
Abstract
Introduction
The superoxide anion (•O2 -), hydrogen peroxide (H2O2) and the hydroxyl radical (•OH-) are major ROS in human tissues. The superoxide anion is generated within the cells by several enzyme systems including nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, nitric oxide synthase, and xanthine oxidase, as well as via the mitochondrial respiratory chain. The one-electron reduction of molecular oxygen produces a relatively stable intermediate, the superoxide anion, which serves as the precursor of most ROS. The mitochondrial electron transport chain contains several redox centers that can leak electrons to molecular oxygen, which serves as the primary source of superoxide production in most tissues.
Superoxide production through these biological systems is subject to intricate regulation by a variety of hormones, cytokines, and toxins. The level of ROS generated in cells is controlled by various intracellular enzymatic antioxidants, such as superoxide dismutase (SOD), glutathione peroxidase (GP), and catalase (CAT). 2,3 SOD catalytically dismutes the superoxide anion to H2O2, whereas CAT and GP render H2O2 harmless within the cells by converting it into water and oxygen. 4 GP utilizes glutathione (GSH) to destroy H2O2 and other lipid hydroperoxides. Therefore, these enzymes regulate the intracellular levels of ROS like H2O2 and superoxide anion. Animal cells produce several different superoxide scavenger enzymes located in different areas of the cell. Cu/ZnSOD accounts for 50–80% of the total SOD and is localized primarily in the cytosol and nucleus. MnSOD is expressed in smaller quantities in the cells, more abundantly in endothelial cells (ECs), and is localized to the mitochondria. Extracellular SOD is bound to the cell membrane through its heparin-binding domain and is located extracellularly. 5,6 Oxidative stress occurs when there is a disturbance in the oxidant–antioxidant balance in favor of the former. This imbalance can cause damage at the macromolecular level including DNA strand breakage, damage to membranes and membrane transport systems, oxidation of enzymes and other proteins, and lipid peroxidation. 7
Recently, many clinical studies have reported that low-level light treatment (LLLT) at a wavelength of ∼635 nm can serve as an anti-inflammatory tool for accelerating wound healing, 8,9 and for pain reduction. 10 –19 Many reports have suggested that scavenging of intracellular ROS by irradiation with visible light can be applied to improve a number of pathological states. 10,20,21 A specially manufactured visible light irradiation tool can be used in treatment of many degenerative diseases and to improve the electron transfer of cytochrome c oxidase in the mitochondria, as well as adenosine-5′-triphosphate (ATP) synthesis and cellular metabolism. 22,23 In a previous study, irradiation at 635 nm had an inhibitory effect on PGE2 synthesis similar to that observed after treatment with cyclooxygenas (COX) inhibitors. 10 However, unlike the mode of action of the usual nonsteroidal anti-inflammatory drugs (NSAIDs), indomethacin and ibuprofen, this inhibitory effect was attributed to a decrease in the ROS levels. In another study, irradiation with 635 nm suppressed ONOO− synthesis via the dissociation of ROS in SH-SY5Y neuroblastoma cells treated with SNP, an NO donor. 24 These phenomena were attributed to the relationship between 635 nm irradiation and scavenging of intracellular ROS.
However, the light-induced ROS scavenging mechanism is not yet completely understood. Especially, its relationship with intracellular enzymatic ROS scavengers is obscure. This study examined the type of ROS that can be scavenged by an 635 nm irradiation system in an in vitro model, and compared the result with that of an 470 nm irradiation system, which operated on a different photoacceptor. In addition, the independent or dependent action of intracellular enzymatic scavengers, such as SOD, GSH, and catalase (CAT) on ROS removal, was investigated with inhibitor studies in an immortalized oral keratinocyte cell line.
Materials and Methods
Cell culture
Immortalized human oral keratinocyte HaCaT cells were maintained in Dulbecco's modified Eagle's medium (DMEM, Gibco BRL, USA) supplemented with an antibiotic-antimycotic solution (Gibco BRL, USA) and 10% heat-inactivated fetal bovine serum (JR Scientific Inc., USA) in a humidified atmosphere of 5% CO2/95% air at 37°C. The cells were cultured for 24 h at a density of 1×105cells/mL in a 9 cm culture dish (Corning Inc., USA) or 96-well plates (Corning Inc., USA) according to the type of analysis to be performed.
Chemical treatments
All chemicals except for SOD (Calbiochem, USA) were purchased from Sigma (Sigma Aldrich, USA). 100 μM xanthine and a specified concentration of xanthine oxidase were added to the cell cultures for superoxide anion generation. A specified concentration of H2O2 was supplied exogenously directly to the cultured cells. 25 SOD or N-acetyl-L-cysteine (NAC) were added to the cultured cells simultaneously with xanthine/xanthine oxidase (XXO) or H2O2 in order to compare their effects with those of 635 nm irradiation. Specified concentrations of inhibitors of intracellular ROS scavengers (diethyldithiocarbamate [DDC], a SOD inhibitor, buthionine sulfoximine [BSO], a GSH inhibitor, and 3-amino-1,2,4-triazole [ATZ], a CAT inhibitor) were added. 26,27 At the conclusion of all chemical treatments, the cultured cells were washed three times with phosphate-buffered-saline.
Light source and irradiation
The cultured cell samples were irradiated for 1 h with a light dose of 5 mW/cm2 and chemically treated simultaneously. A manufactured irradiation tool kit (Biophoton Co, Korea) was built in a 5% CO2 humidified chamber held at 37°C, and irradiation was performed for the specified time. The light at a wavelength of 635 nm from a LED source (U-JIN LED, Korea) was used for irradiation, and the result was compared with that at a wavelength of 470 nm. The detailed information about light irradiation is represented in Table 1. 28
Cell viability
The cell viability was measured by the reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, Sigma, USA). The MTT assay is based on the ability of mitochondrial dehydrogenases in viable cells to reduce MTT to formazan through cell respiration. The medium was removed 24 h after the experimental treatments, and the cells were incubated in phosphate-buffered saline (PBS) containing 30 μL of MTT at 37°C for 3 h. The formazan product was solubilized by the addition of 50 μL of dimethyl sulfoxide (DMSO, Calbiochem, USA). The optical density was measured at 570 nm using an Enzyme-linked immunosorbent assay (ELISA) reader (ELx800uv, BIO Tek Instruments Inc., USA).
