
Abstract
Introduction
Materials and Methods
The detection of intra- and extracellular PpIX
Rat glioma cell line C6 (obtained from American Type Culture Collection, Rockville, MD), human glioma cell lines U87 delta (obtained from Department of Neurosurgery, Saitama Medical College, Saitama, Japan) and U251 (obtained from American Type Culture Collection, Rockville, MD), and human malignant meningioma cell line IOMM-Lee (obtained from Department of Neurosurgery, University of California, San Francisco, CA) were cultured in a monolayer in Dulbecco's Modified Eagle's Medium (DMEM; Invitrogen, Tokyo, Japan) buffered with 25 mM bicarbonate, and supplemented with 10% heat-inactivated fetal bovine serum (Invitrogen, Grand Island, NY) in a fully humidified incubator at 37°C with 5% CO2. A total of 103 cells were seeded into 24 well plates (FALCON, Becton Dickinson Labware, U.S.A.) and the medium was renewed 24 h before the experiment.
5-ALA (COSMO BIO, Japan) was dissolved in phosphate-buffered saline (PBS) and the pH was adjusted to 7.4 with NaOH. 5-ALA was added to the medium so that the final solution contained 0, 0.005, 0.01, 0.05, 0.1, 0.5, and 1.0 mM. 10 Then all cell lines were cultured for 3, 6, 9, 12, and 24 h. It should be noted that prior to this study, we examined the toxicity of 5-ALA for the cell lines, and we set the concentration range at low toxicity.
After the cell culture, the medium was separated from the cells, and PpIX concentrations in medium and cells were individually, that is, extracellular PpIX and intracellular PpIX, examined by means of photometrical assay using Fluoroskan Ascent (Thermo Labsystems Oy, Vantaa, Finland). Fluorescence intensity at 620 nm (excitation at 390 nm) was measured on a Fluoroskan Ascent FL multiwell fluorescence plate reader and converted to the concentration of PpIX. We measured the calibration curve of concentration of PpIX and fluorescence intensity, and converted fluorescence intensity to concentration of PpIX.
The effect of BBB disruption on PpIX fluorescence in the rat brain after 5-ALA administration
Six rats received an intraperitoneal injection of 5-ALA 100 mg/kg 5 min after intra-right carotid infusion of 20% mannitol. 11 –13 As a reference group, three rats underwent intraperitoneal infusion of 5-ALA 100 mg/kg without mannitol administration.
All rats were euthanized, and their brains were removed 4 h after injection of 5-ALA. The rat brain was radiated using excitation laser light at 405 nm, and the fluorescence was observed via a low-cut filter at 500 nm. The laser we used was a diode laser emitting 405 nm of visible light (Ball Semiconductor, Frisco, TX). Its intensity is 300 mW at the tip, and 10 mW at distance of 20 cm from the tip.
The effect of direct infusion of 5-ALA into brain on PpIX fluorescence in rat brain
Seven rats underwent stereotactic cannula placement into the right basal ganglia followed by infusion at a constant rate of 20 μL/h in a total volume of 20 μL using convection-enhanced delivery (CED). 14,15 Three rats were given 180 mM of 5-ALA with PBS solution, three rats were given the same amount of PBS as a negative control, and one rat was given PpIX solution as a positive control.
All rats were euthanized and their brains were removed 4 h after infusion. Fluorescence of rat brains was observed by the same method.
The effect of BBB disruption on PpIX fluorescence at rat brain after PpIX administration
Three rats received an intraperitoneal injection of PpIX 100 mg/kg 5 min after intra-right carotid infusion of 20% mannitol. 11 –13 All rats were euthanized and their brains were removed 4 h after injection. Fluorescence of rat brain was observed by the same method.
All procedures were subjected to the Osaka Medical College Regulations on Animal Experimentation.
