
Abstract
Introduction
After axonotmesis, the inflammatory process is triggered; the proximal stump axons degenerate and distal fibers from the lesion undergo Wallerian degeneration between 48 and 96 h after nerve transection. Particularly, but not exclusively, after axonotmesis, pro-inflammatory cytokine mobilization and increased activity of induced nitric oxide synthase (iNOS), which results in inflammatory cell recruitments to the affected site, occurs, leading to increased production and releasing of several pro-inflammatory mediators, which ultimately reduces significantly the nerve recovery process. In addition, there are also myelin debris degradation and other structural proteins that finally inhibit axonal growth. 3,4
Simultaneously, substances known as neurotrophic factors (e.g., nerve growth factor [NGF], brain-derived neurotrophic factor [BDNF], and neurotrophin [NT]-3 and NT-4/5) play an important role in promoting neuronal survival and axonal regeneration. 5,6 Although neurotrophic factors can be produced spontaneously after axonotmesis, it is known that complete nerve regeneration depends upon trophic conditions highly favorable to both neuronal protein synthesis and neurotrophic factor expressions. 7 –10
Low-level laser therapy (LLLT) in general has been used to promote analgesic 11 and anti-inflammatory 12 actions, wound healing, 13 and neuronal regeneration. 14 –17
Theoretically, LLLT, through its photochemical and photobiological effects produced at the cellular level, 18 could induce both trophic conditions and the inflammatory process inhibition that is necessary for relevant nerve regeneration. 19
Based on the data presented previously and some good preliminary results in our laboratory, we started an investigation to verify if LLLT could promote increases in neurotrophic factor expressions, in addition to inhibiting the inflammatory process.
Methods
All experiments described here complied with all the precepts advocated by Brazilian College of Animal Experimentation (COBEA), and were approved by the Local Ethical Committee (Protocol. 001/10). Thirty-six young adult male Wistar rats were used (weight: 250–350 g) supplied by Universidade Regional de Blumenau breeding.
Animals were housed in individual cages in an isolated room with constant temperature at 22°C and with a 12 h photoperiod (light period from 07 a.m. to 07 p.m.) and free access to food and filtered water.
Axonotmesis
To perform axonotmesis, animals were anesthetized, via intraperitoneal injection, with a xylazine (10 mg/kg) and ketamine hydrochloride (100 mg/kg) cocktail in compliance with aseptic and antiseptic requirements. After reaching the desired depth level of anesthesia (absence of the withdrawal reflex after pinching the interdigital folds), an incision was made at the thigh level of the right hind paw to crush 5 mm before the sciatic nerve bifurcation during 30 sec using the maximum pressure level (three clicks) of a hemostatic forceps (according to Bridge et al.) 20 (Fig. 1).
