
Abstract
Introduction
Mitochondria are a major source of chronic reactive oxygen species (ROS), which impair lipids, protein, and mitochondria DNA, leading to the dysregulation of mitochondrial function. 9 –12 Research has indicated that mitochondrial damage may be central to the impairment of the cells of the regulatory systems – such as the nervous, endocrine and immune systems – and to the communication between them. 13 –15 Therefore, insulin-resistant type 2 diabetes mellitus, obesity, hyperglycemia, hyperlipidemia, and atherosclerosis, which are prevalent in longstanding spinal cord injury (SCI) 16,17 , may be linked to mitochondrial dysfunction.
Although oxidative stress clearly plays an important role in the progression of SCI, most of the experiments conducted to date on oxidative stress and antioxidant defenses occurring at the injury site have been on animal models, 18 –20 and there is still a scarcity of research on mitochondrial function, oxidative stress, and antioxidant intervention on human serum models. Moreover, although there is a considerable body of experimental data from research on humans that demonstrates the positive effects of ILIB on biostimulation, there is still a lack of randomized, sham-control studies to indicate its influence on different metabolic pathways.
Given that the emerging potential applicability of ILIB to biomedical pathologic processes has some way to go in being rigorously trialed with human subjects using appropriate methodologies, this study is a key piece of work in testing some of these in-principle clinical applications. In consideration of these, the rationale and methodology of this study were aimed at filling some of the gaps in the existing research: specifically, by characterizing temporal changes, through a single-blinded, randomized controlled methodology to investigate the effects of ILIB on mitochondrial behavior, oxidative stress biomarkers, and serum biochemical changes in chronic SCI human subjects; and, more broadly, by contributing to the body of knowledge of ILIB and its potential therapeutic effectiveness.
Methods
Participants
Thirty-six individuals were included in the study, of whom 12 were healthy controls (7 males and 5 females) and 24 (18 males and 6 females) had chronic SCI resulting from trauma, and were inpatients or outpatients of a regional SCI rehabilitation program. The SCI subjects submitted to a physical examination and routine biochemical and hematologic laboratory tests, including lipid profile, glucose, urea, creatinine, enzymes aspartate aminotransferase (AST), alanine aminotransferase, and hemograms (red blood cell [RBC], WBC, and platelet counts, and hemoglobin levels), and were receiving standard treatment, including rehabilitation therapy tailored to their specific needs.
Admission eligibility for the study included having an American Spinal Injury Association (ASIA) impairment scale classification of A, B, C, or D, a neurologic injury level between the C3 and L2 spinal cord levels, and medical stability. Subjects with communicable diseases, or active health problems (malaise, flu, fever, cough, change in urine status), or who were taking anti-inflammatory medications at the time of the study, were excluded. The subjects' clinical and demographic profiles are listed in Table 1. Prior to participation, all individuals signed an informed consent form. The investigation was in accordance with the Declaration of Helsinki Principles and conducted under the supervision of the local institutional review board protocols for human subjects and Occupational Health and Safety Administration regulations.
ASIA, American Spinal Cord Injury Association.
Allocation procedures
To achieve our primary research goal, the SCI participants were randomized into either a study or a sham group using a computer-generated random order. The randomization list was supervised by one of the investigators who had no contact with the participants until the group assignment was revealed. Further, the group assignment remained unrevealed to our laboratory staff throughout the experimental period.
Intravascular low-level laser irradiation
We adopted the helium-neon (He-Ne) laser (YJ-ILIB-5, Bio-ILIB Human Energy Ltd, Taiwan) as the conditioning protocol in this study. Each participant sat on a semi-inclined sofa. A 24-gauge intravenous catheter was used for an elbow venous puncture (phlebotomy). The intravenous needle was then replaced by a fiberoptic needle inserted into the inner cannula of the intravenous catheter. The other side of the fiberoptic needle was connected to the He-Ne laser illuminator. The irradiation parameters involved in the ILIB application are shown in Table 2. For the sham control subjects, ILIB was provided without any laser power (0.0 mW). In all other respects, the program was identical for both SCI groups.
