
Abstract
Introduction
A
However, attitudes toward the use of both polychromatic and monochromatic irradiation in medical practice remains reserved, because of fears about the appearance and/or stimulation of neoplastic processes. Such fears are based on the known biostimulation effects of VIS and IR irradiation and, specifically, on their ability to accelerate proliferation not only in normal cells, but also in tumors. However, the effect of irradiation of optic diapason on the proliferation of tumor cells under conditions in vitro, and on the growth of malignant tumors in laboratory animals in vivo are far from being resolved. It has been shown that different wavelengths of low-power laser irradiation can stimulate cell proliferation in culture, 6 –12 have no effect on proliferation of transformed cells, 10,12 or even inhibit proliferation. 13 Low-power laser irradiation can stimulate the growth of transplanted malignant tumors in laboratory animals, 14 but many more studies demonstrate either the absence of the stimulating effect of light on this process, or its inhibition. 15,16 The maximal antitumor effect developed either after direct irradiation of tumors, 15,17 or after irradiation in animals in the area of tumor transplantation. 15
A possible cause for change in the tumor growth rate might be a reorganization of the tumor cell cytoskeleton. 18,19 Indeed, after irradiation with IR using an 808 nm GaAlAs laser, at doses of 5.85 and 7.8 J/cm2 there was a disturbance of the regular structure of cell microtubules of human hepatoma cells in culture. 13,16 This is indicative of the inhibition of cell proliferation and subsequent cell destruction. 16 Lack of information in the literature on the effect of polychromatic VIS and IR irradiation on tumorigenity and the actin cytoskeleton of tumor cells prompted the current study. The proliferation and tumorigenic properties of MH22 cells can be studied both in vitro and in vivo after transplantation to singenic C3HA mice, and was used in this study.
Materials and Methods
Sources of irradiation
The source of combined VIS-IR light used in this study was the Bioptron-2 in the spectral region of 480–3400 nm, 40 mW/cm2, 95% polarization (BioptronAG, Switzerland). For irradiation, doses of 4.8 and 9.6 J/cm2 were used at exposure times of 2 and 4 min.
Q–light (P&B AG, Switzerland) was used for the complete VIS spectrum (380–750 nm, 40 mW/cm2, 95% polarization). For irradiation, doses of 9.6 and 38.4 J/cm2, were used at exposure times of 4 and 16 min.
Experimental animals
C3HA mice weighing 18–20 g were used in experiments (nursery “Rappolovo,” Russian Academy of Medical Sciences). All animal experiments were approved by the local animal ethics committee in accordance with institutional guidelines for the welfare of animals.
Cells of mouse hepatoma
The line-specific hepatoma 22a is the substrain of hepatoma 22, induced with orthoaminoazotoluene in C3HA mice in 1951. Cytological studies indicate that this tumor is of the hepatocellular origin ascribed to anaplastic carcinomas. The murine hepatoma (MH)-22a cells were obtained from the bank of cell cultures of the Institute of Cytology of the Russian Academy of Sciences. Cells were cultivated in Dulbecco's Modified Eagle Medium (DMEM) (Biolot, Russia) with addition of 10% fetal calf serum (Biolot, Russia) and 40 μg/mL gentamicin (Sigma, USA).
Irradiation of hepatoma MH-22a cells in vitro
Hepatoma MH-22a cells were irradiated with VIS-IR or with VIS light. For irradiation, the hepatoma MH-22a cells were seeded into six well plates, and 2×105 cells were seeded per well to achieve the logarithmic growth phase. Before irradiation, the culture medium was replaced by phosphate-buffered saline (PBS), and after irradiation, it was replaced again with medium. Twenty-four hours after irradiation, the cells were transplanted into C3HA mice. In parallel, the cells were characterized by criteria of viability, proliferative activity, and the state of their actin cytoskeleton.
