
Abstract
Introduction
Low-intensity laser irradiation (LILI) has been shown to modulate biological processes such as an increase in ATP synthesis, 1 –4 collagen synthesis, release of growth factors from cells, and cell proliferation. 5 LILI has been widely used to treat medical conditions such as difficult-to-heal wounds, soft tissue injuries, arthritis, and skin traumas, among others. The mechanism of LILI is not well understood and many theories exist. Recent findings suggest that the therapeutic effects of LILI stem in part from intracellular signaling mechanisms activated by the interaction of laser light with the mitochondrial photoacceptor molecule, cytochrome c oxidase. 5,6 It is now generally accepted that the mitochondria are the initial site of action in mammalian cells, and that this is because of a response from cytochrome c oxidase. 7 Eells et al. 5 showed that near infrared light-emitting diodes (NIR-LED) increased the production of cytochrome c oxidase in cultured primary neurons. Houreld et al. 4 showed that visible red laser light (660 nm) influenced mitochondrial enzyme activity; there was an increase in cytochrome c oxidase activity. Silveira et al. 1 showed that LILI at 904 nm increased the enzymatic activity of complexes I, II, III, and IV. An increase in ETC activity leads to an increase in ATP, which can translate into increased cell activities such as cell proliferation and migration, both of which are important aspects of wound healing.
LILI has been shown to significantly increase gene expression of various growth factors in numerous cell types, including transforming growth factor beta (TGF-β), 8 –11 vascular endothelial growth factor (VEGF), 9 and insulin-like growth factor (IGF). 9 Collagens and other proteins involved in the extracellular matrix (ECM) have also been found to be influenced. These include procollagen, 8 collagen, 9,11,12 tissue inhibitors of metalloproteinases (TIMPs), 8,13,14 and matrix metalloproteinases (MMP). 8,11,13,14 Other factors such as bone morphogenic protein (BMP), 10 osteocalcin, 10 hypoxia-inducible factor (HIF), 10 heat-shock protein 70 (Hsp70), 8 cytokines, 8,11,14 –16 and receptor activator of NF-KB (RANK) 17 have also been regulated by LILI.
This study aimed to determine the effect of LILI in the visible red spectrum (660 nm) on genes involved in the ETC and oxidative phosphorylation.
Methods
Cell culture
Human skin fibroblast cells were isolated from excised dermal tissue from consenting adult donors undergoing abdominoplasty (Faculty of Health Sciences, University of Johannesburg, Academic Ethics Clearance Number 01/06). Cells were grown as previously described.
4
For experiments, 6×105 cells were seeded into 3.4 cm diameter tissue culture plates. A wound model was based on the central scratch method, whereby a confluent monolayer of cells is scraped with a sterile 1 mL pipette, thereby creating a cell free zone.
18
A diabetic cell model was achieved by continuously growing cells in complete medium containing additional
Laser irradiation
Fibroblast cells (6×105) were irradiated from above, via fiberoptics, in the dark in 3.4 cm diameter tissue culture dishes, with the lid off, with a 660 nm continuous wave diode laser at a fluence of 5 J/cm2. A summary of the laser parameters can be seen in Table 1. Normal, non-irradiated control cells (0 J/cm2) were kept on the bench in a dark box. Prior to each irradiation, power output was measured using a power meter (Fieldmate, 0398D05) and used to calculate the duration of exposure. Post-irradiation, cells were incubated at 37°C for 30 min.
RNA isolation and cDNA synthesis
The RNeasy Mini Kit (Whitehead Scientific, Johannesburg, South Africa, Qiagen, 74104) with the QIAshredder homogenizers (Whitehead Scientific, Johannesburg, South Africa, Qiagen, 79654) was used on the QIAcube (Qiagen) for the isolation and purification of total RNA. Cells were trypsinized, washed in phosphate buffered saline (PBS), and mixed in 600 μL of a highly denaturing guanidine-thiocyanate-containing buffer (Buffer RLT) which immediately inactivates RNase. Isolated RNA was quantified using the Quant-iT™ RNA Assay kit (AB Applied Biosystems, Johannesburg, South Africa, Invitrogen, Q32852) on the Qubit™ fluorometer (Invitrogen). Purity was determined by calculating the ratio between the absorbance values at 260 and 280 nm.
cDNA was synthesized using the QuantiTect Reverse Transcription Kit (Whitehead Scientific, Johannesburg, South Africa, Qiagen, 205311) according to the protocol instructions. Briefly 1 μg RNA was treated with DNase to ensure elimination of genomic DNA (gDNA). This mixture was incubated at 42°C for 5 min, and then chilled on ice. RT master mix was then added and incubated at 42°C for 15 min followed by 3 min at 95°C. cDNA was stored at −20°C until used for gene expression profiling.