Electron spin resonance (ESR) spin trapping determination of superoxide anion (•O2 -)
The superoxide anion was detected by an ESR spectrometer using 5,5-dimethyl-1-pyrroline-N-oxide (DMPO, Dojindo, Japan) as the spin trap. 29 Briefly, 1 h after chemical treatment or immediately after irradiation for 1 h, 1.5 μL of DMSO, 10 μL of sample (1×105 cells in 0.1M phosphate buffer, pH 7.4) and 40 μL of 1M DMPO were placed in a test tube and mixed. The effects of the hydroxyl radical were eliminated by adding DMSO, which is used as a hydroxyl radical scavenger. The mixture was transferred to glass capillaries (50 μL, Paul Marienfeld GmbH & Co. KG, Germany) and the DMPO-O2 - spin adduct was quantified. The measurement conditions for ESR (JEOL, Japan) were as follows: field sweep, 317–337 mT; field modulation frequency, 100 kHz; field modulation width, 0.4 mT; amplitude, 300; sweep time, 30 sec; time constant, 0.01 sec; microwave frequency, 9.166 GHz; and microwave power, 10 mW. The signal intensities were evaluated from the peak area of the first signal of the DMPO- O2 - spin adducts.
Flow cytometric analysis for detection of H2O2 formation
H2O2 was assayed using 2′,7′-dichlorodihydrofluorescein diacetate (H2DCF-DA; Sigma, USA). DCF-DA enters the cells passively, where it is enzymatically deacetylated by esterases to become the nonfluorescent 2,7-dihydrodichlorofluorescein (DCF-H). H2O2 can oxidize DCF-H to a highly fluorescent DCF form. 30 For measuring the intracellular H2O2 level, the cells were detached 1 h after the chemical treatment or immediately after irradiation for 1 h using a trypsin-EDTA solution, and then incubated with 10 μM of DCF-DA for 20 min. After incubation, the H2O2 level was analyzed by flow cytometry (Beckman Coulter, USA) with excitation at 485 nm and emission at 530 nm.
Lipid peroxidation
At 24 h after treatment, the level of lipid peroxidation was measured using a malondialdehyde (MDA) colorimetric assay kit (Oxford Biomedical Research, USA). Oxygen radicals affect the double bonds of polyunsaturated fatty acids in the cell membrane resulting in the formation of highly reactive lipid hydroperoxides. MDA is one of the most studied lipid peroxidation products. 31 The concentration of intracellular MDA was measured as an indicator of oxidative stress. A colorimetric assay reagent was applied to the cells according to the manufacturer's instructions, and the absorbance was measured at 586 nm using a colorimetric microplate reader (Bio Tek, USA).
Statistical analysis
All experiments were carried out performed in triplicate. The data are expressed as mean±standard deviation. In order to analyze the differences, the results were subjected to an analysis of variance (ANOVA) using SPSS for Windows Version 12.0 (SPSS, USA). A p value <0.05 was considered significant.
