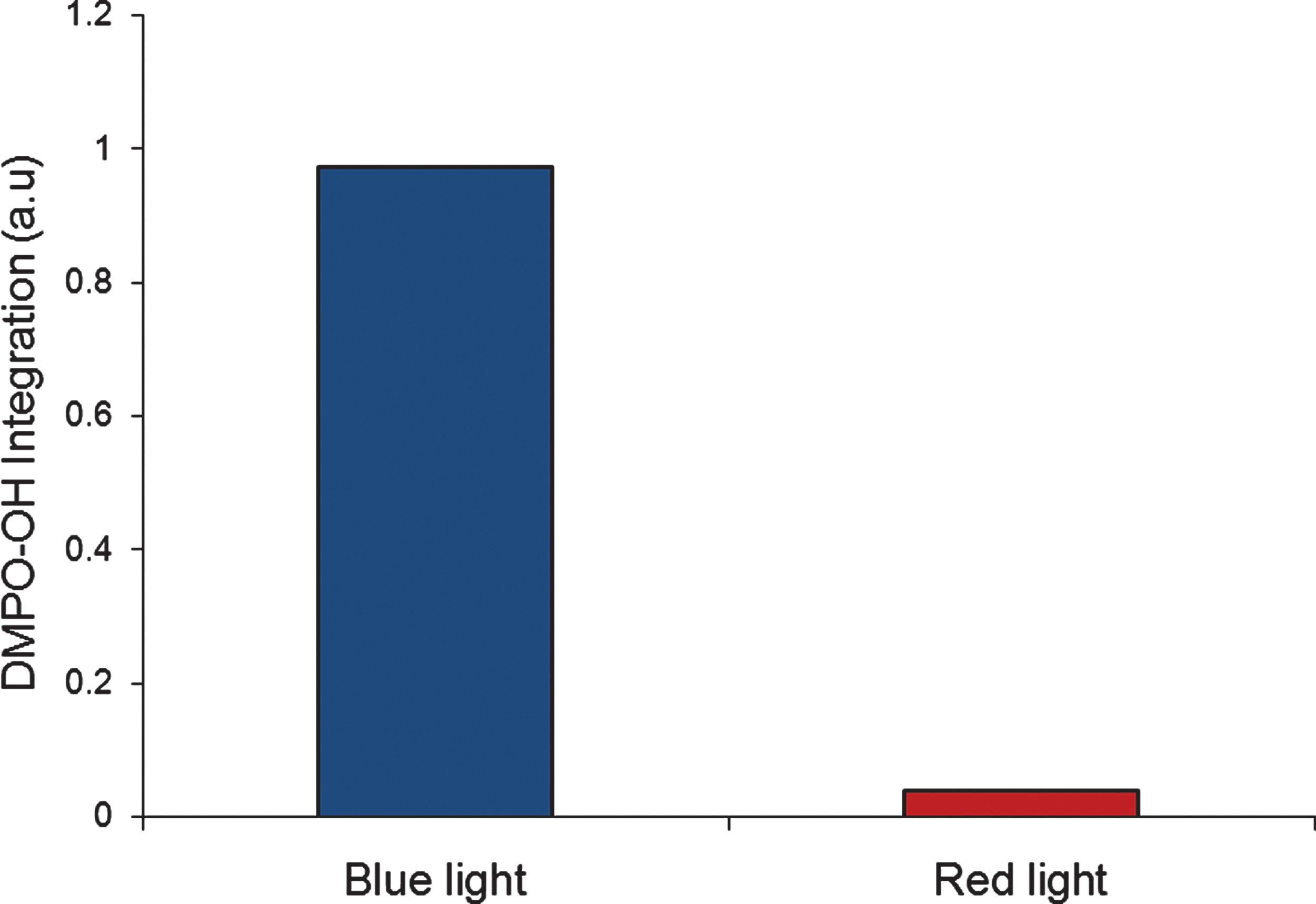
ROS generation in sperm membrane following red or blue light illumination. The graph shows the normalized (to light intensity) double integrated DMPO-OH spin adduct signal in sperm membranes (1.8 μg/mL) illuminated for 170 sec, with the appropriate dichroic filters for red or blue light illumination.
ROS production in illuminated denatured sperm membranes
In order to determine whether ROS generation depends upon the membrane proteins activity or whether light interaction with the membrane is a chemical process only, the EPR spectra of denatured plasma membranes before and after light irradiation were measured. As can be seen in Fig. 4, there is an increase in the DMPO-OH signal despite the denaturation (Fig. 4b), proving the presence of oxyradicals in illuminated denatured membranes.

The effect of illumination on ROS production by denatured membranes:
Discussion
In this study, the interaction of visible light with the sperm plasma membrane was analyzed to determine the ability of light to elicit oxygen radicals in the membrane preparations. Sperm membranes were chosen since, in our previous work, 12 we found enhancement of capacitation and acrosome reactions, which are ROS-dependent processes, occurring in the sperm plasma membrane, 44 following visible-light illumination.
Production of ROS in visible-light–illuminated plasma membranes has been demonstrated in the past by adding various photosensitizers.
45,46
As can be seen from Fig. 1, sperm membranes, without any additional exogenous photosensitizers, can produce oxyradicals (•OH and/or
However, in order to address whether light interaction with the NOX complex depends on the functional state of these proteins, we denatured plasma membranes and studied ROS formation in illuminated denatured membranes. As can be seen in Fig. 4b, illumination of denatured membranes resulted in a clear DMPO-OH signal that confirmed the formation of oxyradicals in these membranes. Therefore, it emerges that light generates ROS mostly through a photochemical process, involving only chromophores and oxygen without the involvement of cellular proteins.
Following light illumination and ROS generation by membranal chromophores, the redox state of the membrane might change, thereby activating cellular activities. It has been suggested that the redox state of the membrane is important for regulation of cell function and growth. 49 For example, in endothelial cells, a change in the cell surface thiol/disulfide redox state causes oxidation of several surface proteins, which in turn affects cell adhesion and aggregation. 50 Oxidation and reduction of specific cysteine groups on membranal protein tyrosine kinases, which function as a molecular switch, can trigger various signaling pathways. 51,52 Changes in the membrane redox state can also cause proton release and changes in intracellular calcium ([Ca2+]I), and alter the growth process. 49 In a previous study, we showed that visible light causes a transient increase of [Ca2+]i. 12,53 Based on these results, we suggest that light induces some intracellular changes in response to the change in redox state, thus leading to cell–membrane activation. In particular, in sperm membranes, this leads to enhanced capacitation and acrosome reaction. 54 –56
Conclusions
In this study, we show the generation of oxyradicals by isolated sperm plasma membranes following visible-light illumination. Blue light was found to be more effective than red light. ROS generation was found to be mainly a photochemical process, which is not dependent upon enzymes.
The ROS-induced change in the membrane redox state might lead to membrane channel opening, thereby stimulating biological processes. In sperm cells, generation of ROS in plasma membranes might play an important role in the capacitation process and acrosome reaction (necessary stages in fertilization).
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.