
Abstract
Introduction
Muscle contraction and relaxation require the action of creatine kinase (CK). Phosphocreatine, formed by the reaction of this enzyme, constitutes a reservoir of high energy phosphate that is available for quick resynthesis of adenosine triphosphate (ATP). This high concentration of ATP is then accessible for muscle contraction. In cases of muscle denervation, the level of CK, as well as muscle weight, decreases. 6 In normal intact muscle, acetylcholine receptors (AChR), which play a key role in neuromuscular transmission, are concentrated at the neuromuscular junction. A nerve impulse triggers the release of acetylcholine, producing a much larger end plate potential, which excites the muscle membrane and leads to muscle contraction. The amount of AChR in neuromuscular transmissions appears to increase, and covers the entire extrajunctional area following muscle denervation. In the denervated muscle, the amount of AChR increases prior to muscle degeneration. 7 Much of the literature on the effects of long-term denervation of mammalian skeletal muscle has focused on experimental studies of total sciatic section in rats. 8 –10
In the last decade, interest in the therapeutic effect of laser phototherapy on muscle has risen sharply. In an earlier study, the author used He-Ne laser 11 for the treatment of lesioned muscles, and found that laser could increase mitochondrial activity in muscular fibers, activate fibroblasts and macrophages, and stimulate angiogenesis. Schwartz et al. 12 found that after irradiation of muscle cell cultures (632 nm, 3 J/cm2, 20 mW), there was a rise in the levels of the neurotropic nerve growth factor (NGF) secreted by skeletal muscles, which influences the survival and regeneration of sympathetic neurons in the peripheral nervous system. Other neurotropic growth factors have been reported to be biostimulated by laser therapy, such as growth associated protein (GAP)-43 13 and the fibroblast growth factor. 14
Studies have reported a transient elevation of intracellular calcium in skeletal muscle myotubes immediately after irradiation. This calcium elevation probably triggers cytokine release from the myotubes after irradiation. 15 In addition, it was demonstrated that infrared light protects cardiomyocytes from hypoxic damage by an upregulation in the expression of inducible nitric oxide synthase (iNOS), 16 and that vascular endothelial growth factor was associated with cardioprotection and enhanced angiogenesis. 17 Although these results were shown on cardiac cells, preservation of skeletal muscle is not excluded. Previously, Rochkind et al. suggested that laser phototherapy on intact muscle and muscle cells 18 could have a therapeutic effect that induced biochemical changes, which might be a trophic signal for increased activity of CK, thus preserving a reservoir of high energy phosphate available for quick resynthesis of ATP that would enable the survival of AChR.
The present study applied an experimental model 8 –10 to test the impact of laser therapy on complete muscle denervation in the rat gastrocnemius muscle. This recently standardized model has been used to characterize muscle survival, by detecting AChR and CK activity at different time points (7, 14, 21, 30, 60, 120, and 210 days). Previously, we had focused on the effectiveness of laser phototherapy in treating severely injured peripheral nerves, and potential applications for denervated muscles. 18,19 In this study, we investigated the response of denervated muscles to laser phototherapy at different temporal milestones, to better characterize the cellular mechanisms of laser–muscle interaction.
Material and Methods
Animals and surgical procedure
The hindlimbs of the rats were denervated when the animals were 4 months of age, at which time they had already completed their major growth phase. Denervation at earlier ages, especially during the immediate postnatal period, involves additional variables, including death of severed motor neurons and decreased stability of the muscle fibers. 20,21 This time frame was selected to avoid affecting the skeletal muscles during the period of postnatal instability.
The animal protocol was approved by the Ethics Committee for Animal Care of the Tel-Aviv Sourasky Medical Center. Ninety-six male Wistar rats, 4 months old, weighing ∼250 g, were used. Before and after the operation the animals were kept in plastic cages in rooms with a 12 h light–dark cycle, with free access to food and water. The animals were anesthetized with an intraperitoneal injection of xylazine (20 mg/kg) and ketamine (100 mg/kg). The right thigh of each rat in each group was prepared for an aseptic procedure.
After exposure of the right sciatic nerve by dissecting it from the biceps femoris and semimembranosus muscles, 10 mm of nerve segment was removed. The muscular and subcutaneous layers and skin were closed with 3/0 sutures. Removal of the sciatic nerve segment resulted in complete denervation of the muscles.
Laser treatment
The rats underwent laser irradiation (HeNe continuous wave [CW] laser, 632.8 nm, 35 mW, 30 min) every day, for 14 days. Low-power laser irradiation was delivered transcutaneously to the right gastrocnemius muscle. The therapeutic procedure started immediately after surgery. All animals were treated in the same manner. In order to maintain a similar treatment protocol both laser irradiated and unirradiated rats were anesthetized.