Results
The effect of antioxidants, inhibitors of intracellular antioxidants, and ROS generation on cell viability
To determine the concentrations of SOD and NAC, the cell viability was first measured in the XXO-treated HaCaT cell line. The cells were then treated with different doses of SOD (0, 100, 200, 400, and 800 U/mL, Fig. 1A) and NAC (0, 0.01, 0.1, 1, and 10 mM, Fig. 1B). Cell viabilities were expressed as the percentage of cell viability of untreated controls. And 200 U SOD and 0.1 mM NAC were used for further study.
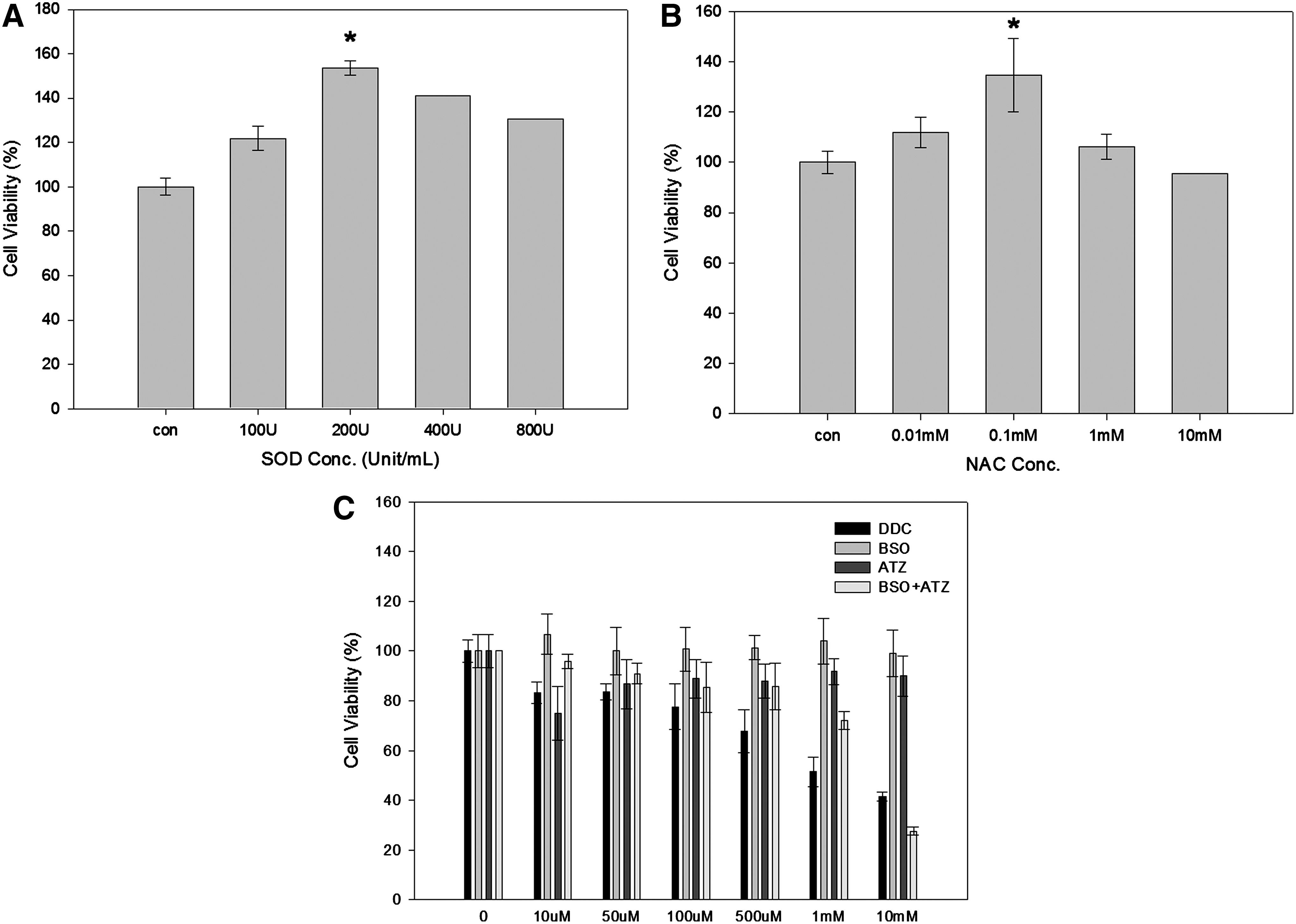
Cell viability after treatment at indicated concentration of superoxide dismutase (SOD) (0, 100, 200, 400, 800 U/mL
To determine the effect of the inhibitors of intracellular antioxidants on cell viability, different concentrations (0, 10, 50, 100, 500 μM, 1, 10 mM) of DDC, BSO, and ATZ were added to the cultured cells and incubated for 24 h. Incubation with DDC, a known inhibitor of Cu/ZnSODs, led to a dose-dependent decrease in cell viability (Fig. 1C). However, BSO or ATZ did not individually cause a decrease in cell viability, not even at a concentration of 10 mM. However, simultaneous treatment with BSO and ATZ decreased cell viability in a dose-dependent manner. Therefore, 1 mM DDC and BSO+ATZ were used in further studies.
The addition of increasing amounts of XXO or H2O2 to the cultured cells led to a dose-dependent decrease in cell viability to <40% of the control and eventually to cell death (Fig. 2A and B). SOD (200 U) treatment and 635 nm irradiation prevented cell death. Especially at a concentration of 40 mU/mL XXO, 635 nm irradiation contributed to the recovery of cell viability to 80% of the control. On the other hand, 635 nm irradiation was assumed to be an ineffective tool for preventing loss of cell viability in the cells exposed to exogenous H2O2.
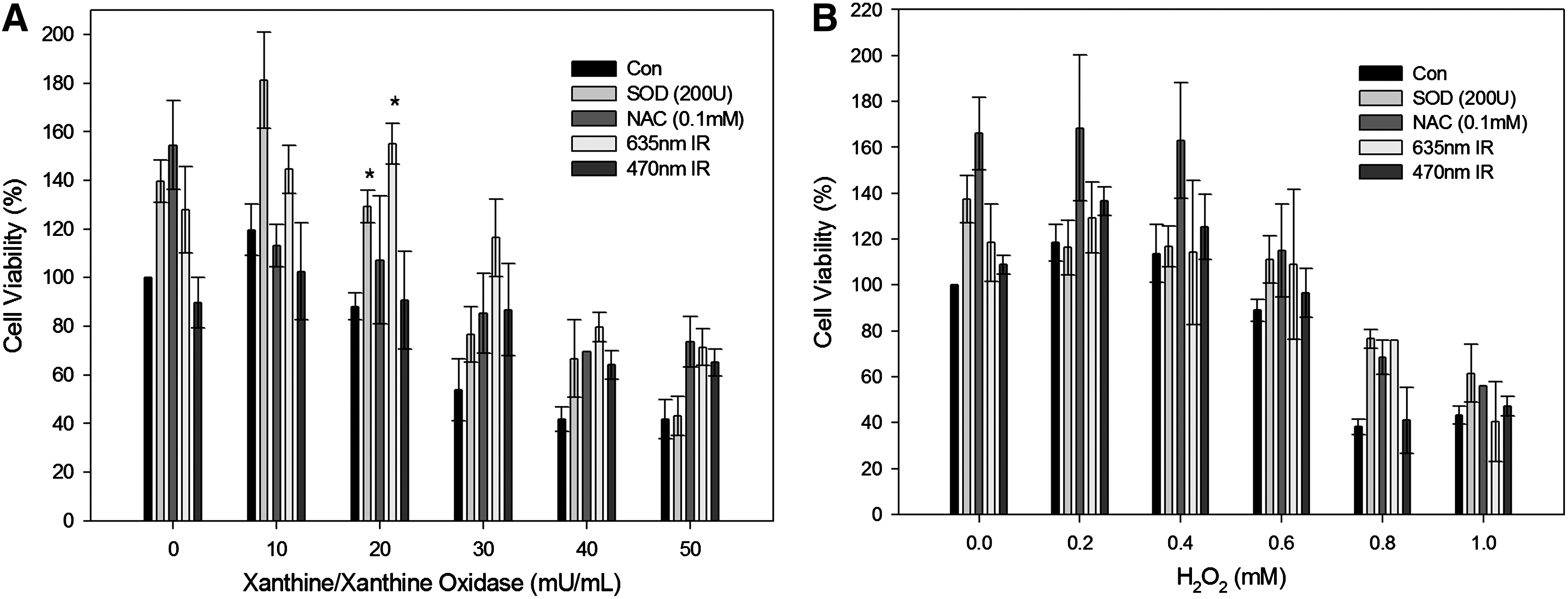
HaCaT cell viability after exposure to indicated concentration of xanthine/xanthine oxidase (XXO)
Level of superoxide anion/H2O2 and inhibitory effect of lipid peroxidation
The superoxide anions were measured by ESR analysis in the XXO (40 U)-treated cells (Fig. 3). The ESR signal intensity (peak area) increased to 4662 (arbitrary unit) by XXO treatment, and then decreased to 2210 by 635 nm irradiation. The level of superoxide anion production was decreased to 2689 by SOD treatment. However, there was no change in the level of superoxide anion production in the DDC (1 mM)-treated cells. Irradiation at 635 nm caused a decrease in the level of superoxide anion production in the presence or absence of DDC. The peak area at 470 nm irradiation was always higher than that observed at 635 nm irradiation.