Transplantation of hepatoma MH-22a cells
The hepatoma MH-22a cells were transplanted subcutaneously into the area of the back to intact C3HA mice at 2×105 cells per mouse. The time and appearance of tumors as well as their size and the time of death of the mice were recorded every 5 days for 10–40 days after transplantation of tumor cells. The volume (V) of tumors was calculated by the rotation ellipsoid formula
where L=maximal diameter of the tumor, and D=minimal diameter (mm). Three series of experiments were performed, and 150 mice were divided into five corresponding groups. Groups 1 and 2 were transplanted with cells irradiated with VIS-IR light at doses of 4.8 and 9.6 J/cm2, respectively, whereas groups 3 and 4 were irradiated with VIS light at doses of 9.6 and 38.4 J/cm2, respectively. Group 5 (control) was administered with unirradiated cells.
Evaluation of proliferative activity and viability of MH-22 cells by flow cytometry
Viability and proliferative activity of cells were evaluated on an EPICS XL flow cytometer (Beckman Coulter, USA). Unirradiated cells served as control. To calculate the number of dead cells, propidium iodide (Invitrogen, USA) was added to the cell suspension at a concentration of 0.05 mg/mL. After incubation for 5 min at room temperature, each sample was analyzed. The percentage cells stained with propidium iodide relative to the total number of cells was determined.
For analysis of the cell cycle, saponin (Fluka, USA) was added at a concentration of 0.2 mg/mL, RNase (Sigma, USA) was added at a concentration of 0.25 mg/mL, and propidium iodide (Invitrogen, USA) was added at a concentration of 0.05 mg/mL. After incubation for 30 min at room temperature, the samples were analyzed on a flow cytometer. The data were processed with ModFit LT software (Verity Software House, Topsham).
Staining of actin microfilaments
Twelve hours prior to experiment, 4×105 MH-22a cells were seeded onto Petri dishes with cover slips and washed three times with PBS. The cover slips were transferred onto parafilm, fixed with 3.7% paraformaldehyde (Sigma), and rinsed three times with PBS. For visualization of actin microfilaments, cells were stained with rhodamine falloidin for 15 min at 37°C in an atmosphere of 5% CO2. The preparations were rinsed three times with PBS, and embedded in Fluoroshield™ medium. Intensity of staining of preparations was estimated on an AxioObserver confocal microscope (Carl Zeiss, Germany). Images were processed using ImageJ software.
Statistical analysis
Statistical analysis was performed using Wilcoxon nonparametric criteria. Significance of results on the survival of mice and transplantation of cells were determined with use of the Fisher criterion. Statistical processing of results was performed using Statistica 6.0. Differences among groups were considered significant at p≤0.05.
Results
Effect of polychromatic light on tumorigenicity of hepatoma cells
Tumor cell proliferation or transplantability of tumor cells, dynamics of tumor growth, and survival of animals were determined. The hepatoma cells were irradiated with VIS-IR or VIS light 24 h prior to their transplantation into mice.
The proliferation of tumor cells or transplantation success of irradiated cells was evaluated from the number of animals with tumor development (Fig. 1). In the control group of mice, when unirradiated cells were transplanted, tumors developed in 100% of mice by day 10. Proliferation of cells irradiated with VIS-IR light at doses of 4.8 and 9.6 J/cm2 led to a decrease in number of tumor cells at day 10, to 78% and 72%, respectively; however, the number of animals with tumors gradually increased at day 40 by 90–95% (Fig. 1). Proliferation of cells irradiated with VIS light at doses of 9.6 and 38.4 J/cm2 at day 10 increased to 90% and 94% respectively. Tumor development of cells irradiated with VIS light at a dose of 9.6 J/cm2 remained unchanged (90%) throughout the entire term, whereas after irradiation at a dose of 38.4 J/cm2, the tumors decreased from 94% to 78% by day 40, and in 22% of animals the tumors were resolved.