Results
The intracellular and extracellular concentration of PpIX
Both intra- and extracellular PpIX concentrations were increased in a dose-dependent manner, up to 0.5 mM of 5-ALA in the C6 and U87 delta cell lines (Fig. 1). The PpIX concentration of extracellular space is 8–12 times higher than that of intracellular space. On the other hand, the PpIX production is quite low in both intra- and extracellular spaces in the U251 and IOMM-Lee cell lines.
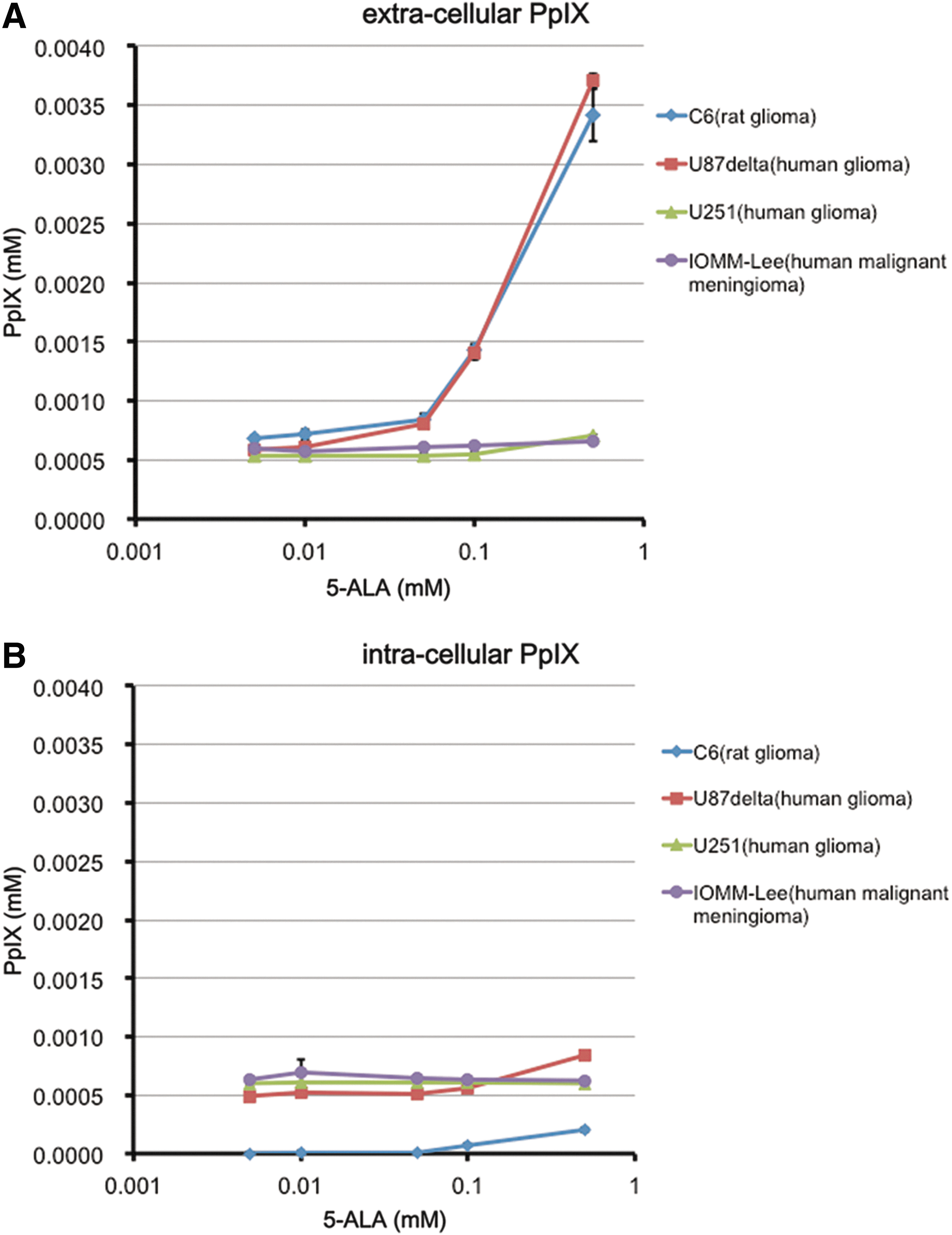
In vitro, we exposed various cultured glioma and meningioma cell lines to different conditions of 5-aminolevulinic acid (5-ALA). After this, the degree of fluorescence of the culture medium and cells (i.e., extracellular Protoporphyrin IX [PpIX] and intracellular PpIX), was each quantitatively measured and analyzed by means of photometrical assay. The extracellular
The effect of BBB disruption on PpIX production after 5-ALA administration in the rat brain
In the three BBB-disrupted rat brains, diffuse PpIX fluorescence could be detected, especially in the white matter, by systemic administration of 5-ALA (Fig. 2A). The disruption of BBB was confirmed by methylene blue distribution (Fig. 2B). No methylene blue or PpIX fluorescence was observed in the reference group without intracarotid infusion of mannitol. PpIX fluorescent areas were larger than the methylene blue area.