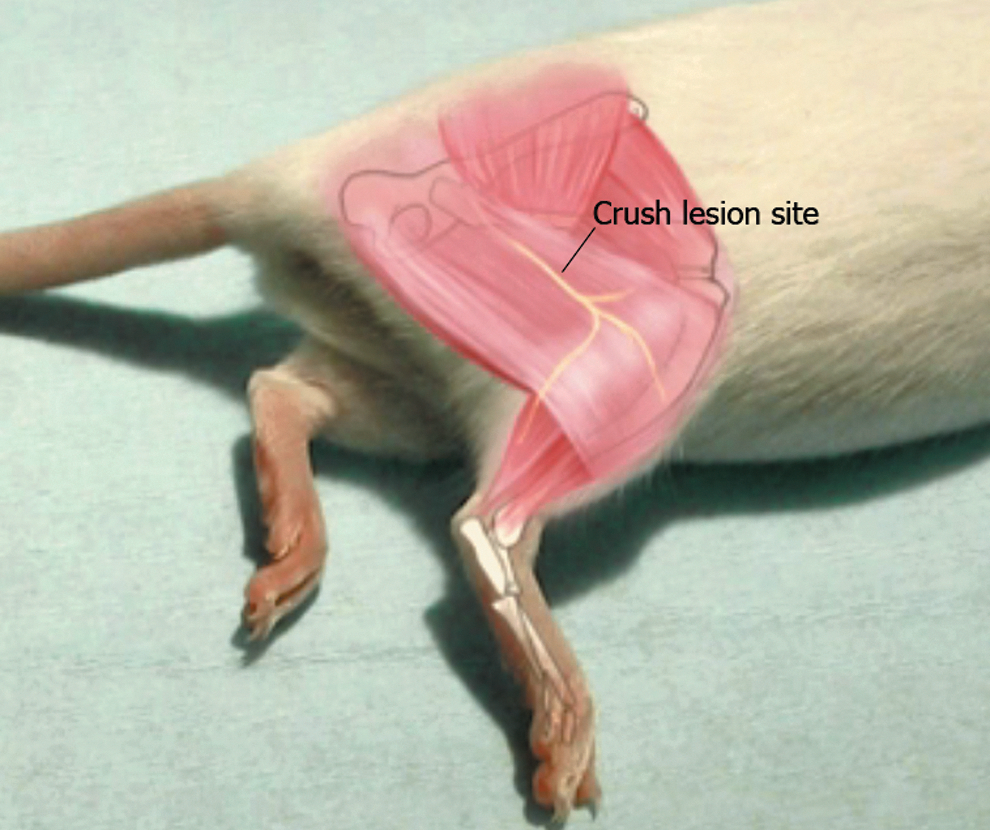
Representation of the right sciatic nerve crush injury level.
Grouping
Thirty-six rats were divided into three groups, consisting of 12 animals each (6 experimental and 6 control). The control animals were subjected to nerve crush, but did not receive LLLT. The experimental groups underwent transcutaneous LLLT (10 consecutive different spots on the nerve path: four points above, five points below, and one point on the lesion epicenter) on a daily basis, beginning 24 h after crush injury. Detailed LLLT information is described in Table 1. Animals from group 1 underwent to LLLT for 7 days; those from group 2 underwent to LLLT for 14 days; those belonging to group 3 underwent to LLLT for 21 days. The laser parameters were adjusted and provided by the equipment manufacturer.
mRNA expression
After the experiments' final phase, animals were killed by an overdose of anesthesia to remove samples from the sciatic nerve lesion epicenter which, were submitted for polymerase chain reaction with reverse transcription (RT-PCR) analysis. 21
The specific primers to verify the mRNA expression of neurotrophic factors (BDNF, NGF and NT-3) and iNOS, were synthesized based on their gene sequence referred in GenBank (Table 2). The PCR products were analyzed by electrophoresis on agarose gel (1%) (Bio America Inc., Miami, FL), stained with ethidium bromide, and visualized over UV light (Vilber-Lourmat®, Photo Capt-System). The relative product concentration of RT-PCR for BDNF, NGF, NT-3 and iNOS were compared with GAPDH (constitutive gene process control amplification), and all reactions were performed in a thermocycler (BioCycle MJ96+, Biosystems, São José dos Pinhais, PR, Brazil). The bands were detected and analyzed using Photo-Capt version 12.4 for Windows®, which quantified its intensity.
RT-PCR, polymerase chain reaction with reverse transcription; bp, base pair; GADPH, glyceraldehyde 3-phosphate dehydrogenase; BDNF, brain-derived neurotrophic factor; NGF, nerve growth factor; NT-3, neurotrophin-3; iNOS, induced nitric oxide synthase.
Data analysis
Data related to experimental and control groups were collected at three different times: 7 (G1), 14 (G2), and 21 (G3) days after nerve crush injury, and were expressed as mean±SEM. The InStat program (GraphPad Software) was used for the statistical analysis. The data were examined by ANOVA followed by the Tukey post-hoc test to determine differences among groups, and the results were considered significant when p<0.05. For the graph constructions, GraphPad Prism program version. 3.0 was used.
Results
It was impossible to observe any changes in NT-3 expression, either spontaneously or after LLLT. For practical reasons no data will be presented here.
On the other hand, as can be seen in Fig. 2, BDNF expression began to appear soon after axonotmesis. It was possible to notice that the mechanisms related to BDNF expression seemed to respond quickly to nerve aggression, and tended to increase through time.