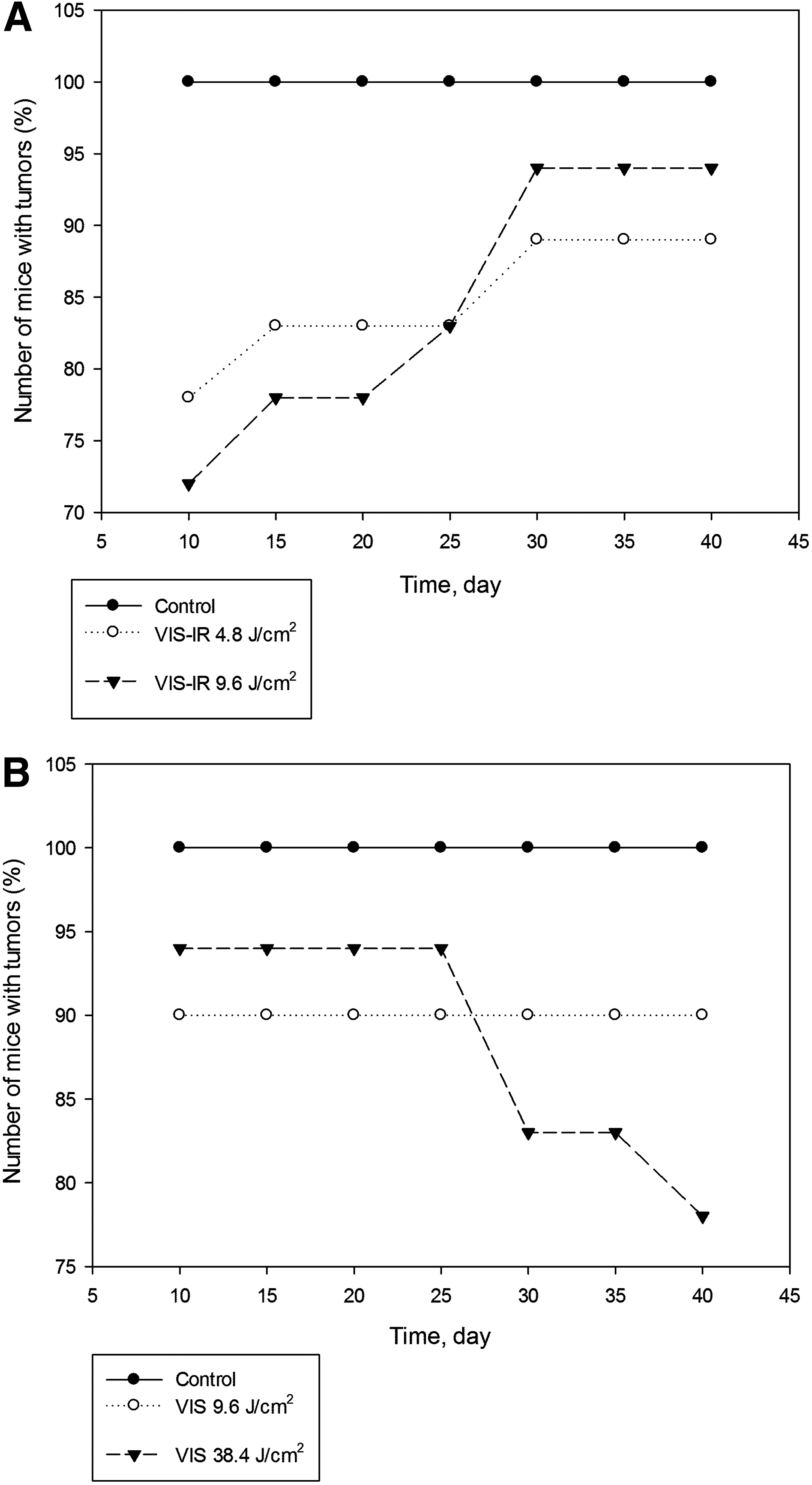
Number of mice with tumor development after transplantation of MH-22a cells irradiated with
Studying the dynamics of tumor growth showed that after irradiation with the VIS-IR light at a dose of 9.6 J/cm2, the tumor volume was 1.8 times lower than in the controls at day 10, and 16.2 times lower than the controls at day 25, whereas after irradiation at a dose of 4.8 J/cm2, it was 3.5 and 3.9 times lower, respectively. Irradiation of hepatoma cells with the entire spectrum of VIS light at a dose of 9.6 J/cm2 did not affect the tumor growth rate at any time of observation. However, after using of the maximum dose of the VIS light (38.4 J/cm2), tumor growth inhibition was observed only from day 25 after transplantation of tumor cells (Fig. 2).