Gene expression profiling
The SABiosciences human mitochondrial energy metabolism RT2 Profiler PCR Array System (Whitehead Scientific, Johannesburg, South Africa, SABiosciences, PAHS-008A) was used to evaluate mitochondrial gene expression (Table 2). This PCR array profiled the expression of 84 key genes involved in mitochondrial respiration, including genes encoding components of the ETC (complexes I–IV) and oxidative phosphorylation complexes. Real-time quantitative PCR (qPCR) detection was performed according to the manufacturer's specifications. Briefly, cDNA was thawed on ice and made up to a final volume of 111 μL. cDNA was added to the RT2 qPCR master mix (containing ROX™ as the reference dye and SYBR green) and 25 μL was added to each well of the 96-well PCR array and all air bubbles removed. Real-time qPCR was performed on the Stratagene MX3000p (Anatech, Olivedale, South Africa) using the following cycles: 1 cycle at 95°C for 10 min; 40 cycles at 95°C for 15 sec, and 1 min at 60°C. A dissociation (melting) curve was also performed to ensure the amplification of a single product in each well. This was done at 95°C for 1 min and at 65°C for 2 min; readings were taken at 65–95°C at 2°C per minute intervals. A single peak at temperatures >80°C had to be reached.
Wells A1 through G12 contain genes involved in the electron transport chain (complex I to IV) and oxidative phosphorylation (adenosine triphosphate [ATP] synthase). Wells H1 through H5 contain housekeeping genes to normalize polymerase chain reaction (PCR) array data. Well H6 contains the human genomic DNA control (HGDC). Wells H7 through H9 contain replicate reverse transcription controls (RTC), and wells H10 through H12 contain replicate positive PCR controls (PPC).
The plate included positive PCR controls (PPC), gDNA controls, and five housekeeping/reference genes. All threshold cycle (Ct) values for PPCs had to be 20±2 and for gDNA>35; all wells with Ct values>35 were considered negative. Ct values for all the wells were exported to an Excel-based Data Analysis Template (available from the SABiosciences website) with the appropriate pathway-focused genes (PAHS-008A). The software performed all the necessary calculations (ΔΔCt method) and interpretation of the control wells. The fold-change (2^(-ΔΔCt)) was also calculated and if >1, results were reported as fold upregulation, and if <1 results were reported as fold downregulation.
Statistical analysis
Experiments were repeated three times (n=3). The student t-test was performed by the SABiosciences excel-based Data Analysis Template and reported as significant if p<0.05. Results are shown as a 3D profile.
Results
RT-PCR results showed that three genes, cytochrome c oxidase subunit VIb polypeptide 2 (COX6B2), cytochrome c oxidase subunit VIc (COX6C), and pyrophosphatase (inorganic) 1 (PPA1) were significantly upregulated (p<0.01, p<0.01, and p<0.05, respectively) in diabetic wounded cells (Fig. 1a and b). COX6B2 (Gene ID: 125965) and COX6C (Gene ID 1345) belong to the complex IV family, whereas PPA1 (Gene ID 5464) is involved in oxidative phosphorylation. Four genes, COX6C, ATP synthase, H+transporting, mitochondrial Fo complex, subunit B1 (ATP5F1), NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, 11 (NDUFA11), and NADH dehydrogenase (ubiquinone) Fe-S protein 7 (NDUFS7) were significantly upregulated (p<0.001, p<0.01, p<0.05, and p<0.05 respectively) in wounded fibroblast cells (Fig. 1c and d). ATP5F1 (Gene ID: 515) is involved in oxidative phosphorylation, and NDUFA11 (Gene ID: 126328) and NDUFS7 (Gene ID: 374291) are part of complex I. Two genes, ATPase, H+/K+exchanging, beta polypeptide (ATP4B) and ATP synthase, H+transporting, mitochondrial Fo complex, subunit C2 (subunit 9) (ATP5G2) were significantly upregulated (p<0.05) in ischemic cells post-irradiation (Fig. 1e and f). ATP4B (Gene ID: 496) and ATP5G2 (Gene ID: 512) both encode subunits of ATP synthase. A summary of the significantly upregulated genes is displayed in Table 3.