AChR and CK activity measurement
The sample consisted of 96 rats, 48 of which were treated with laser and 48 of which were untreated. Under general anesthesia, both the treated and untreated rats were euthanized at seven time points: day 7 (10 rats), day 14 (10 rats), day 21 (10 rats), day 30 (5 rats), day 60 (4 rats), day 120 (5 rats), and day 210 (4 rats). AChR concentration was measured on days 7, 14, 21, 30, 60, 120, and 210, in both the control and laser-irradiated muscles, by exposing the gastrocnemius muscle homogenate to 6×10−8 M 125I-α-bungarotoxin (50–150 Ci/mmol) for 1 h in tris buffer, as described elsewhere. 22,23 Unbound toxin was removed by six washings with 2 mL tris buffer. Bound activity was counted by laying filters on a flat crystal 2.54-cm diameter γ counter.
CK activity was measured by a specific spectrophotometrical method 24,25 on the same homogenate as AChR, using a spectrophotometer at 340 nm with a Sigma kit at 7, 14, 21, 30, 60, 120, and 210 days in both intact (control) and laser-irradiated intact and denervated muscles.
Analysis and statistics
All the statistical analyses and calculations were done using MatLab software (Ver. 2008b, The MathWorks, Inc.). We used nonparametric statistics since the number of rats at each sample time point was too small to evaluate a normal distribution. The figure presentation is aligned with our statistics; and expresses the data as the median±MAD. All significance levels were calculated using a Mann–Whitney U test when the tests were independent observations (for example, when comparing the treated vs. the untreated rats) and a Wilcoxon signed-rank test when comparing two related samples (for example, when comparing the data for the denervated and intact leg of the same rat).
Correlations between muscles in the same rat (laser irradiated and unirradiated) were calculated to evaluate the dynamic change of AChR and CK over time.
Results
Two groups of rats were used in this study, one group that was laser-irradiated (laser group) and another that was untreated (control group). Two muscle survival indicators were explored: AChR, which plays a key role in neuromuscular transmission, and CK, which is an important enzyme for supplying a source of energy to the muscle. In each rat we measured AChR and CK from both legs: the leg that underwent denervation (denervated muscle) and other that remained intact (intact muscle). To identify changes in AChR and CK and to address ongoing changes resulting from the laser treatment, we conducted measurements at different time periods (7, 14, 21, 30, 60, 120, and 210 days) which cover a long-term period in the rat's life expectancy. Each rat was euthanized at a specific time period according to protocol and AChR and CK were measured. Follow-up was impossible, as our measurement procedure included a large portion of muscle. Samples were taken as follows.
AChR control group
Forty-eight double samples (48 for denervated and 48 for intact) at seven time points (day 7 [n=10], day 14 [n=9], day 21 [n=10], day 30 [n=5], day 60 [n=5], day 120 [n=5], and day 210 [n=4]).
CK control group
Thirty-one double samples (31 for denervated and 31 for intact) at seven time points (day 7 [n=4], day 14 [n=4], day 21 [n=5], day 30 [n=5], day 60 [n=5], day 120 [n=5], and day 210 [n=3]).
AChR laser group
Forty-eight double samples (48 for denervated and 48 for intact) at seven time points (day 7 [n=10], day 14 [n=10], day 21 [n=10], day 30 [n=5], day 60 [n=4], day 120 [n=5], and day 210 [n=4[).
CK laser group
Thirty-three double samples (33 for denervated and 33 for intact) at seven time points (day 7 [n=5], day 14 [n=5], day 21 [n=5], day 30 [n=5], day 60 [n=5], day 120 [n=5], and day 210 [n=3]).
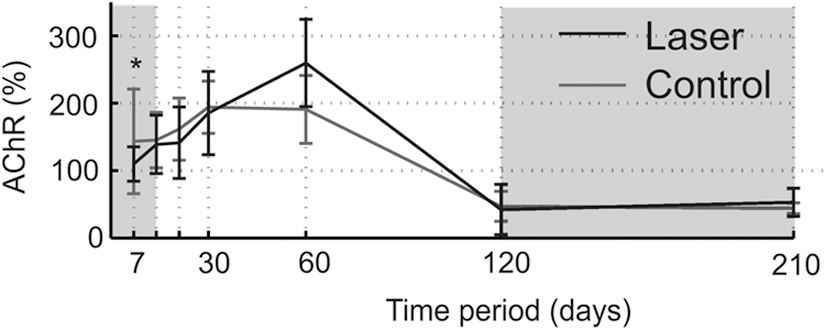
Figure 1 shows differences between groups in the amount of AChR in the denervated muscle versus the intact muscle. Note that similar local trends were observed between the denervated (solid line) and intact muscle (dashed line) within each group because of the double sampling of the denervated and the intact leg at each time point. The AChR from the denervated muscle had a biphasic effect compared with the intact muscle. Specifically, AChR in the denervated muscle exceeded that in the intact muscle AChR from day 30 to day 60 and then decreased below the intact muscle AChR (days 120–210). This biphasic effect was found in both the laser and control groups. To quantitatively assess the difference in AChR between the denervated and intact muscles, several times periods were pooled.