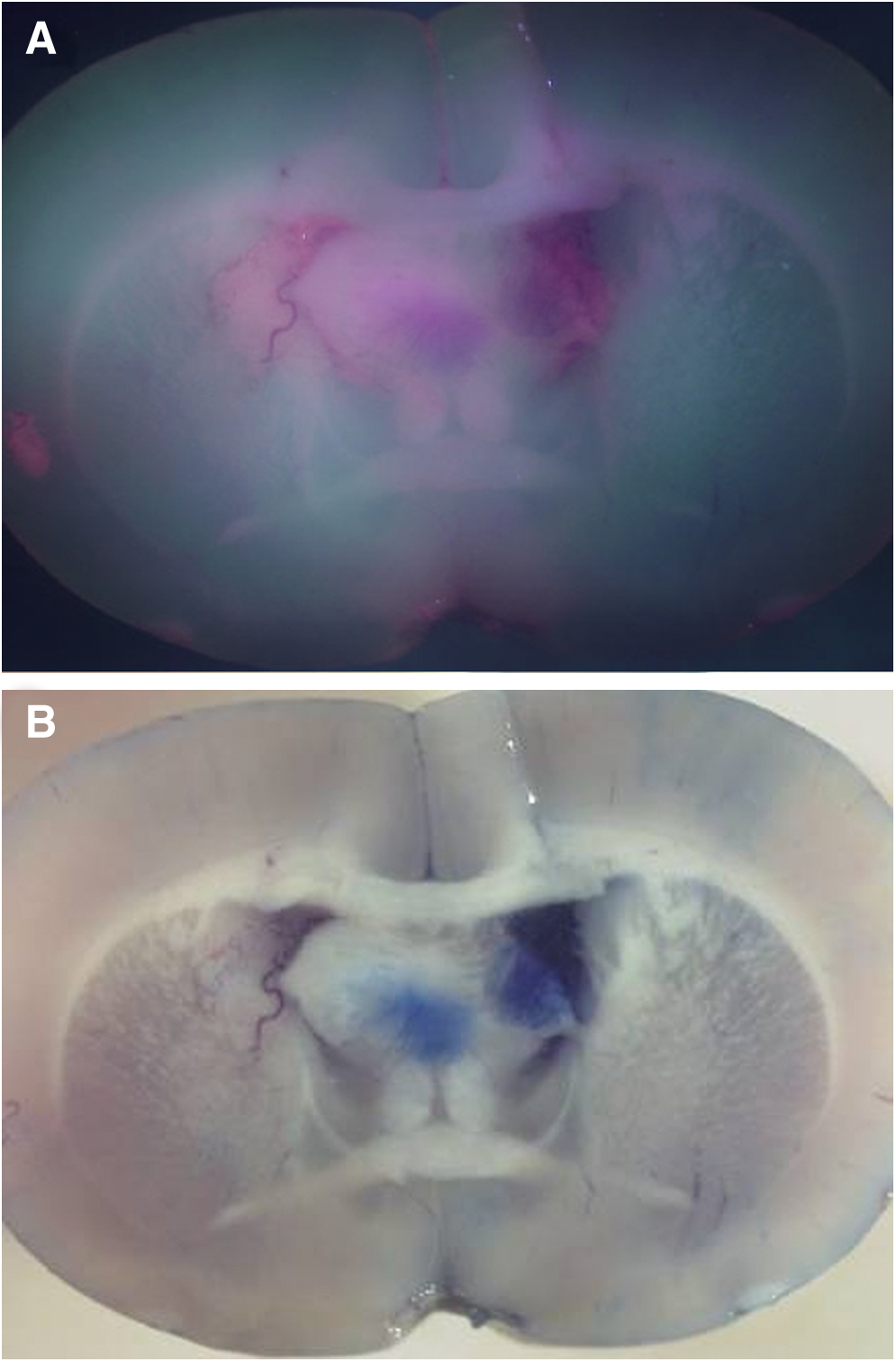
Six rats received an intraperitoneal injection of 5-aminolevulinic acid (5-ALA) with or without an intracarotid infusion of mannitol. The rat brain was radiated using excitation laser light at 405 nm and the fluorescence was observed via a low-cut filter at 500 nm. In the three blood–brain barrier (BBB)-disrupted rat brains, diffuse Protoporphyrin IX (PpIX) fluorescence could be detected, especially in the white matter, by systemic administration of 5-ALA (
PpIX fluorescence at rat brain after direct infusion of 5-ALA into the brain
All the animal brains into which 5-ALA was directly infused by CED (Fig. 3A) showed strong PpIX fluorescence (Fig. 3B). PpIX fluorescence spread to the contralateral hemisphere via the corpus callosum. Direct infusion of PpIX (Fig. 3C,D) also caused the PpIX fluorescence at the injected area. On the other hand, all animal brains in which only PBS was directly infused by CED showed no PpIX fluorescence, and rat brains with BBB disruption by intracarotid infusion of mannitol showed no PpIX fluorescence after systemic intraperitoneal administration (Fig. 4A,B).
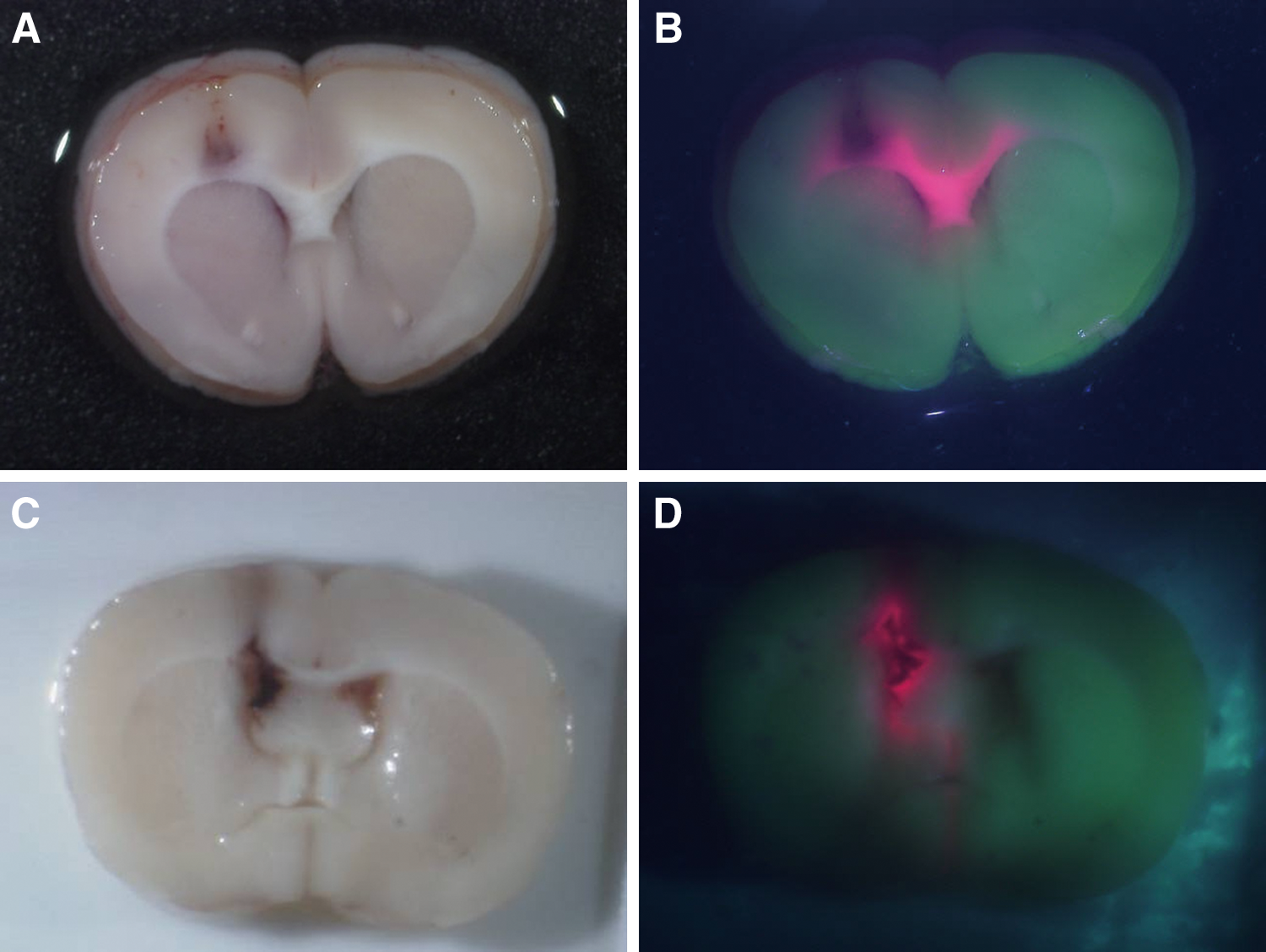
Seven rats underwent stereotactic cannula placement into the right basal ganglia followed by infusion using convection-enhanced delivery (CED). Three rats were given 5-aminolevulinic acid (5-ALA) phosphate-buffered saline (PBS) solution, three rats were given the same amount of PBS as a negative control, and one rat was given Protoporphyrin IX (PpIX) solution as a positive control. All rats were euthanized, and their brains were removed after infusion. Fluorescence of rat brains was observed. All the animal brains into which 5-ALA was directly infused by CED