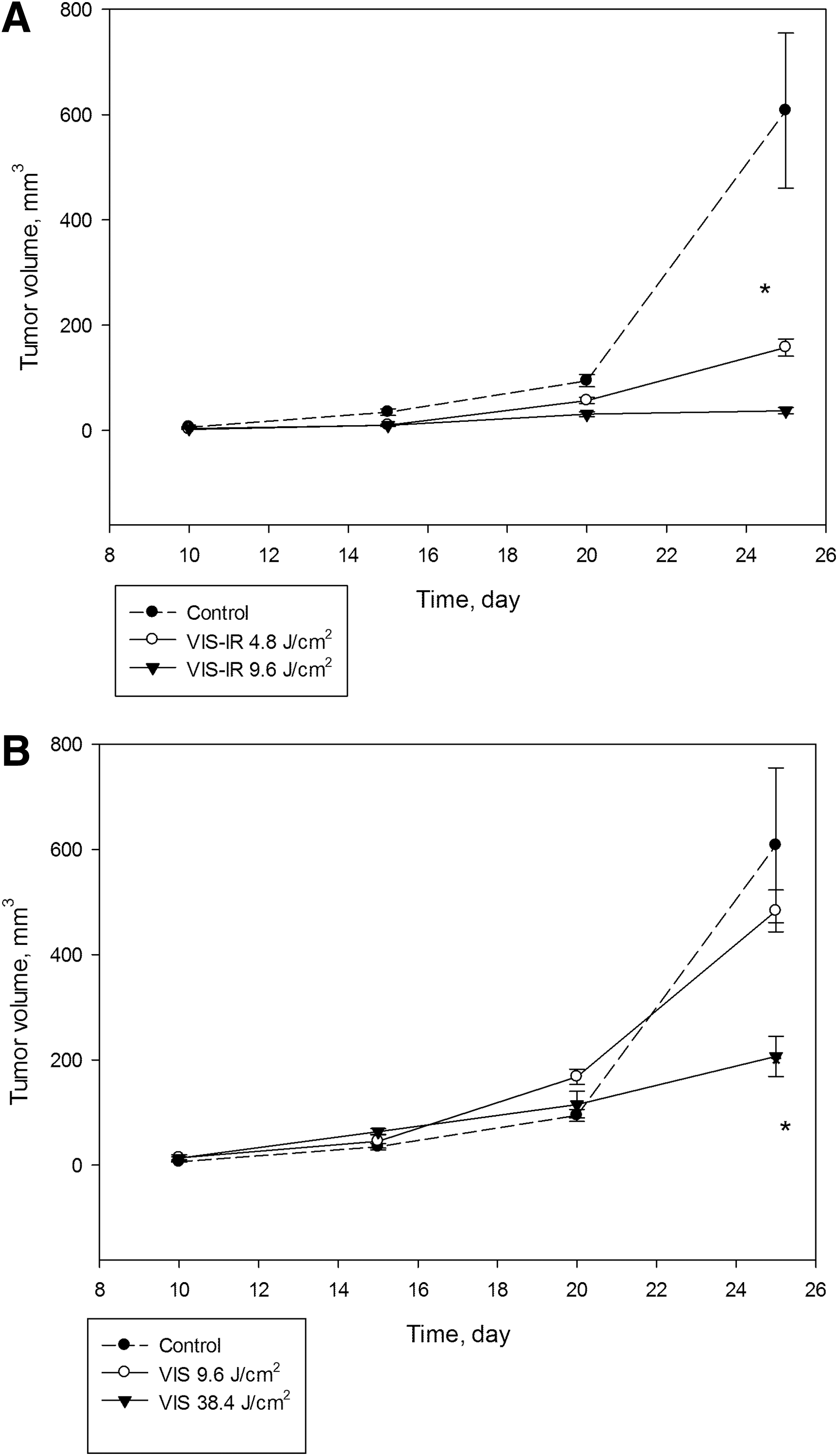
Dynamics of tumor growth in mice after transplantation of MH-22a cells irradiated with
When looking at the survival rate of animals after transplantation of the MH-22a cells. it was seen that transplantation of hepatoma cells irradiated with VIS-IR animals survived significantly longer than in the control group (Fig. 3). In the control group, the mice began to die at day 20, whereas the mice with transplanted cells irradiated with VIS-IR light at doses of 4.8 and 9.6 J/cm2 only died beginning at day 30. At day 25, mice with tumors that received IR-irradiated cells had 0 mortality compared with 27% mortality in the control group. At day 40, 72–83% of mice survived, whereas only 41% in the control group survived. In the case