Longitudinal level measurements of acetylcholine receptors (AChR) in intact and denervated gastrocnemius muscle in rats that underwent laser irradiation (laser group, black lines) and rats that had no additional treatment (control group, gray lines). Laser treatment was administered for 14 days (time frame of 0–14 days) denoted by the dark gray background. Important time frames in which the portions of AChR in the denervated muscle were above or below the AChR compared with the intact muscle are denoted by the light gray background. Data were calculated for median±median absolute deviation (MAD). Significant difference between the denervated muscle and the intact muscle are indicated by black (laser group) and gray (control group) asterisks. Asterisks denote significant p values (*p<0.05; **p<0.01).
Figure 2 compares the AChR in the denervated and intact muscles for days 0–21, 30–60, and 120–210. The biphasic effect shown in Fig. 1 was statistically significant: the AChR in the denervated muscle was significantly higher for days 30–60 and significantly lower for days 120–210. Interestingly, there were significant differences in AChR for days 0–21. In the laser group, AChR in the denervated muscle decreased significantly compared with the intact muscle; whereas in the non-laser-treated group, the AChR in the denervated muscle increased significantly (as expected after denervation) in comparison with the intact muscle (control).

Acetylcholine receptors (AChR) median level in the non-laser-treated and treated rats at three time frames, as denoted by the gray background in Figure 1. Level of AChR median in three time frames (0–21, 30–60, 120–210). Data were calculated for the median AChR level. Error bars indicate median absolute deviation (MAD) for each time frame. Asterisks denote significant p values (*p<0.05; **p<0.01; ***p<0.001). The percent change from the denervated muscles is noted (gray, control group; black, laser group). Nonsignificant changes are omitted.
To test the differences between the laser group and the control group, we calculated the relative changes in AChR in the denervated muscle versus the intact muscle (the intact muscle was considered the reference within each group separately) (Fig. 3). This process eliminates errors from direct comparison of different animals (for example comparison between AChR of denervated muscles in the laser versus the control group), and thus increases the reliability of the results. In this analysis, 100% was the baseline; a relative change above this line indicated that the AChR in the denervated muscle exceeded the AChR in the intact muscle, and vice versa. As expected from previous results, there was a clear biphasic pattern in both groups, with an increase in AChR at days 30–60 and a decrease at days 120–210. There was a significant difference between the laser and the control groups in the early time period of day 7. To quantify this difference, we calculated the relative change for the 0- to 21-day time frame (Fig. 4). The results showed a significant relative change in the laser treatment group; namely, lower AChR in the denervated leg than in the control. It is well known that in a denervated muscle, the AChR increases prior to muscle degeneration. 7 These findings may, therefore, suggest a protective effect of laser treatment that can slow the increase in AChR in the denervated muscle, which typically occurs soon after denervation.
Acetylcholine receptors (AChR): relative change in control group (gray line) and laser group (black line). Indicates a relative change of the denervated muscle from the intact muscle in the same rat. Asterisks denote significant p values (*p<0.05).
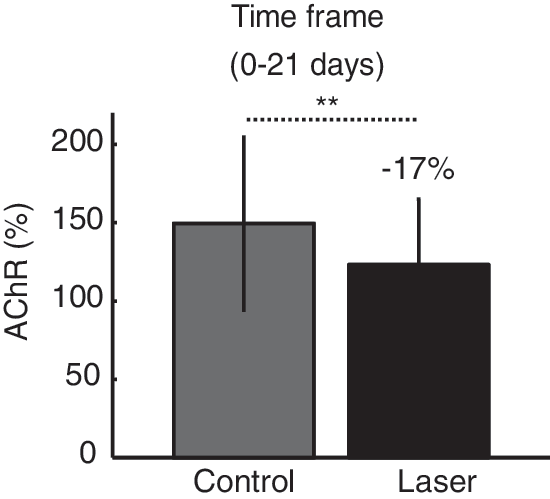
Acetylcholine receptors (AChR): relative change during first 21 days (time frame 0–21 days). In order to evaluate the difference, we compared the relative change of AChR in the laser group (black bar) to the control group (gray bar). Significant effects are presented with asterisks and denote significant p values (**p<0.01).