ILIB, intravascular laser irradiation of blood.
Biochemical analysis
Blood samples were taken from the SCI study group for biochemical analysis after a 12 h overnight fast at the baseline, the 10th, and the 15th days during the ILIB intervention period, and then after the end of the ILIB intervention, on the 30th day (the 45th day after the initiation of ILIB).
Isolation of WBCs from peripheral blood
Each participant provided a sample of fresh peripheral heparinized venous blood, which was centrifuged at room temperature at 3000 g for 10 min. We then transferred the buffy coat layer to 1.5 mL of Eppendorf, and washed it with RBC lysis buffer (containing 155 mM NH4Cl, 10 mM KHCO3, and 0.1 mM EDTA, pH 7.4) and incubated it at room temperature for 10 min. Each sample was centrifuged at 1500 g for 5 min at room temperature, and the cell pellets were resuspended in 1 mL of phosphate-buffered saline (PBS, pH 7.4) in preparation for the subsequent experiments. A hemocytometer was used to count the WBCs. The total number of viable WBCs was determined according to their exclusion of 0.4% Trypan Blue (Sigma-Aldrich).
Measurement of mitochondrial DNA (mtDNA) copy number from leukocytes
To measure the copy number of the mtDNA in the leukocytes, we followed a real-time polymerase chain reaction (PCR) technique according to a method described earlier by Lee et al. 21 , using a LightCycler-FastStart DNA Master SYBR Green I Kit. An aliquot of 50 ng of DNA was amplified in a 10 μL reaction capillary with 0.25 μL of each primer (20 μM), 5 μL of LightCycler SyBr Green mixed reagent and 3.5 μL of PCR-grade H2O. We established the real-time PCR condition as: 10 min start at 95°C, a subsequent 45 cycles of 20 sec each at 95°C; 20 sec at 62°C; and 20 sec at 72°C. At the end of every extension phase, we measured the fluorescence intensity of the PCR product at 79°C. The sequences of primers used to amplify the mitochondrial DNA (mitochondrial NADH dehydrogenase 1 gene, ND1) were: ND1R, 5’-ACCATTTGCAGACGCCATAA-3’ and ND1F, 5’-TGAAATTGTTTGGGCTACGG-3’. The sequences of primers used to amplify the nuclear DNA (18S rRNA) were: 18S1546F, 5’-TAGAGGGACAAGTGGCGTTC-3’ and 18S1650R, 5’- CGCTGAGCCAGTCAGTGT-3’. We normalized the crossing points (Cp) in quantitative PCR curves between the ND1 and 18S rRNA genes using RelQuant software (Roche Applied Sciences, Mannheim, Germany) to measure the relative mtDNA copy number. To check the individual experiments for the quantification of mtDNA copy numbers performed by RT-PCR, we used DNA from CCD966SK normal skin fibroblasts as the internal standard for calibrating the relative mtDNA copy numbers in each sample from each individual RT-PCR run. The Ct (ND1 to 18S) for CCD966SK was therefore defined as 1.0, and we were therefore able to calculate the relative mtDNA copy number for each sample in each individual experiment set. In this way, we precluded any errors from arising from an individual quantitative PCR run. The PCR product for ND1 is 108 base pairs and for 18S, 105 base pairs. The amplification of the PCR product was followed by a melting curve analysis to monitor the homogeneity of the amplicons. At the final RT-PCR step, melting curve analysis was performed, consisting of a denaturation at 95°C followed by a renaturation step by a slow melt approaching 68°C with a ramping rate of 0.2°C/sec with continuous data acquisition.