of irradiation of tumor cells with VIS light at a dose of 9.6 J/cm2, no differences in the survival of mice were revealed. Irradiation of cells with VIS light at a dose of 38.4 J/cm2 led to the survival of mice at day 40 being increased (72% versus 41% of controls). Among the mice that survived, there were animals with completely resolved tumors.
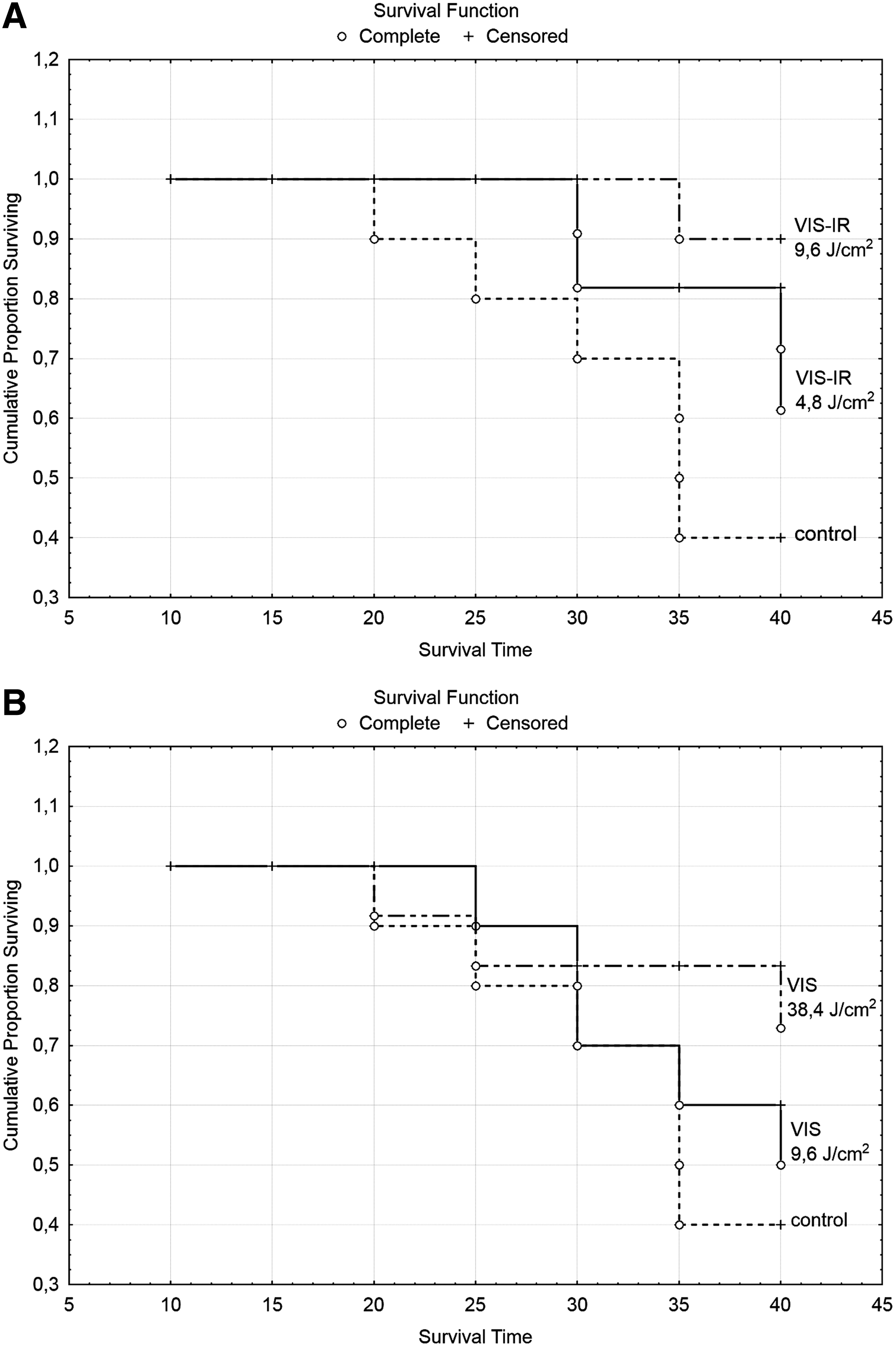
Survival rate of mice with tumors after transplantation of MH-22a cells irradiated with
Effect of polychromatic light on proliferation and viability of hepatoma cells
At the day of transplantation, the percentage of dead MH-22a cells in culture amounted to 7.6±0.5%. Irradiation of hepatoma cells with two kinds of polychromatic light did not affect the portion of viable MH-22a cells for 24–72 h. No statistically significant changes were observed (Table 1).