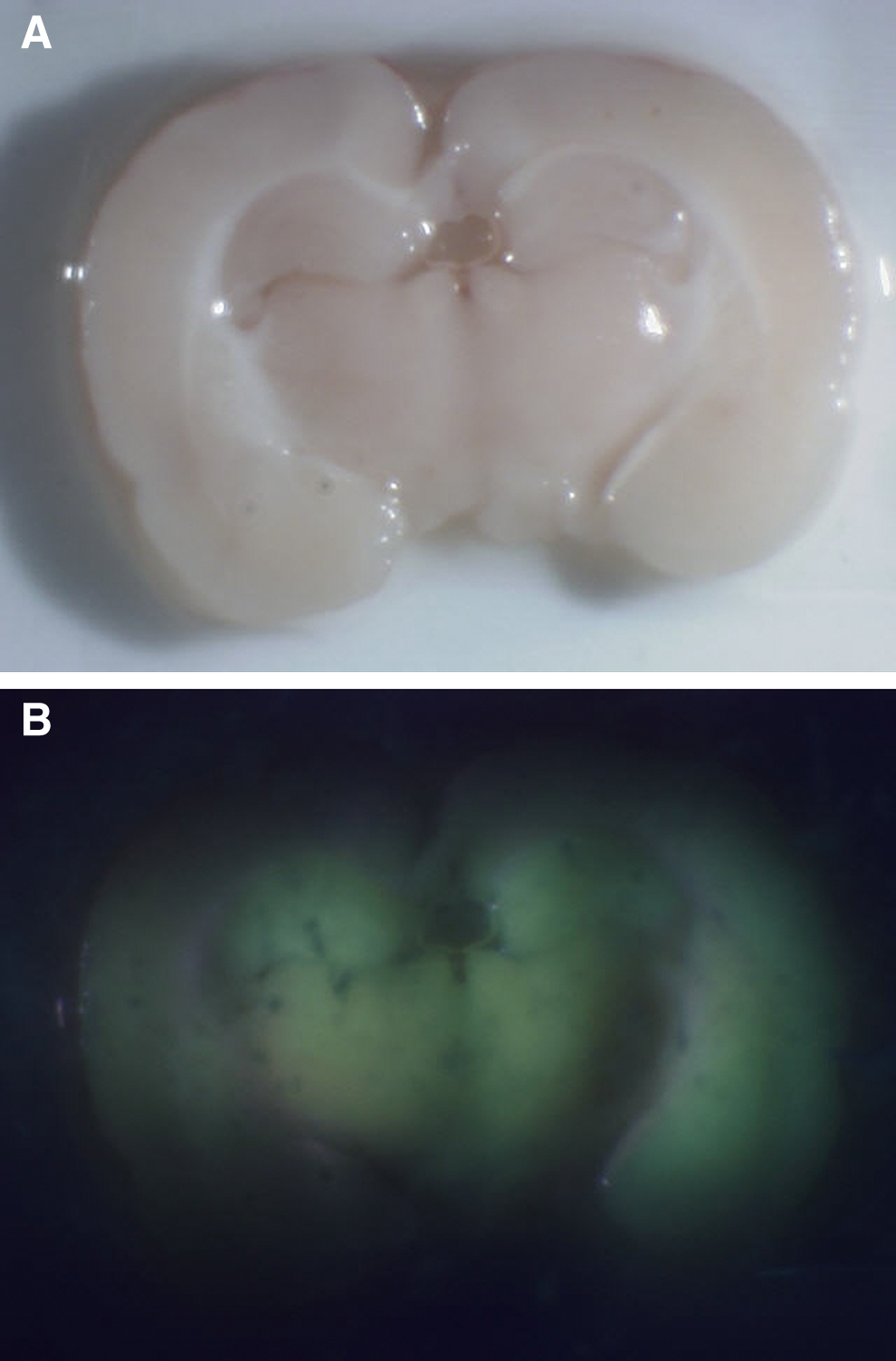
Three rats received an intraperitoneal injection of Protoporphyrin IX (PpIX) after intra-right carotid infusion of mannitol. All rats were euthanized and their brains were removed after injection. Fluorescence of rat brain was observed. Rat brains with blood–brain barrier (BBB) disruption showed no PpIX fluorescence after intraperitoneal administration.
Discussion
It has been established that fluorescence-guided resection using 5-ALA is useful in glioma surgery, because of its high tumor specificity and safety. 1,2,9,16 And for fluorescent diagnosis and photodynamic therapy, the pharmacokinetics and localization of 5-ALA-induced PpIX as a photosensitizer in cells are important. 17 Wu et al. described that PpIX is produced initially in the mitochondria, and then diffuses to the cytosol. 18 Stummer et al. explained that in brain edema, fluorescent porphyrins are first synthesized within the tumor, and then transferred to albumin and lipoproteins in edema, to be then carried into adjoining tissue. 8 We previously reported that not only glioma, but also non-tumor tissue, showed vague and, sometimes, strong fluorescence of PpIX with the administration of 5-ALA. For example, the perinecrotic area in radiation necrosis or neurodegenerative disease shows fluorescence after administration of 5-ALA. 9