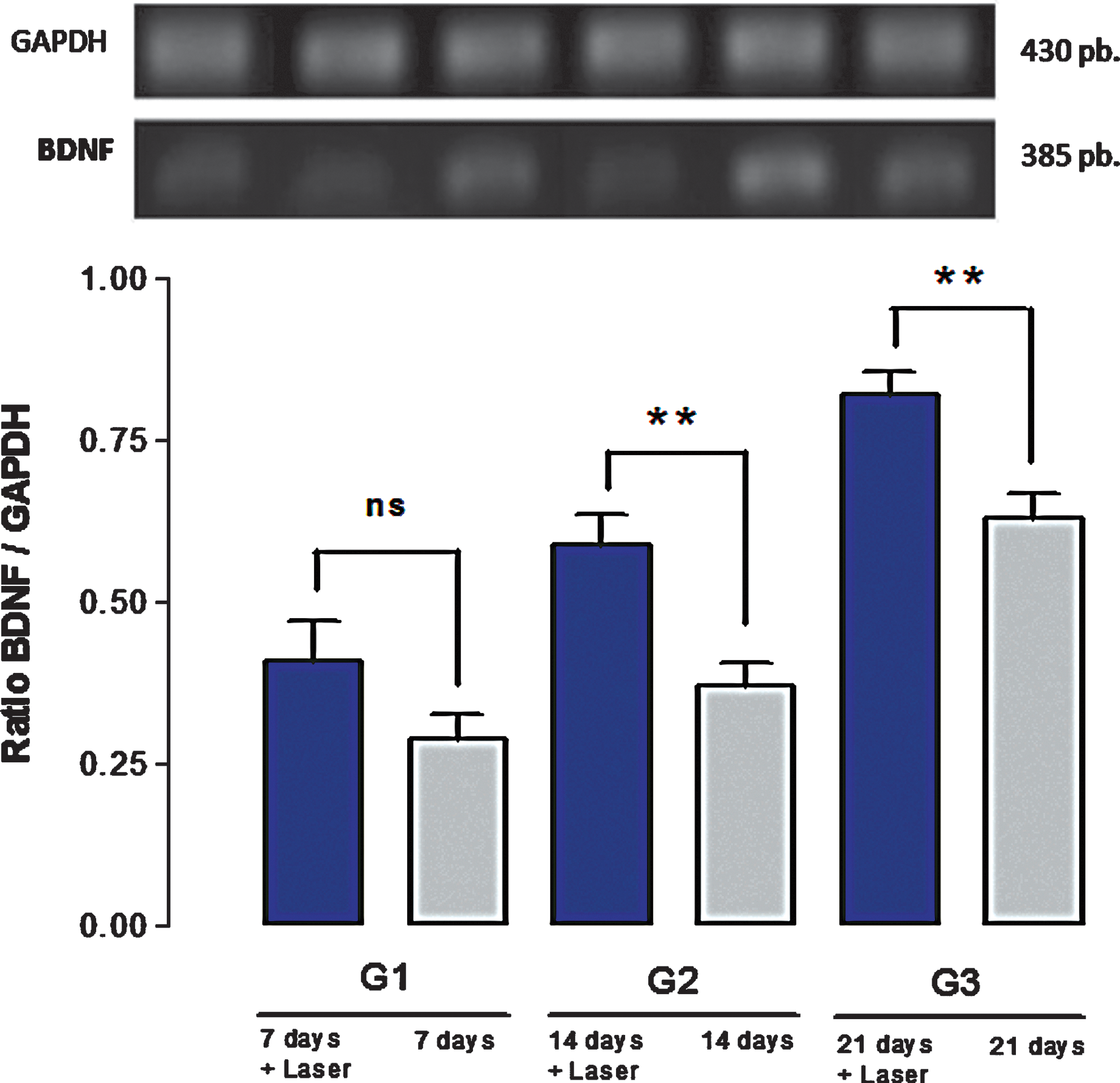
Brain-derived neurotrophic factor (BDNF) expression over 21 days. The upper portion illustrates, by comparison, the GAPDH and BDNF expression bands. Below, there are columns corresponding to BDNF expression with (irradiated) and without (nonirradiated) low-level laser therapy (LLLT) at days 7, 14, and 21 after nerve crush injury. **Significant differences, p<0.01.