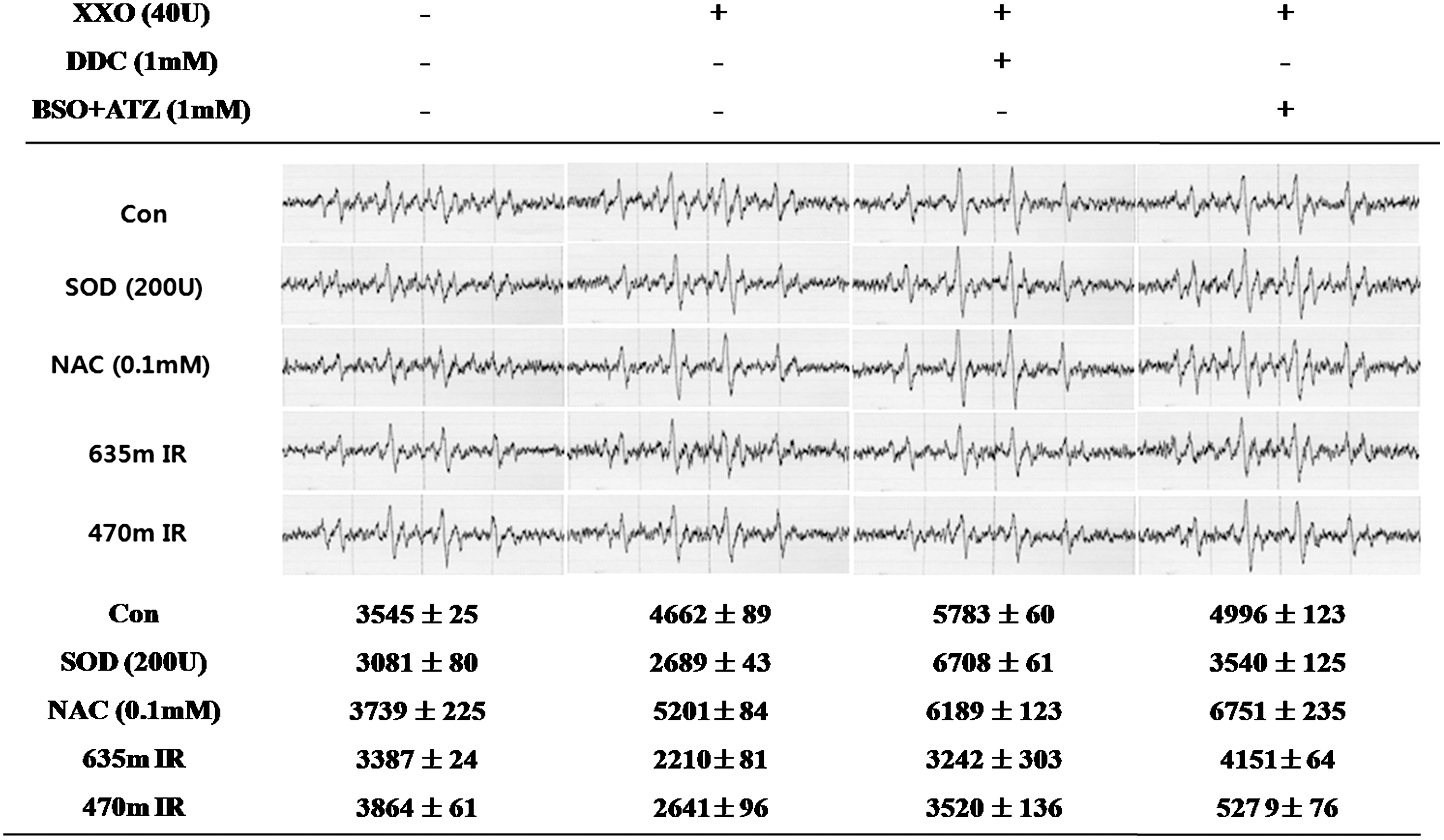
DMPO-O2 - peak and first peak area of DMPO-O2 - in a HaCaT cell line by electron spin resonance (ESR) spectroscopic measurement after a xanthine/xanthine oxidase (XXO) (40 mU/mL) treatment. Superoxide dismutase (SOD) (200 U), N-acetyl-L-cysteine (NAC) (0.1mM) treatment or 635 or 470 nm irradiation was performed for 1 h simultaneously with the XXO treatment. In addition, diethyldithiocarbamate (DDC) (1 mM) or buthionine sulfoximine and 3-amino-1,2,4-triazole (BSO+ATZ) (1 mM) were added together with XXO. The DMPO-O2 - peak was measured 1h after initiating the treatment. The values indicate the means±SD (n=3).
In the cells treated with H2O2 (0.8 mM), the level of superoxide anion production was decreased by 635 nm irradiation in all the groups (Fig. 4). The addition of NAC, a GSH precursor, led to a slight decrease in the level of superoxide anion production to 3726, but it could not reduce the level of superoxide anion production in the BSO+ATZ (1 mM)-treated cells. The peak area at 470 nm irradiation was slightly decreased to 2801 when the cells were treated with DDC in the presence of, H2O2, however it was always higher than that observed at 635 nm irradiation.