Measurement of total cellular ATP in WBCs
We measured the total cellular ATP content of the WBCs using the Bioluminescent Somatic Cell Assay Kit (Sigma-Aldrich, St. Louis, MO) in accordance with the manufacturer's instructions and our experience. 22 Once the WBCs were separated from the whole blood samples, we released the total cellular ATP contents by mixing an aliquot of 50:l WBC suspension with 150:l Somatic Cell Releasing Reagent. One half (100:l) of the mixture was then transferred into a black OptiPlate™-96F 96-well plate (Packard Biosciences, Groningen, The Netherlands) containing 100:l ATP Assay Mix. We then measured the luminescence intensity using the Victor2 TM 1420 multilabel counter machine (PerkinElmer Life Sciences, Inc., Boston, MA). The ATP sample standards in the range of 0–300 pmoles were used, and each ATP value was normalized with WBC numbers.
Analysis of malondialdehyde (MDA) in blood serum
MDA in the participants' blood serum was measured using an MDA assay kit (MDA-586, OxisResearch, Inc. Portland, OR) with spectrophotometric analysis. Measurements were made according to the manufacturer's instruction and our own experience. 23 The assay was involved in the reaction with a chromogenic regent N-methyl-2-phenylindole (NMPI) to form an intensely colored carbocyanine dye with a maximum absorption of 586 nm. The methodology we used here is specific to MDA, rather than to other lipid peroxidation byproducts such as 4-hydroxy alkenal, because under the same experimental conditions they are unable to produce significant absorbance at 586 nm. Using this assay, an MDA standard curve was established at the concentration range of 0–50 μM.
Determination of total antioxidant capacity (TAC) of blood serum
The TAC of the participants' serum was measured using a PAO kit (Japan Institute for the Control of Aging, Nikken SEIL Co., Fukuroi, Japan) in accordance with the manufacturer's instruction and our own experience. 22 An aliquot of 100 μL of diluted serum from each participant was mixed with 50 μL of Cu2+ solution, which can be reduced to form Cu+ in correspondence with the antioxidant status of serum after 5 min of incubation at room temperature. We then added an aliquot of 50 μL of a chromogenic solution to react with the Cu+ ion. The absorbance at 490 nm was measured immediately. We determined the TAC levels of the serum by reducing the Cu2+ solution with corresponding concentrations of uric acid. To gauge the antioxidant power of the samples, we multiplied the corresponding uric acid concentration by 2189 (1 mM uric acid=2189 μ mole/L copper reducing power).
Statistical analysis
Statistical analysis of the processed data was performed with commercial software (SPSS 17.0 for Windows; SPSS Inc.). Descriptive statistics were used to elucidate a set of known data in a clear and concise manner, as in term of its mean and variance. Herein we used descriptive statistics to summarize a data set of subject characteristics and display major findings. The Wilcoxon signed-rank test was used to compare the biochemistry changes across time versus the baseline values of the study group. One-way analysis of variance (ANOVA) was used to analyze the baseline data of MDA, TAC, mtDNA and total cellular ATP in WBC among the normal, sham, and study groups. A two-way repeated measures ANOVA was used to analyze antioxidant activity of the body and mitochondrial activity with time (baseline, day 1, and day 15 post-intervention) as the within- and between-group factor. When an intervention between group and time was found, post-hoc tests were performed to determine the real within- and between- group differences. The level of significance was set at 0.05.
Results
Across all demographic indicators, there were no significant differences between the sham and study groups (Table 1). The complete blood cell counts, liver transaminases, blood urea nitrogen, creatinine, creatinine phosphokinase, and urine pH were unchanged before and after the intervention. However, low-density lipoprotein (LDL) in the study group showed a significant decrease at both days 10 and 15, with significantly higher high-density lipoprotein (HDL) found at day 45 as compared with the baseline measurement (Table 3).
Values were Mean±SD.
Wilcoxon signed-rank test was used to compare the data changes across time versus baseline in study group.
p <0.05.