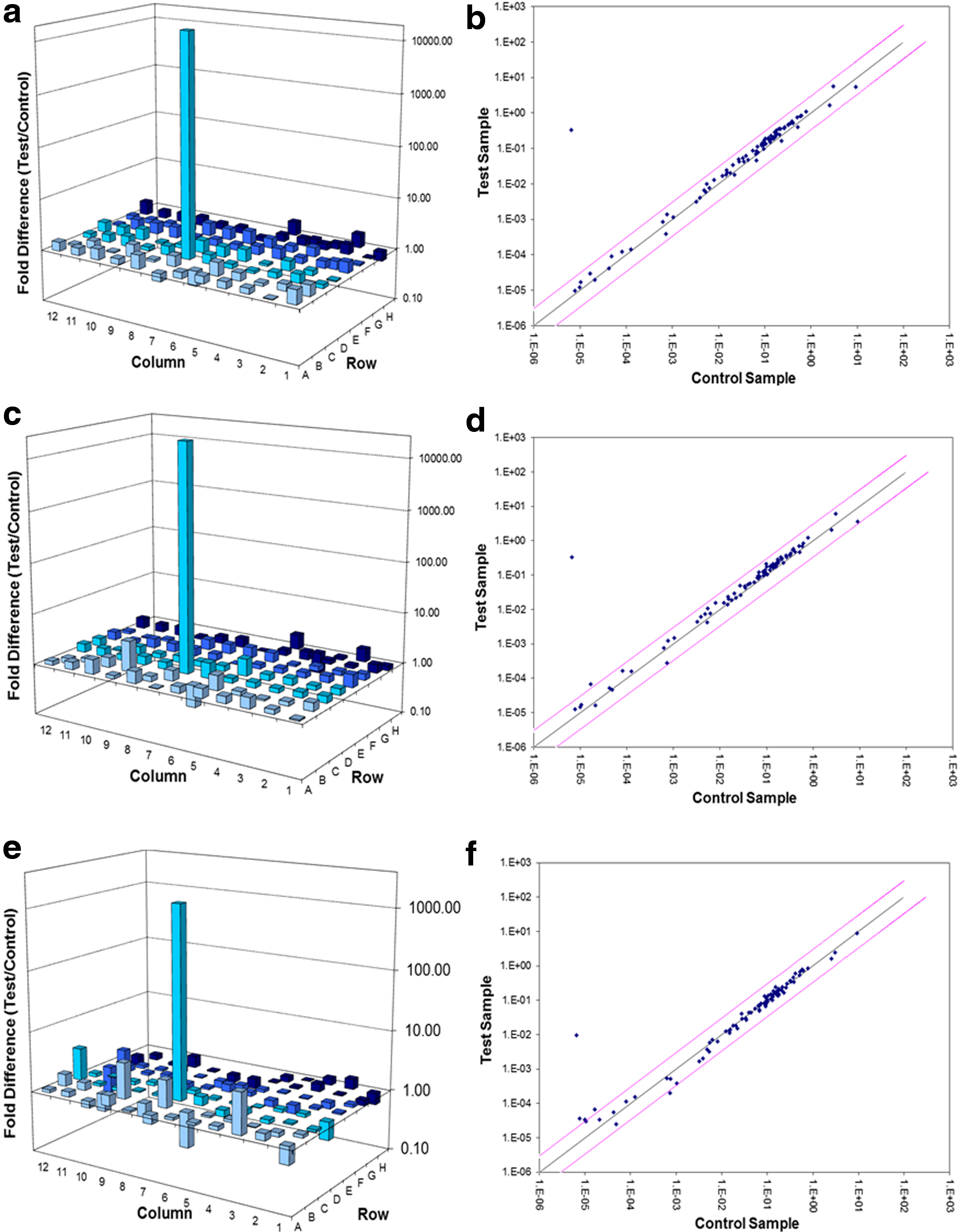
Real-time reverse transcription quantitative polymerase chain reaction (RT-qPCR) was performed on irradiated (660 nm, 5 J/cm2) wounded (
LILI, low-intensity laser irradiation; NADH, nicotinamide adenine dinucleotide; ATP, adenosine triphosphate.