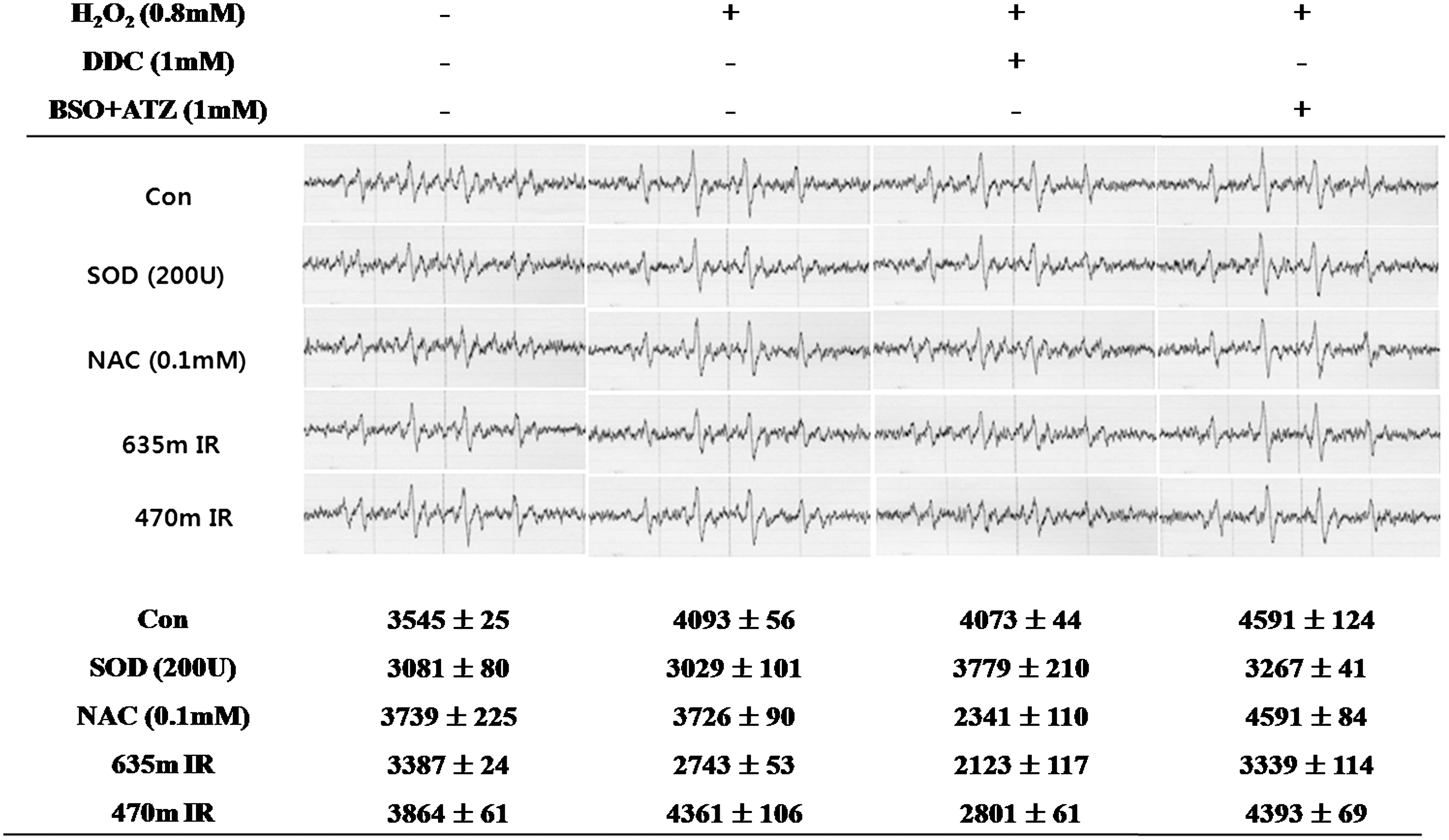
DMPO-O2 - peak and first peak area of DMPO-O2 - by electron spin resonance (ESR) spectroscopic measurement on H2O2 (0.8 mM) induced in HaCaT cell line. Superoxide dismutase (SOD) (200 U), N-acetyl-L-cysteine (NAC) (0.1 mM) treatment or 635nm, 470nm irradiation was performed for 1 h simultaneous with H2O2. Diethyldithiocarbamate (DDC) (1mM) or buthionine sulfoximine and 3-amino-1,2,4-triazole (BSO+ATZ) (1 mM) were also added with H2O2 at the same time in each group. At 1 h after the treatment, the DMPO-O2 - peak was measured. The values indicate the means±SD (n=3).
The concentration of intracellular H2O2 was decreased to 1.33 (arbitrary unit) by 635 nm irradiation or to 1.21 by 470 nm irradiation in the XXO-treated cells (Fig. 5). DDC treatment led to a slight decrease or increase in the intracellular H2O2 level. However, none of the treatments including SOD, NAC, or 635 or 470 nm irradiation decreased the H2O2 concentration in the BSO+ATZ (1 mM)-treated cells.