ILIB, intravascular laser irradiation of blood; TG, triglycerides; HDL, high-density lipoprotein; LDL, low-density lipoprotein; IL, interleukin; TNF, tumor necrosis factor.
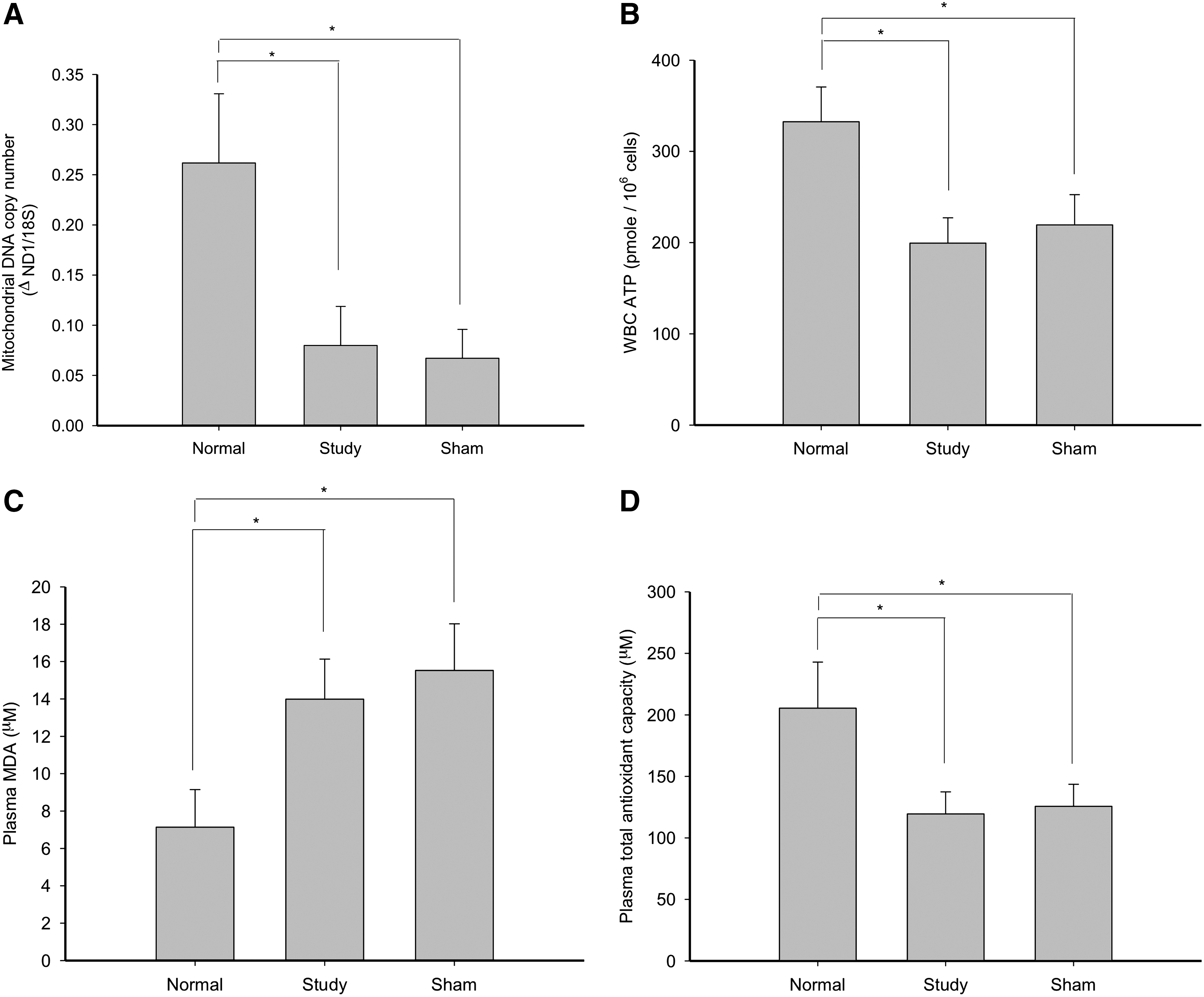
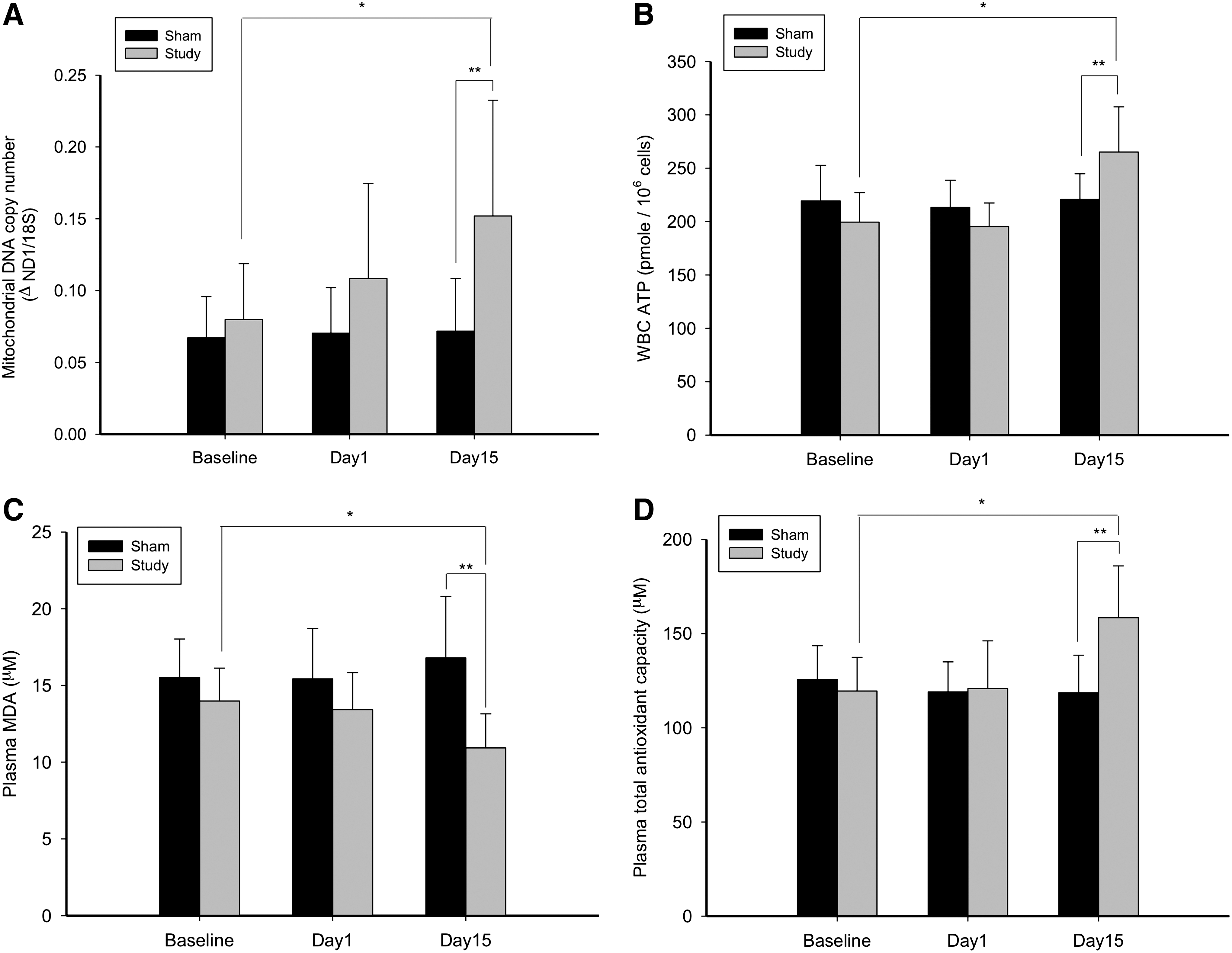
The baseline measurements revealed that there were significant differences among groups. The normal, aged-matched subjects has a significantly higher mtDNA copy number, WBC ATP synthesis, and TAC, and less MDA than did the study and sham subjects of the SCI group (Fig. 1). At day 15 of the ILIB intervention, the study group was found to have a significantly higher mtDNA copy number, WBC ATP synthesis, and TAC with significantly reduced MDA than did the sham group (Fig. 2). The intragroup comparisons for the study group revealed that the mtDNA copy number, WBC ATP synthesis, and TAC at day 15 were significantly increased, with significantly reduced MDA (Fig. 2). The intragroup comparisons for the sham group, by contrast, demonstrated no statistical differences for any biomarkers.