No statistically significant changes were observed.
A possible cause for change of tumorogenicity of MH-22a cells after their irradiation with VIS-IR and VIS light might have been deceleration of cell proliferation. To determine the viability of this, the cell cycle phases at 24, 48, and 72 h after irradiation were studied. Irradiation did not change the cell cycle distribution and the portion of cells in each cycle phase remained without significant changes within the same boundaries (Fig. 4).
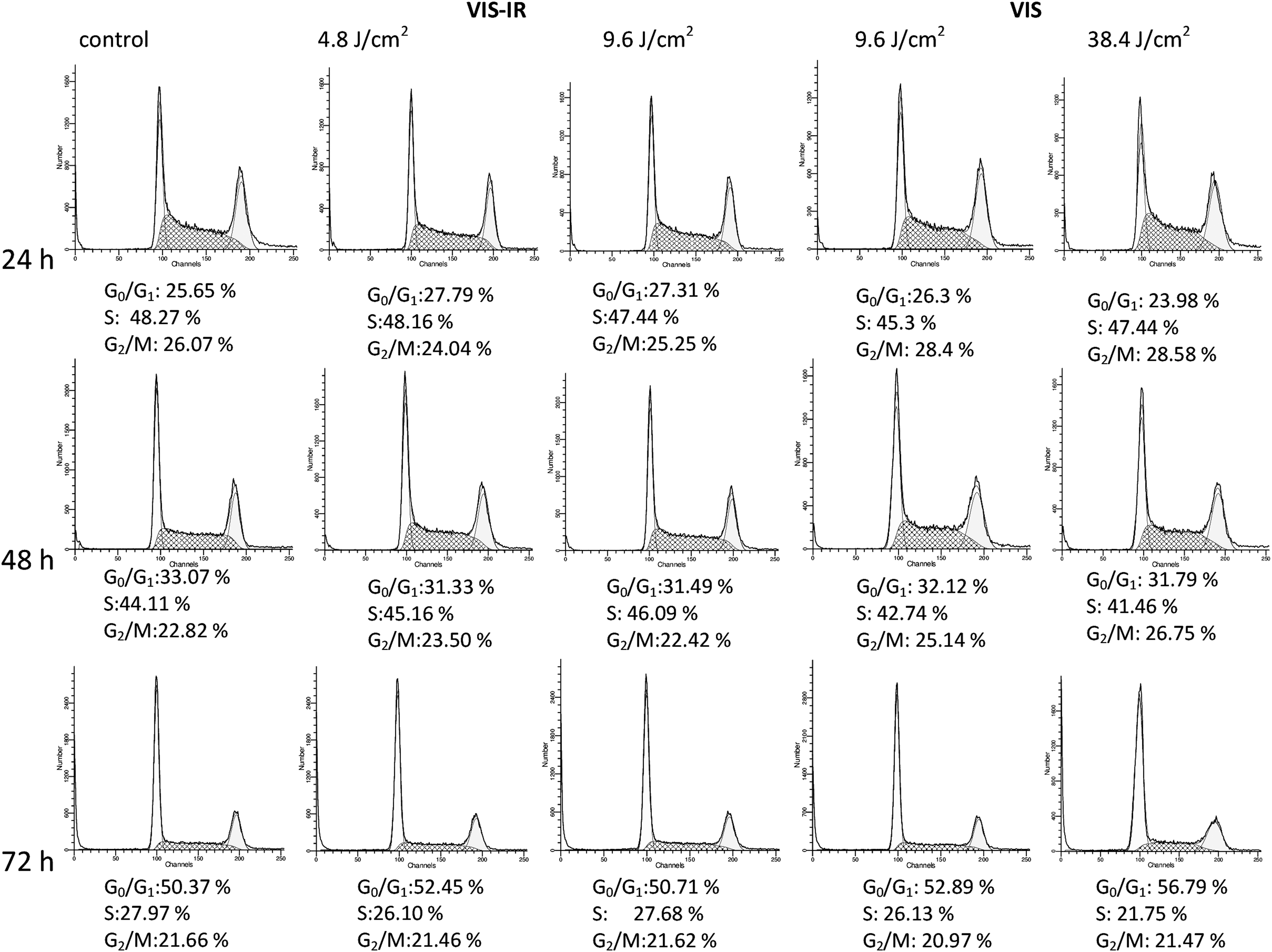
Effect of visible (VIS)-infrared (IR) irradiation on passage of cell cycle phases by MH-22a cells 24, 48, and 72 h after irradiation with VIS-IR and VIS light in different doses.
Effect of the polychromatic light on structural state of the actin cytoskeleton of hepatoma cells
Evaluating the actin cytoskeleton of unirradiated cells (controls) in vitro, three cell populations were identified (Fig. 5). The first population (A) included cells with clearly pronounced actin fibrils, and accounted for 55% of the total number of cells. In two other populations, the actin cytoskeleton was disorganized; however, this “disorganization” was of a different character. Population B, amounting to 40% of the total number of the unirradiated cells, was characterized by localization of actin predominantly in the cytoplasm. In population C, actin was located mainly near the membrane, and this portion of cells did not exceed 5%.