CK content in the denervated muscle was compared with that in the intact muscle (Fig. 5). In general, from the 21-day time point to the last measured time period, there was less CK in the denervated muscle than in the intact muscles (Fig. 5 top). This effect was found for both the laser and control groups. To assess the effect of laser treatment on CK, the relative change in the denervated and intact muscle was calculated (Fig. 5 bottom). Surprisingly, at the 30-day time point, the relative change was significantly lower in the control group; that is, there was a higher proportion of CK in the denervated muscle during laser treatment. This result was further confirmed by inspecting relative change in CK at the 30-day time period (Fig. 6) where the data showed that all subjects in the laser group had lower relative changes than in the control group.
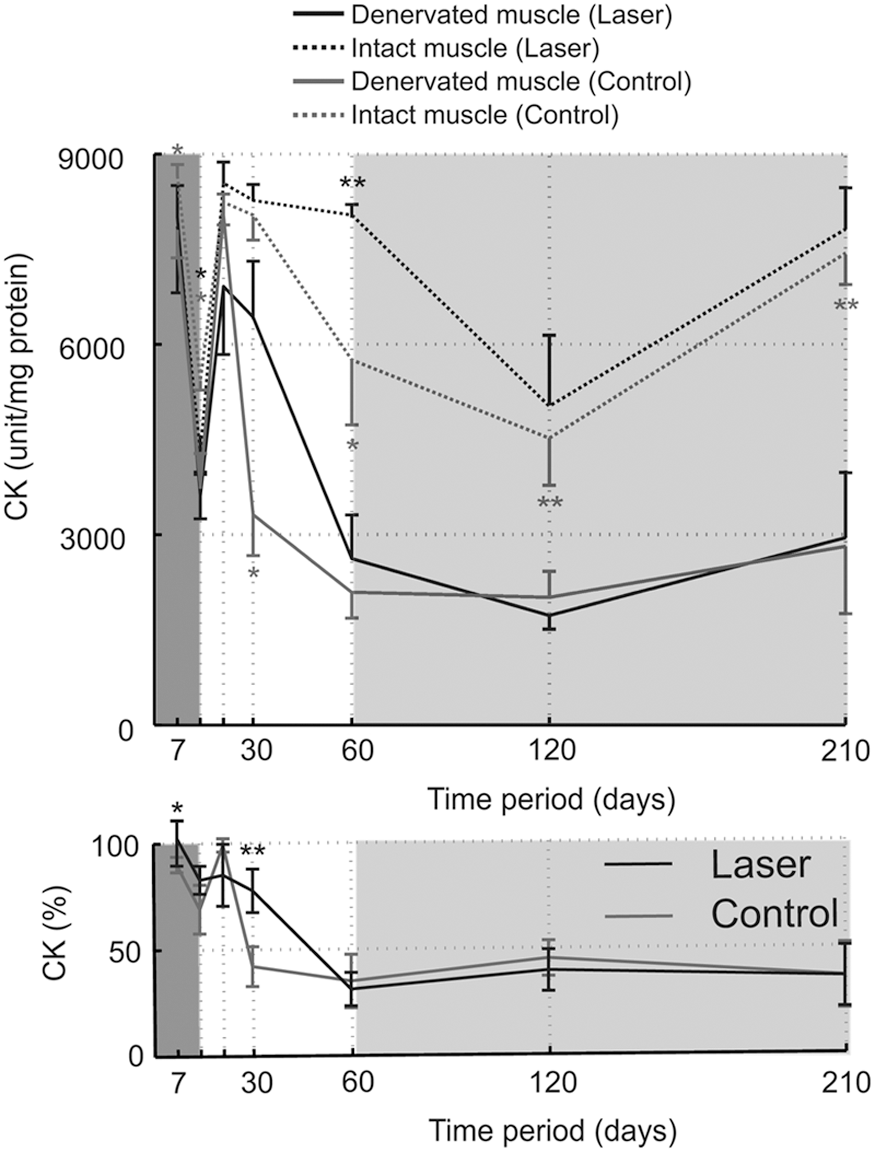
Upper panel: longitudinal level measurements of creatine kinase (CK) in intact and denervated rat gastrocnemius muscle in rats that underwent laser irradiation (laser group, black lines) and rats that had no additional treatment (control group, gray lines). Laser treatment was administered for 14 days (time frame of 0–14 days) denoted by the dark gray background. Key time frames in which the portions of CK in the denervated muscle were below the CK compared with the intact muscle are denoted by the light gray background. Data were calculated for the median±median absolute deviation (MAD). Significant differences between denervated muscle and intact muscle are indicated by black (laser group) and gray (control group) asterisks. Asterisks denote significant p values (*p<0.05; **p<0.01). Lower panel: CK relative change in the control group (gray line) and laser group (black line). Indicates relative change of the denervated muscle from the intact muscle in the same rat. Asterisks denote significant p values (*p<0.05; **p<0.01).