Discussion
Use of He-Ne laser (632.8 nm) to promote functional regeneration of mitochondria and pro-oxidant/antioxidant equilibrium in living bodies is common; moreover ILIB with He-Ne laser has been applied in aid of treatment of acute cerebral infarction, rheumatoid arthritis, arteriosclerosis of the leg, and redifferentiation therapy of malignant tumors. 24 –28 However, over the past 30 years, there has been a small, but growing, body of clinical studies that has been challenging the status-quo belief with regard to the therapeutic effectiveness of ILIB, and at present there are no generally accepted practical ILIB strategies. Expectations that ILIB can reduce oxidative stress in chronic SCI patients have to date been based on findings that mitochondrial photostimulation enhances ATP synthesis and contributes to redox signaling in the clearing of free radicals; 29 and on the assumption that mitochondria are the primary targets of red light radiation with 632.8nm wavelength of whole cells. 5 –7,29 This study is the first to rigorously characterize temporal changes in mitochondrial function and antioxidant activity following ILIB therapy in chronic SCI subjects. Although the overall procedure required 15 days of 1 h 4 mW He-Ne laser ILIB intervention sessions, all of the subjects were able to tolerate the whole procedure with no adverse effects, which is consistent with and reinforces previous findings. 4
The main findings of this study were: (1) that chronic SCI patients were indisputably in states of mitochondrial damage and oxidative stress in comparison with normal subjects; (2) there was a significant increase in the mtDNA copy number, WBC ATP synthesis, and TAC, with significantly reduced MDA production in the study group as compared with the sham group at day 15 of ILIB intervention; 3) there was a significant increase in the mtDNA copy number, WBC ATP synthesis, and TAC, with significantly reduced MDA production in the study group participants at day 15 of ILIB intervention as compared with their baseline values; and 4) there was significant decrease in LDL of the study group participants at day 10 and day 15 of ILIB intervention, with significant increase in their HDL as compared with their baseline measurements.