Discussion
Mitochondria are double membranous organelles found in varying quantities, depending upon cell type, in the cytoplasmic fraction of cells. They are involved in a variety of metabolic pathways, such as apoptosis, cellular differentiation, cellular division and growth, and cellular signalling. However, the most important pathway they are involved with is the production of energy in the form of ATP through the ETC, via a process known as oxidative phosphorylation. Four complexes (I, II, III, and IV) form part of the ETC, and some authors consider ATP synthase to be a fifth complex. In complex I, NADH is oxidized to NAD and the free electrons are passed onto ubiquinone (coenzyme Q). Succinate is oxidized to fumerate by complex II with the passage of electrons to ubiquinone which is itself reduced to ubiquinol. Electrons are then passed onto complex III, and subsequently cytochrome c and complex IV where molecular oxygen is reduced to water. ATP synthase utilizes the energy released when protons are pumped back through the enzyme, across the electrochemical gradient back into the matrix, to phosphorylate ADP to ATP.
LILI has been shown to modulate the activity of these ETC enzymes. Houreld et al., 4 showed that 660 nm increased the activity of cytochrome c oxidase (complex IV). At a wavelength of 904 nm an increase in succinate: ubiquinone oxidoreductase (complex II) and cytochrome c oxidase activity was seen, 1,23 whereas an increase in NADH: ubiquinone oxidoreductase (complex I), cytochrome bc1 (complex III), and cytochrome c oxidase activity was seen at 660 nm. 24 Karu 25 has put forward that cytochrome c oxidase is the photoacceptor and photosignal transducer for laser light in the visible (400–760 nm) and infrared (IR)-A region (760–1440 nm). It has also been shown that LILI with a He-Ne laser increases mitochondrial membrane potential, cyclic adenosine monophosphate (cAMP) and ATP via enhanced cytochrome c oxidase activity. 26 This study evaluated the effect of laser light at a wavelength of 660 nm and a fluence of 5 J/cm2 on 84 genes involved in the ETC and oxidative phosphorylation in wounded, diabetic wounded, and ischemic fibroblast cell models.
COX6C, ATP5F1, NDUFA11, and NDUFS7 were all upregulated in irradiated wounded cells. NDUFA11 and NDUFS7 both encode for a subunit in complex I, which has >40 subunits. COX6C encodes for a subunit in complex IV and is involved in the regulation and assembly of the complex. Cytochrome c oxidase is a heteromeric complex consisting of three catalytic subunits, encoded for by mitochondrial DNA, and multiple structural subunits, all coded for by nuclear DNA. 27 ATP5F1 encodes for a subunit of ATP synthase, more specifically the b subunit of the proton channel. ATP synthase is made up of two multi-subunit complexes, namely the membrane-spanning component (Fo), which comprises the proton channel and the soluble catalytic core (F1). The catalytic fraction is made up of five subunits (alpha, beta, gamma, delta, and epsilon) assembled with a stoichiometry of three alpha, three beta, and a single representative of the other three subunits (gamma, delta, and epsilon). The proton channel has nine subunits (a, b, c, d, e, f, g, F6, and 8). 27
COX6C, COX6B2, and PPA1 were all upregulated in irradiated diabetic wounded cells. COX6C and COX6B2 both encode for the structural subunits in complex IV. PPA1 is a member of the inorganic pyrophosphatase (PPase) family and catalyses the hydrolysis of pyrophosphate to inorganic phosphate. 27 In ATP synthase, an inorganic phosphate group is added to ADP to produce ATP; therefore, the protein coded for by PPA1 plays an important role in the oxidative phosphorylation of ADP.