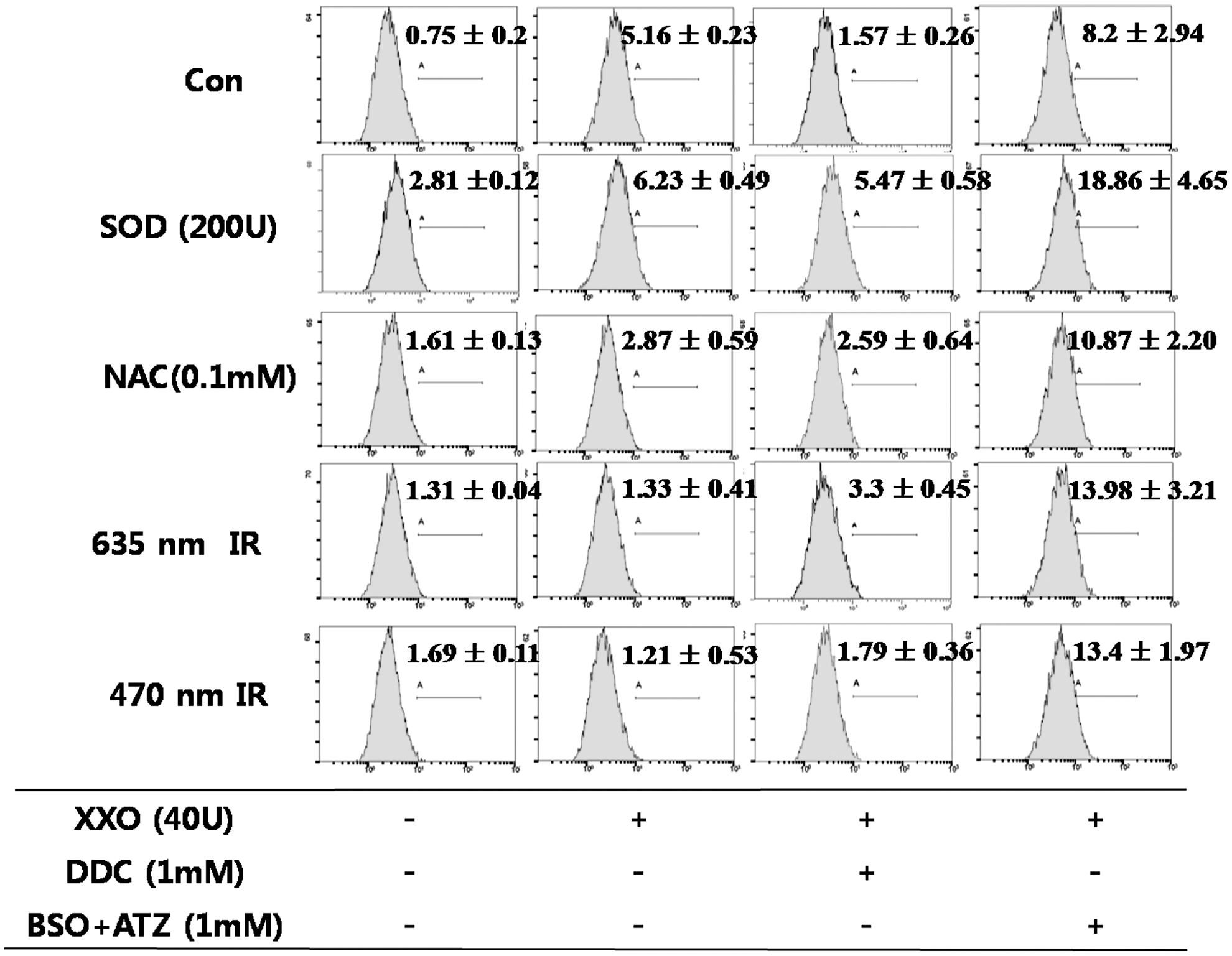
Flow cytometry analysis of 2′,7′-dichlorodihydrofluorescein (DCF) on xanthine/xanthine oxidase (XXO) (40 mU/mL) induced in the HaCaT cell line. Superoxide dismutase (SOD) (200 U), N-acetyl-L-cysteine (NAC) (0.1 mM) treatment or 635 or 470 nm irradiation was performed for 1 h simultaneous with XXO. Diethyldithiocarbamate (DDC) (1 mM) or buthionine sulfoximine and 3-amino-1,2,4-triazole (BSO+ATZ) (1 mM) were also added with XXO at the same time in each group. At 1 h after treatment, flow cytometry was performed. The value indicate the means±SD (n=3).
An exogenous treatment with H2O2 increased the intracellular H2O2 level to 11.14 (Fig. 6). NAC scavenged H2O2 to some extent. In the BSO+ATZ (1 mM)-treated cells, only 470 nm irradiation resulted in a decrease in the intracellular H2O2 level.
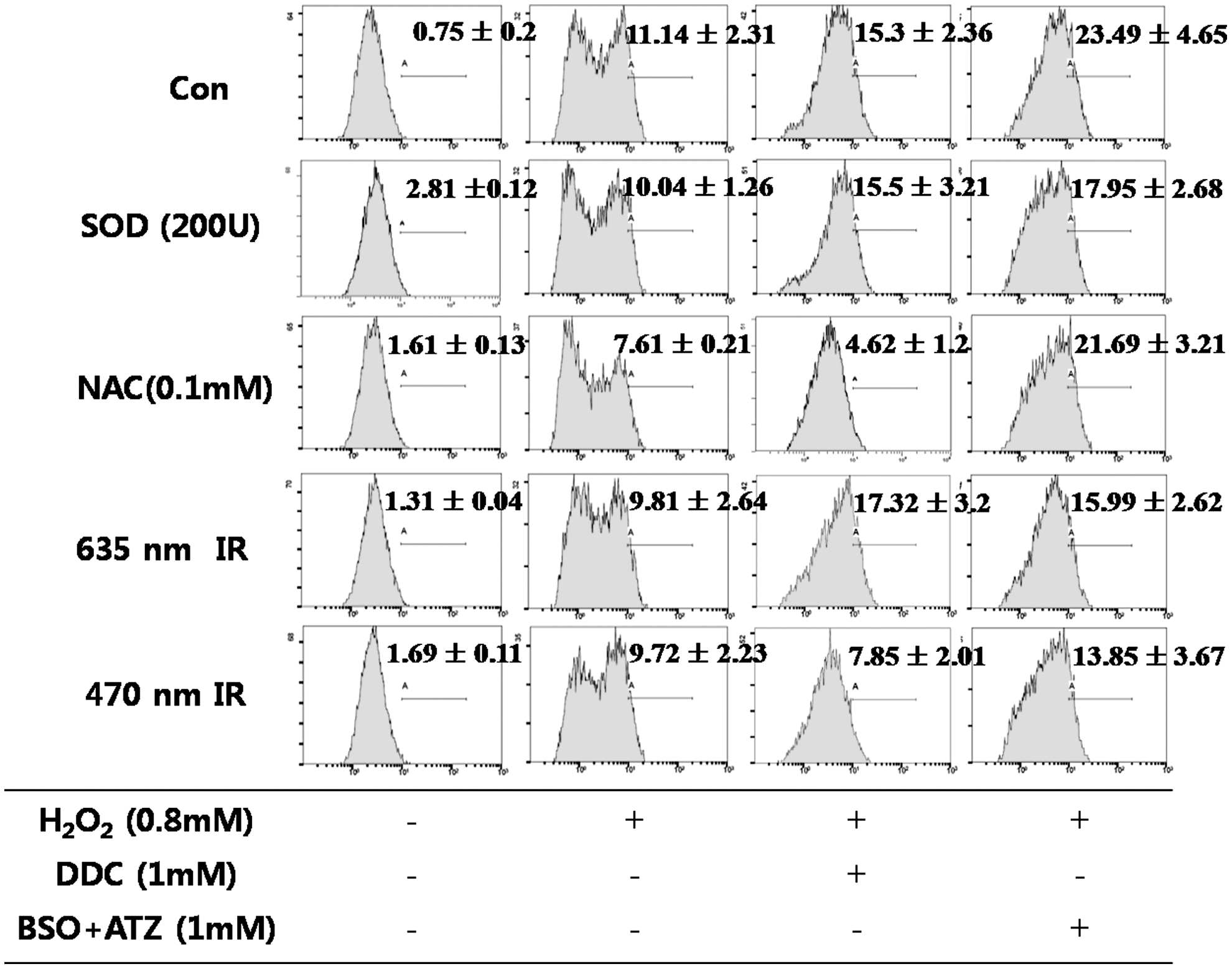
Flow cytometry analysis of 2′,7′-dichlorodihydrofluorescein (DCF) on H2O2 (0.8 mM) induced in the HaCaT cell line. Superoxide dismutase (SOD) (200U), N-acetyl-L-cysteine (NAC) (0.1 mM) treatment or 635 or 470 nm irradiation was performed for 1 h simultaneous with H2O2. Diethyldithiocarbamate (DDC) (1 mM) or buthionine sulfoximine and 3-amino-1,2,4-triazole (BSO+ATZ) (1 mM) were also added with H2O2 at the same time in each group. At 1 h after the treatment, flow cytometry was performed. The value indicate the means±SD (n=3).