More important are the findings related to BDNF expression after LLLT applications. Figure 2 shows an enhancing effect of laser on BDNF expression after axonotmesis, reaching an expression peak at the 21st day of consecutive LLLT applications. On the 7th day, it was observed that the irradiated group showed greater BDNF concentrations than did the control group, although this difference was not statistically significant. However, observing both G2 and G3 data, it was possible to notice that LLLT was able to promote significant increases in BDNF expression (Fig. 2) when compared with the respective control group. This behavior shows that LLLT enhances BDNF expression's natural cadence.
NGF expression followed the same trend observed in BDNF expression, that is, fast and immediate responses after axonotmesis followed by progressive increases until the 21st days after sciatic nerve crush (Fig. 3). However, NGF expression profile is slightly different. NGF expression apparently happens tonically during the first 14 days after axonotmesis followed by progressive increases until the 21st day. Additionally, a closer inspection of both Figs. 2 and 3 demonstrates that NGF expression is nearly twice BDNF expression.
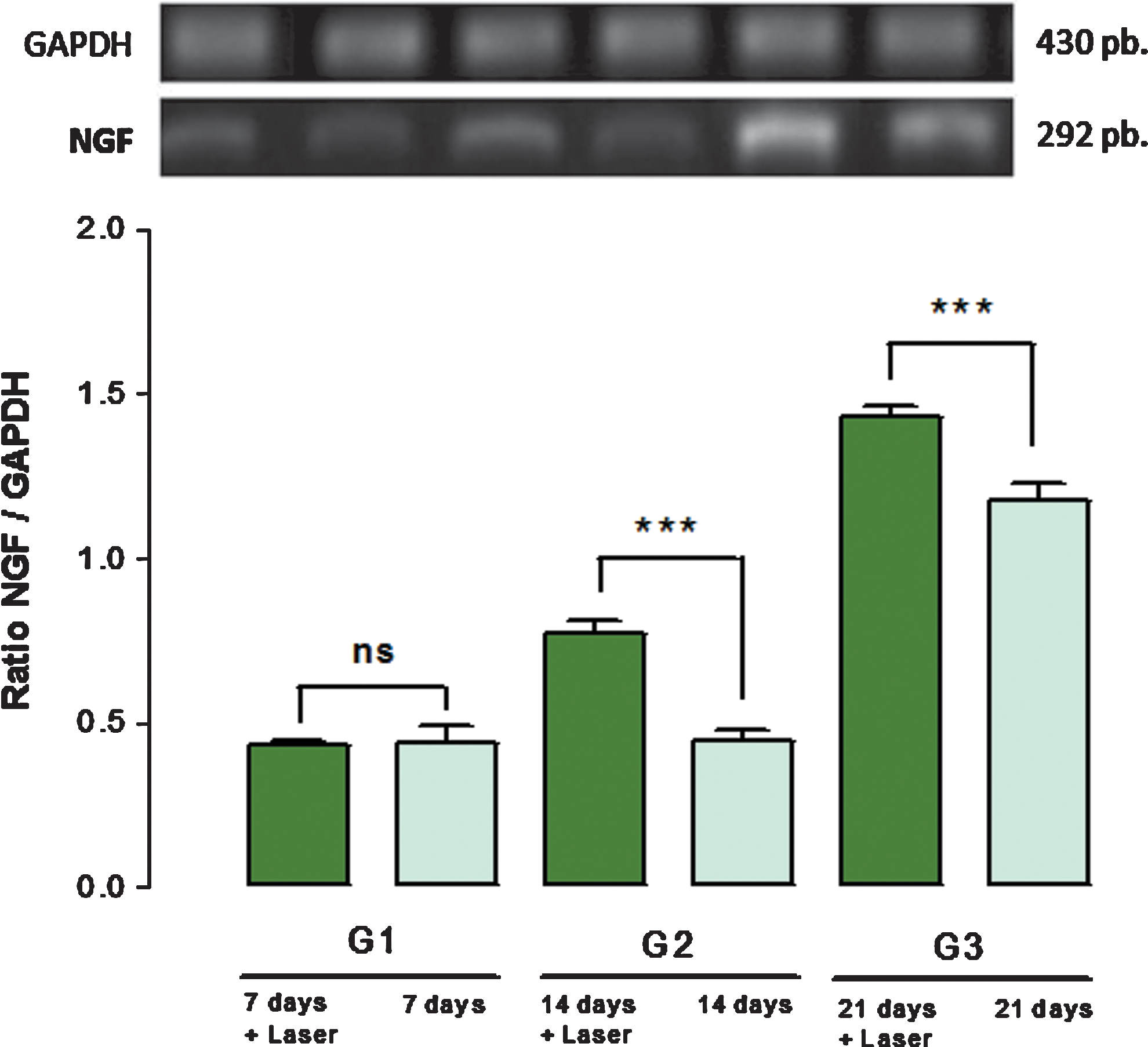
Nerve growth factor (NGF) expression over 21 days. The upper portion illustrates GAPDH and NGF expressions. Below, there are columns corresponding to NGF expression with (irradiated) and without (nonirradiated) low-level laser therapy (LLLT) at days 7, 14, and 21 after nerve crush injury. *** Significant differences, p<0.001.