ATP4B and ATP5G2 were both upregulated in irradiated ischemic cells. ATP4B encodes for a protein (beta subunit) belonging to a family of P-type cation-transporting ATP synthase. ATP synthase is a heterodimer consisting of a catalytic alpha subunit and a smaller glycosylated beta subunit. ATP synthase is a proton pump catalysing the hydrolysis of ATP coupled with the exchange of H+ and K+ ions across the plasma membrane. 27 ATP5G2 codes for a precursor of subunit c of the proton channel in ATP synthase. Subunit c is encoded for by three separate genes that specify precursors with different important sequences but identical mature proteins. 27
Raza et al. 28 found that the activities of complexes III and IV were significantly decreased in streptozotocin (STZ)-induced diabetic rats, that complexes I and II were moderately decreased, and that mitochondrial respiratory complexes may play a role in reactive oxygen species homeostasis and oxidative stress. This study showed that there was an increase in transcription of genes involved in complex IV and ATP synthase post-irradiation of diabetic wounded cells. In a previous study, it was also shown that LILI was able to significantly increase the activity of cytochrome c oxidase and ATP in irradiated diabetic mitochondria. 4 McDaniel et al., 29 showed that LILI influenced the expression patterns of genes involved in the extracellular matrix and adhesion molecules. It is evident that LILI alters the expression of a wide variety of genes involved in a range of metabolic and cellular pathways, and more of this type of research is needed to fully understand the complete molecular effect. What is interesting about the results in this study is that LILI upregulated different genes in the different cell models. NDUFA11 and NDUFS7, which form part of complex I, were found to be upregulated only in irradiated wounded fibroblast cells. Complex I has been found to be decreased in diabetic situations, which may explain why enzymes relating to this complex were not upregulated in hypoxic and diabetic wounded irradiated fibroblast cells. Mitochondrial dysfunction and DNA mutations have been detected in diabetic patients. 30 Silveira et al., 1 showed that LILI at 904 nm increased the enzymatic activity of complex I in wounded rat muscle cells.
LILI has been shown to stimulate wound healing in vivo; however, the exact underlying effects are poorly understood. This study has shown that irradiation in the red spectrum of light stimulates the expression of genes involved in the ETC, and hence ATP production. An increase in ATP will ultimately lead to an increase in DNA and RNA synthesis, and, therefore, increased protein synthesis. There is also a stimulation of other biological molecules and signaling cascades needed for cellular regeneration and wound repair. 31 Despite the fact that this study has been done in vitro, it is possible to reason that similar results would be found in the clinical setting. However, this would have to be confirmed. This study provides a link between what is seen in vitro with what is found in vivo. In a study conducted by Lapchak and Taboada, 32 it was shown that there is a direct relationship between cortical ATP levels in embolized rabbits and fluence. Oron and colleagues 33 showed that LILI applied to mice following traumatic brain injury lead to an increase in ATP, angiogenesis, antioxidants, and decreased apoptosis. It has been shown that reduced axonal transport of mitochondria is a contributory factor in Parkinson's disease, and that this reduced transport is directly related to ATP deficiency. LILI has restored axonal transport in neuronal cells in patients with Parkinson's disease. 34 The metabolite of methanol, formic acid, inhibits cytochrome c oxidase and damages the retina and optic nerves. Eells et al. 35 demonstrated that LILI is able to significantly attenuate the effects of methanol intoxication in a rodent model.
Conclusions
In conclusion, this study showed that laser irradiation at a wavelength of 660 nm and a fluence of 5 J/cm2 induced the transcription of genes encoding for subunits involved in the ETC complexes I, IV, and V (ATP synthase) in human skin fibroblast cell models. There is an upregulation of genes involved in energy metabolism and oxidative phosphorylation, thus stimulating the increase in ATP production, which drives other metabolic processes involved in wound healing, all leading to normalization of cellular functions. Thus, LILI at 660 nm not only increases the activity of these enzymes, but also increases transcription of the genes that code for these proteins. Further investigations into the biochemical events produced by LILI could further reveal the mechanisms behind laser irradiation. However, a strong connection between the mitochondria and signalling cascades has been made.
Footnotes
Acknowledgments
This study was funded by the University of Johannesburg, and the African Laser Centre. Lasers were supplied by the National Laser Centre, South Africa.
Author Disclosure Statement
No competing financial interests exist.