As mentioned, the origin of non-neoplastic PpIX is unknown. To elucidate the origin, we formulated three hypotheses and examined them by in vitro and in vivo experiments. Based on the results shown in Figure 1, it is considered that PpIX, produced by tumor cells, moves to the extracellular fluid. However, it is uncertain whether or not, depending upon the cell lines, there are differences in the intracellular and extracellular concentration of PpIX in the ability to convert to PpIX and to transport to extracellular space, because we have not verified them. The involvement of a certain transporter has been suspected. One transporter, ATP-binding cassette (ABC) transporter ABCG2, plays a role in the metabolism of heme. 19,20 Ishikawa et al. showed that ABCG2 transports porphyrins across the plasma membrane to maintain intracellular porphyrin homeostasis. 21 Wakabayashi et al. found that ABCG2 is responsible for the cellular homeostasis of porphyrins and their related compounds. 20 Robey et al. showed that ABCG2-overexpressing cells also displayed decreased intracellular fluorescence of protoporphyrin IX generated by exogenous application of 5-ALA. 22 And, Ishikawa et al. found that ABCG2 plays a key role in regulating the cellular accumulation of porphyrin derivatives in cancer cells, and thereby affects the efficacy of photodynamic therapy and photodynamic diagnosis. 21 Thus, the result shown in Fig. 2, in which PpIX production was detected after intraperitoneal injection of 5-ALA, in rat brains with BBB disruption, indicates the possibility that 5-ALA leaks from blood vessels at the brain tumor and is converted to PpIX by glial cells in the surrounding normal brain. And the result shown in Fig. 3, in which the rat brain after direct infusion of 5-ALA into the brain by CED shows PpIX fluorescence, identically indicates that normal glial cells produce PpIX by exposure to 5-ALA. On the other hand, the result shown in Fig. 4, in which no PpIX fluorescence was seen in the BBB-disrupted rat brain after intracarotid infusion of mannitol and systemic administration of PpIX, indicates the unlikelihood that PpIX produced at other organs leaks from blood vessels at the brain tumor.
In summary, from our study, for the mechanisms for PpIX fluorescence at the brain surrounding tumors after 5-ALA administration, we consider the following. First, the PpIX produced by the main tumor mass is moved to the extracellular fluid; then PpIX reaches to the surrounding brain via the bulk flow of edema fluid. Second, the 5-ALA leaks from the main tumor, where the BBB is disrupted; then 5-ALA reaches the surrounding brain and is converted to PpIX in the glial cells or infiltrating tumor cells. This mechanism explains how 5-ALA-induced PpIX fluorescence can be observed in radiation necrosis and neurodegenerative disease, without tumor cells. We found the hypothesis that PpIX produced at other organs would move into the blood, leak at the main mass, and reach the surrounding brain to be less probable. However, in this study, the following is not elucidated: how each hypothesis affected the origin of PpIX of edematous brain tissues, which