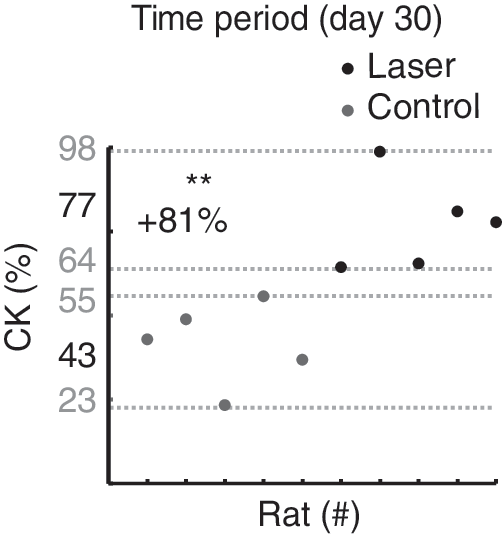
Creatine kinase (CK) relative change in control and laser groups at day 30. To illustrate the laser effect, the relative change in individual rat CK (each dot represents a rat) is shown as a scatter plot. This graph shows that laser-treated rats (black) and untreated rats (gray) are fully separated, indicating a robust effect of laser on CK at this time point (**p<0.01).
Discussion
Post-traumatic prevention of muscle atrophy is a major challenge in restorative medicine. When muscles are denervated, as in cases of complete peripheral nerve injury, they deteriorate progressively. Although some muscle regeneration does occur, 26 it is at a level insufficient to replace the degenerative loss.
Prolonged denervation causes multiple functional and morphological changes in skeletal muscle because of the absence of motor and trophic regulatory control by the nerve. The most prominent features of denervated skeletal muscles are the rapid atrophy of muscle fibers and a decrease in the number of both myonuclei and satellite cells. 2 –5 One of the main explanations for the incomplete restoration of very long-term denervated muscle is a failure of regenerating nerves to reach all of the atrophic muscle fibers and establish mature muscle–nerve contacts. 27 –29
Previous studies have shown that the sites of former end plates could be detected in surviving skeletal muscle fibers even after 17 months, 27 and as much as 2 years following muscle denervation. 30 Although these findings extend the time of preservation of former end plate sites on the surface of atrophic muscle fibers considerably, it is very difficult to assess the functional condition of these long-term denervated sites of former neuromuscular junctions with respect to their capacity to accept growing axons if re-innervation were to occur. If not re-innervated, the regenerating myofibers undergo atrophy and degeneration. 31 To lessen or temporarily prevent this process, especially in cases of complete peripheral nerve injury in which the affected nerve is reconstructed by grafts, tube, or primary anastomosis, laser phototherapy may be an effective tool that preserves denervated muscle until nerve sprouting into the muscle occurs. The early protective effect of laser treatment on denervated muscle 8 and in an experimental ischemia perfusion muscle injury model has been previously described. 32,33 Late denervation has been widely studied in animal models. In rats, it has been shown that for the first 7 months after denervation, myofibers exhibit a net loss of nuclear domains followed by nuclear groupings. 34
Conclusions
In the present study, we found that laser irradiation on the denervated muscle had a significant protective effect at two time periods: during the first period of 21 days for AChR and at 30 days for CK activity. We found that in early stages of muscle degeneration, laser treatment temporarily preserved AChR and CK in the denervated muscle close to its initial level before injury, and partially maintained CK activity and the amount of AChR in the denervated muscle during the consecutive stages of muscle degeneration. This laser effect confirms our previous results on muscle cells and intact muscles. 18 Hence laser phototherapy, by tending to impede the increase in AChR and the decrease in CK after denervation, has a potential for future therapeutic applications.
Footnotes
Acknowledgments
We thank Einav Ivanov for assistance in the preparation of this article, and Dr. Elhanan Meirovithz for the data analysis and statistics.
Author Disclosure Statement
No competing financial interests exist.