In vivo data are still limited concerning the mechanism by which secondary SCI deterioration occurs following initial primary injury. However, several lines of evidence have suggested that ROS formation and the consequent development of oxidative stress are important events associated with SCI. 20 ROS are difficult to measure directly as they are highly reactive and short lived. Indirect measures such as MDA and other biomarkers are therefore generally used to predict the amount of ROS or the extent of oxidative stress occurring in the SCI subjects. 30 The baseline findings of this study, which established that chronic SCI subjects had significantly higher baseline MDA content, lower TAC, and impaired mitochondrial metabolic activities than did normal subjects, support the abovementioned evidence of the development of ROS formation and oxidative stress in secondary SCI deterioration. The data recorded for the chronic SCI study and sham groups of this research also confirm previous findings that the application of a He-Ne laser could improve mitochondrial activities (DNA and ATP synthesis) in WBC and reinforce antioxidant defenses in the blood serum of chronic SCI patients. Although the underlying mechanisms have yet to be clearly elucidated, our findings do point to a potential link between photostimulation and mitochondrial proliferation and/or cellular homeostasis. As ILIB is known to generate low levels of ROS that can upregulate cellular antioxidant defenses and improve cell signaling, 31 it is feasible that TAC increases as a result of ILIB intervention in compensation for a beneficial, low-level increase in cellular ROS, and that serum MDA decreases because the proximate membrane protein-glutathione peroxidase 4, which is responsible for lipid peroxidation, is downregulated. 32 Our results here support the concept that ROS act as key secondary messengers contributing to ILIB-induced biologic outcomes. As the prevalence of adiposity soars in chronic SCI patients, appreciation of the close links between obesity and disease increases. The strong relationships between lipid metabolism and poor health outcomes, including cardiovascular disease, diabetes, and metabolic syndrome, mandate clarification of cholesterol constituent changes in chronic SCI patients. It is of interest to note that the LDL of the study group decreased during the application period of ILIB and that its HDL was subsequently raised. The redistribution of cholesterol was strongly evidenced in concordance with markedly reducing serum MDA and increasing serum TAC. We consequently propose that ILIB plays a central role in the regulation of lipid metabolism, which improves LDL catabolism, reduces the oxidation of LDL, and increases HDL levels. This beneficial effect of ILIB on LDL catabolism could be partially related to the upregulation of LDL receptor expression. 33 –35 The mechanism by which ILIB increases HDL is not fully understood, but is likely related to increased expression of lipoprotein lipase or decreases in apolipoprotein (Apo) A-I clearance, 34,36 which may augment the biosynthesis of HDL. 34 It is clear that in vivo, HDL can inhibit the oxidative modification of LDL. In addition to the antioxidative Apo A-I, glutathione and paraoxonase 1 are all HDL-associated enzymes, which inactivate lipid peroxides and other oxidized LDL lipid components. 36,37
Limitations
There were several limitations to the methodology of this study. The lack of continuous monitoring of mitochondrial function and of antioxidant effects before and after each ILIB session made it impossible to examine the precise moment at which mitochondrial proliferation and cellular homeostasis were reached and their duration. However, our decision to administer ILIB 15 times in this protocol was in accordance with previous studies and suggestions by Weber et al. 4 Further research on different laser energies and application settings would advance these findings and identify optimal treatments for adults with chronic SCI. This study focused on the proof that oxidative stress in chronic SCI subjects is alleviated after exposure to He-Ne laser. The relationship between the generation of antioxidant defenses and clinical motor/sensory improvement was beyond the scope of this study comprising a small number of participants undergoing a short-term period of treatment. The achievement of significance for every study group participant of this study firmly indicates the value of further exploring and testing its findings in a large-scale trial of ILIB intervention as part of longer-term treatment of chronic SCI patients. We expect that more alleviation of oxidative stress in longer-term trials may have better results in neurologic and metabolic recovery of SCI, as oxidative stress, in addition to ROS, is a key element in the pathophysiology of SCI. 18 –20
Conclusions
The baseline findings of this study across normal and chronic SCI patients established that our chronic SCI patients were in states of mitochondrial damage and oxidative stress. ILIB intervention was found to significantly increase subjects' mtDNA copy number, WBC ATP synthesis, and TAC; significantly reduce MDA production; significantly decrease LDL; and significantly increase HDL. Our findings on human subjects have therefore reinforced the body of research that contends that ILIB therapy promotes mitochondrial DNA or total cellular ATP synthesis, antioxidant activity, and the reduction of lipid peroxidation and dyslipidemia generation, contributing to the alleviation of chronic conditions, such as chronic SCI. Given this precedent and the alleviation of oxidative stress and ROS-scavenging capacity, we believe that this clearly signals ILIB as a potential addition to the current armament of treatment for those with SCI.
Footnotes
Author Disclosure Statement
No competing financial interests exist.