Comparisons between irradiated and nonirradiated groups revealed that LLLT was also able to promote increases in NGF expression. Although there is no statistically significant difference between irradiated and nonirradiated groups on the 7th day after axonotmesis, on the 14th day LLLT produces its most marked effects, reaching its action peak on the 21st day after sciatic crush injury. Unlike BDNF, which is progressively and constantly expressed, NGF expression seems to happen in jumps, manifesting itself in an intense and tonic way during 2 weeks and then undergoing an abrupt increase in its expression from the 14th day to the 21st day after nerve crush injury (Fig. 3).
It was also observed that the iNOS enzyme's spontaneous behavior was similar to NGF and BDNF expressions. Soon after axonotmesis, an increasing iNOS expression took place, reaching its peak at the 21st day after the nerve crush injury. In addition, it was also observed that LLLT produced statistically significant reductions in iNOS expression at the 21st day after axonotmesis, when compared with respective control group, as can be seen in Fig. 4.

Induced nitric oxide synthase (iNOS) expression over 21 days. The upper portion illustrates GAPDH and iNOS expressions. Below, there are columns corresponding to iNOS expression with (irradiated) and without (nonirradiated) LLLT at days 7, 14, and 21 after nerve crush injury. ***Significant differences, p<0.001.
Discussion
Although progress has been made in regeneration techniques over the last 30 years, functional recovery after peripheral nerves lesions is often incomplete. Several studies have focused on enhancing nerve regeneration using neurotrophic factors, which play an important role in nerve regeneration. Neurotrophic factors produced by the lesion surrounding Schwann cells promote neurite extension after axotomy, suggesting that they may be necessary for regenerating neurons in vivo. 2 –6,22
The iNOS expression reduction, and, mostly, the increased expression of both BDNF and NGF are the findings that helped us to understand, at least in part, the functional recovery produced at an accelerated rate after LLLT in previous experiments conducted in our laboratory, 14,16,17 as well as by other elegant investigations. 15,23 –25 Under the same experimental conditions as described here, HeNe applications produced increasing functional improvement detected by the Sciatic Function Index and by ankle stance angle starting at the 14th day after axonotmesis. 14,16 Similarly, the gastrocnemius electromyography showed both fibrillation potentials and large positive sharp wave reduction starting at the 14th day, as well as polyphasic potentials (nascent potentials) appearing starting at the 21st day of LLLT. 17
A robust set of evidence has shown that LLLT is able to induce regenerative responses in different tissues, including peripheral nervous tissue. 15 However, that information is insufficiently clear about the relationship among LLLT, neurotrophic factors, and inflammatory markers, as the laser action mechanisms are not yet fully elucidated in this context.
It has been shown that LLLT significantly increases the genes' and/or proteins' expression of various growth factors, making the tissue regeneration more effective. 26 In 1993, Anders and colleagues evaluated the HeNe laser effect on the horseradish peroxidase (HRP) retrograde axonal transport of rat facial nerve, and found that LLLT increased nerve regeneration, as there was remarkable retrograde tracer deposit in the cell body of that nerve motor nucleus. 27 Schwartz and colleagues 28 demonstrated that cultured myotubes irradiated with HeNe during 24 h increased NGF expression, possibly because of increased NGF synthesis by muscle cells. Currently it is known that NGF is one of several neurotrophic factors that have the ability to promote survival, differentiation, maintenance, and, when possible, the regeneration of neurons. 6
Although the physiological effects and action mechanisms are completely different from LLLT, femoral nerve electrical stimulation of rats increased the BDNF expression on motor neurons and consequently accelerated axonal regeneration, verified through the distal and proximal portion reconnections of the damaged nerve, 29 showing us the strong relationship between BDNF expression and nerve regeneration after peripheral nerve lesion.