ROS initiates the lipid peroxidation reactions that lead to loss of membrane integrity and cell death. Therefore, lipid peroxidation can represent oxidative stress caused by an imbalance between oxidants and antioxidants. MDA, a highly reactive bifunctional molecule, is an end product of membrane lipid peroxidation. The MDA level was increased by XXO treatment (Fig. 7). DDC treatment led to an increase in the MDA levels in the SOD- or NAC-treated groups, whereas the MDA levels in the 635 and 470 nm irradiated groups were reduced.
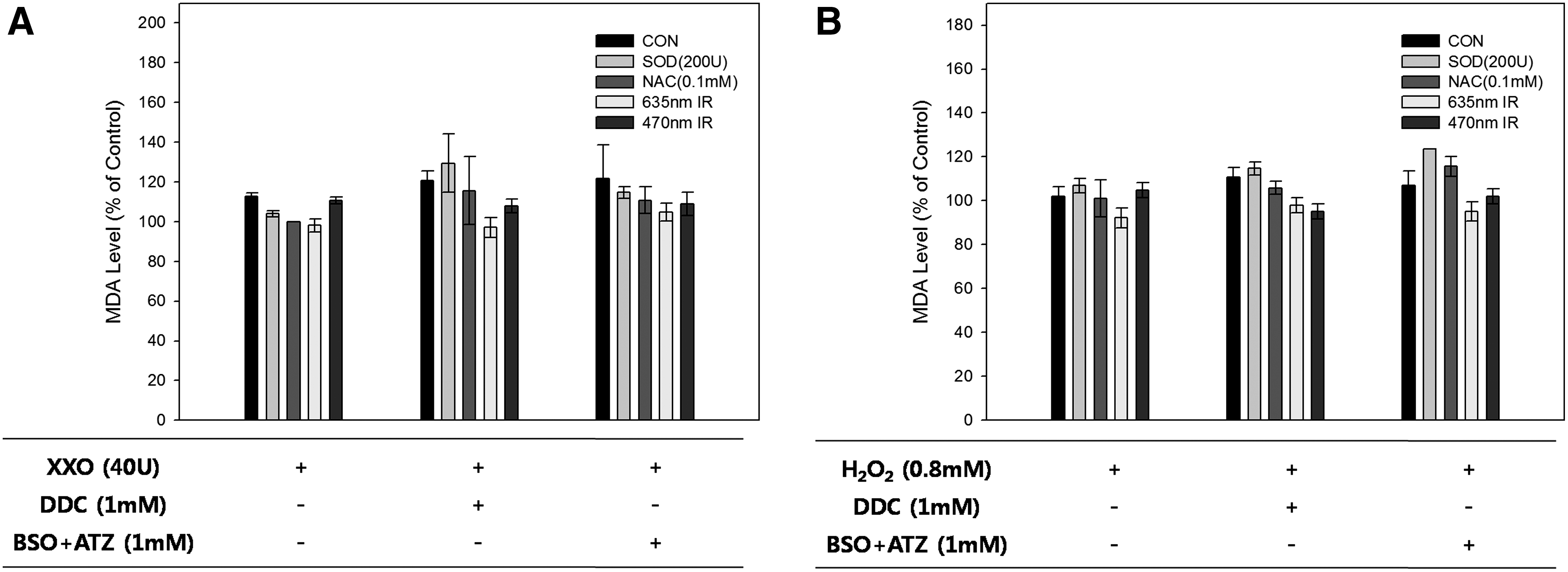
Lipid peroxidation assay by the malondialdehyde (MDA) level on xanthine/xanthine oxidase (XXO) (
Discussion
Human epithelial cells are exposed to exogenous and endogenous oxidizing agents that promote the generation of harmful ROS. 32,33 The intracellular concentration of ROS, in turn, is widely regarded as the critical factor in aging and in a number of degenerative diseases. 1
ROS are very short-lived molecules, and their production and removal is tightly regulated by enzymatic or nonenzymatic antioxidants. Oxidative stress can be defined as an increase in the reduction potential by the reactions described previoiusly, or as a decrease in the activities of the enzymes that prevent oxidative denaturation, including SOD, CAT, GP, GSH reductase (which regenerates GSH), and NADH-metHgb reductase. 2 An imbalance in these enzyme activities causes oxidative stress that can lead to cell death. 7
In the present study, superoxide anion generation in vitro by XXO led to a dose-dependent decrease in cell viability/proliferation and cell death. The addition of SOD and intracellular superoxide anion scavenging led to an increase in cell viability in the XXO-treated HaCaT cells. SOD, a ubiquitous metal-containing enzyme, converts superoxide anion to H2O2 according to the reaction shown below: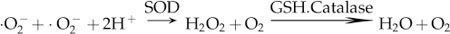
Irradiation at 635 nm also led to an increase in cell viability in the XXO-treated cells in a manner similar to that observed in the SOD-treated cells. Therefore, 635 nm irradiation appears to act as a superoxide anion scavenger. The superoxide anion can be reduced during electron transitions, in which elevated electron are induced to drop from higher energy levels to lower energy levels by UV irradiation. 34 –36 However, the superoxide anion exhibits absorption at 245 nm and does not have absorption band in the visible range of the spectrum. This means that the dissociation or detachment of the superoxide anion depends upon the intracellular antioxidants including the enzymatic or nonenzymatic antioxidant systems.