contains cells other than tumor cells, and how 5-ALA is converted to PpIX. And, our study is about ALA-induced PpIX fluorescence of the surrounding brain tumor. In other words, we studied the fluorescence in cells other than brain tumor cells, such as normal brain cells, and, therefore, we have not mentioned the brain tumor model.
Conclusions
We sometimes experience PpIX fluorescence in brain tissues near tumors after 5-ALA administration. It is thought that the PpIX produced by the main tumor mass is moved to the extracellular fluid, and reaches the surrounding brain via bulk flow of edema fluid, and/or that 5-ALA leaked from the main tumor where the BBB is disrupted reaches the surrounding brain and converts to PpIX in the glial cells or inflammation cells. We need to pay closer attention to tumor-specific characteristics of PpIX fluorescence.
Footnotes
Acknowledgments
This work was supported by Grants-in-Aid for Scientific Research (C) (23592147) from the Japanese Ministry of Education, Culture, Sports, Science, and Technology (MEXT) to Dr. Kuroiwa. All procedures were subjected to the Osaka Medical College Regulations on Animal Experimentation. Drs. Masubuchi and Fujishiro performed in vitro/in vivo studies; Drs. Kawabata and Nonoguchi designed experiments and interpreted data; Drs. Kajimoto and Kuroiwa supervised all in vitro /in vivo studies, and reviewed the manuscript; and Dr. Miyatake reviewed the manuscript. All authors have read and approved the final manuscript.
Author Disclosure Statement
No competing financial interests exist.