It is noteworthy that, as far as we know, there are no data in the literature establishing categorically the relationship between LLLT and BDNF and NGF neurotrophic factor expressions after peripheral nerve lesions.
Additionally, the lack of response in NT-3 expression is consistent with findings of other researchers who also failed in identifying, either by induction or spontaneously, the NT-3 induced recruitment by peripheral nerve injury. 30 –32
The therapeutic effects of different lasers on biological tissues, such as trophic, regenerative, analgesic, and anti-inflammatory, are very great. 33 As for its anti-inflammatory effects, LLLT seems to be directly related to reduction of nitric oxide (NO) production by activated cells. 34 NO is a potent inflammation mediator synthesized by nitric oxide synthase (NOS) in various parts of the body, including in some neurons and macrophages. 34 The iNOS enzyme is a NOS isoenzyme that is activated in macrophages and endothelial cells, and also in Schwann cells by direct action of pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α) or interleukin-1 beta (IL-1 β), released after inflammatory lesions including nerve injury. 26 Schwann cells are known because their ability in producing a variety of growth factors is important to the maintenance and growth of nerves, and the peripheral nerve lesion such as axonotmesis quickly starts the Wallerian degeneration, activating iNOS to produce NO, and at the same time reducing the neurotrophic factor levels produced by Schwann cells. 35
NO cellular sources in the peripheral nervous system have been extensively described and were therefore not studied further. There is much evidence that monocytes/macrophages active in both peripheral nerves and Schwann cells inflammation are the predominant NO sources after nerve injury. 36 –39
Moreover, the inflammatory stimulus leads to excessive free radical production, reactive oxygen species (ROS) such as hydrogen peroxide, hydroxyl radical, and nitrogens, such as peroxynitrite, which are highly toxic to the tissues, and together they produce oxidative stress. 34,40 In some situations in which high concentrations of these “species” are produced, they can damage vital cellular constituents as well as activating transcription factors that are harmful to nerve regeneration. 13 In this context, we think that HeNe or AsGa lasers can decrease ROS production 12 and protect the Schwann cells to release trophic factors, reducing the recovering time after nerve injury. 35
LLLT can potentially become a therapeutic alternative to anti-inflammatory drugs, as demonstrated through the study of inflammatory process induction by a polysaccharide extracted from Saccharomyces cerevisiae cell wall followed by LLLT application that showed iNOS inhibition, and therefore reduction in NO production, leading to significant reduction in the nerve regeneration, 12 Although during that investigation a laser was used with different physical characteristics than those of the one used in our experiments, we also noticed iNOS expression reduction after using laser, which may explain, at least in part, the inflammatory process reduction detected by other investigators, who observed reduced pro-inflammatory cytokines TNF-α and IL-1β mRNA expression. 27,40,41 Finally, we suspect that iNOS expression reduction could be a result of a direct LLLT effect on the pro-inflammatory cytokines TNF-α and IL-1β expression. 42
Conclusions
The results presented here confirm the LLLT beneficial effects on peripheral nerve regeneration and reveal a possible interaction between HeNe laser and the particularly increased neurotrophic factor expressions. In addition, LLLT was able to inhibit iNOS expression, maybe reducing the inflammatory process mediated by that enzyme. Finally, more studies must be held in order to confirm the hypothesis that pro-inflammatory cytokines are responsible for inducing iNOS expression in this experimental model and to try to establish direct correlations between iNOS and neurotrophic factors after LLLT.
Footnotes
Acknowledgment
The authors thank the Conselho Nacional de Desenvolvimento Cientifico e Tecnológico (CNPq) for the scientific initiation scholarship given to Lessandra Esper Abdala Gomes.
Author Disclosure Statement
No competing financial interests exist.