In the present study, after adding the SOD inhibitor, DDC to the XXO-treated cells and measuring the resultant intracellular superoxide anion levels, it was observed that the effects of superoxide anion were decreased by 635 nm light irradiation independent of the intracellular enzymatic antioxidant SOD. As a result, the amount of superoxide anion was decreased by SOD treatment and SOD did not have any effect on the intracellular superoxide anion levels in the DDC (1 mM)-treated cells. Irradiation at 635 nm led to a decrease in the superoxide anion levels regardless of the presence or absence of DDC, which suggests that 635 nm irradiation removes the superoxide anions in a SOD-independent manner. The scavenging of superoxide anion by 635 nm irradiation led to a decrease in oxidative stress and lipid peroxidation in the presence or absence of DDC.
In the cells treated with H2O2, the level of superoxide anions was decreased by 635 nm irradiation. NAC treatment resulted in a slight decrease in the level of superoxide anions but it could not reduce the level of superoxide anions in the BSO+ATZ-treated cells (BSO, ATZ are the inhibitors of GSH and CAT, respectively). The concentration of intracellular H2O2 was decreased by 635 nm irradiation in the XXO-treated cells, and, consequently, the superoxide anion was scavenged by 635 nm irradiation, even though H2O2 could not be removed via the same pathway that was involved in superoxide anion scavenging. Treatment of cells with H2O2 also led to a dose-dependent decrease in cell viability. H2O2 is frequently used as the oxidative stimulus in order to examine the various effects of oxidative stress. Treatment with exogenous H2O2 led to a significant increase in the intracellular hydrogen peroxide levels. NAC treatment and the subsequent decrease in the intracellular H2O2 levels led to an increase in cell viability in the H2O2-treated cells. NAC is a GSH precursor, which is a H2O2 scavenger causing a decrease in the cytostatic and cytotoxic effects. 18 The effect of H2O2 on the viability of cells was attributed to the finely regulated detoxification of intracellular H2O2: NAC can increase the cell viability. However, 635 nm irradiation did not alter cell viability compared with that by NAC. It means that 635 nm irradiation led to scavenging of superoxide anion rather than that of H2O2.
If that was the case, how could 635 nm irradiation reduce the oxidative stress independent of SOD? In general, the elevation of antioxidants was evident in various studies after LLLT. Application of LLLT 1 h after occlusion of the femoral artery to the gastrocnemius muscle still had a similar effect on the creatine-phosphokinase activity, as did application of a laser immediately after the onset of ischemia. 37 LLLT enhances the production of ATP in cells, preserves the mitochondria, alleviates the oxidative stress, and elevates ATP production in an ischemic heart. 22 This phenomenon may, in turn, prevent the degeneration of muscle cells under severe hypoxic conditions in an ischemia-reperfusion injury. LLLT wavelengths that have been reported to have stimulating effects in biological systems include blue (400–500 nm), green (510–550 nm) yellow (560–590 nm), red (600–700 nm) and near-infrared (710–1100 nm). 22 Different chromophores have been proposed to be operating as photoacceptors depending upon the wavelengths chosen and the precise biological system under investigation. Blue and green have been proposed to be absorbed by flavins and flavoproteins. Yellow and red light have been proposed to be absorbed by porphyrins. Red and near infrared light have been proposed to be absorbed by cytochrome c oxidase (unit four in the mitochondrial respiratory chain). The absorption spectra obtained for cytochrome c oxidase in different oxidation states were recorded and found to be very similar to the action spectra for the biological responses to light. 38,39 Cytochrome c oxidase is known to be major component of the respiratory chain for producing ATP. Increases in ATP are one of the most often observed changes in vitro after LLLT, and increased cytochrome c oxidase activity would explain the increased ATP levels. 40 Particularly in the stressed or hypoxic cells, nitric oxide (NO) produced in the mitochondria can inhibit respiration by binding to cytochrome c oxidase and competitively displacing oxygen. 41 In a previous research, NO donor treatment in SH-SY5Y neuroblastoma cells led to ROS generation resulting in mitochondrial-dependent apoptotic progression. However, 635 nm irradiation suppressed ONOO− synthesis via the dissociation of ROS, led to a decrease in the oxidative stress, and inhibited the apoptotic pathway. 24 This suggests that light absorption displaced the NO and allowed the cytochrome c oxidase to recover and cellular respiration to resume, which would explain the decrease in oxidative stress in many of the observations made in LLLT. The cells or tissues damaged as a result of stress or hypoxia are likely to respond more to LLLT than normal cells and tissues, and their cytochrome c oxidase is more likely to be operating at suboptimal level because of NO inhibition.
Conclusions
In conclusion, 635 nm LED irradiation can help remove the intracellular superoxide anion in SOD-inactivated HaCaT cells, thereby resulting in a decrease in lipid peroxidation, followed by an increase in cell viability. Therefore, 635 nm irradiation can help alleviate oxidative stress that is independent of the activities of the intracellular enzymatic antioxidants, such as SOD, GSH, and CAT.
Footnotes
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2010-0007676).
Author Disclosure Statement
No competing financial interests exist.