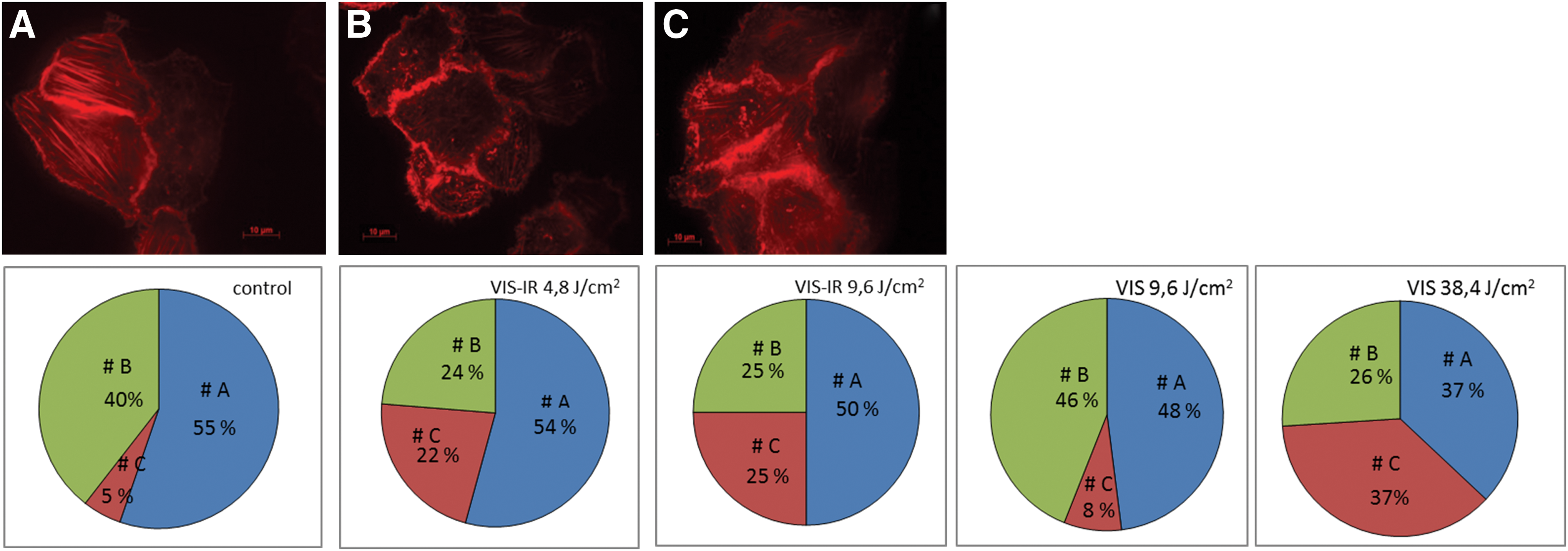
State of actin cytoskeleton of MH-22a cells in vitro, irradiated with VIS-IR and VIS light.
After irradiation of hepatoma cells with VIS-IR light, at doses of 4.8 and 9.6 J/cm2, the amount of cells in population A (54% and 50% vs. 55% in controls, respectively) remained unchanged. The number of cells in population C increased sharply (five times) at the expense of their decrease in population B. After exposure to VIS light at a dose of 9.6 J/cm2, the portion of cells in populations A, B, and C remained similar compared with the controls at 48%, 46%, and 8%, respectively. However, significant cytoskeleton changes were observed after irradiation of hepatoma cells with VIS light at a dose of 38.4 J/cm2, when cell distribution changed to 37% (A), 26% (B), and 37% (C). At this dosage, a decrease in cell tumorigenity was observed, and complete disappearance of the already formed tumors occurred.
Discussion
The results of the present study indicate that irradiation of hepatoma cells with VIS-IR or only VIS light at a wide range of doses did not stimulate tumor growth and did not enhance proliferation/transplantability of cells after transplantation into mice. Moreover, irradiation with VIS-IR light at doses of 4.8 and 9.6 J/cm2 led to significant inhibition of hepatoma growth rate and decreased transplantability of tumor cells. The survival of animals increased significantly in irradiated transplanted hepatoma cells. However, this effect was not observed after irradiation of tumor cells with VIS light at a dose of 9.6 J/cm2, and only irradiation of cells at a dose of 38.4 J/cm2 led to inhibition of the tumor growth rate at 25–40 days after transplantation of hepatoma cells.
Our results agree well with studies that revealed a decrease in the tumor growth rate after direct irradiation in situ or after action of light on the area of transplantation of tumor cells soon after their injection into animals. 15,17 In these studies, red light of He-Ne or ruby lasers (633 and 694 nm, respectively) were used as irradiation source. In these studies, the course of irradiations used was of different durations and irradiation doses varied from 40 to 70 J/cm2. In several studies, the antitumor effect of irradiation was recorded after using doses of irradiation that stimulated proliferating processes in the animal body, such as cutaneous wound healing and hemopoiesis. 20 The degree of the growth suppression of tumors of different tissue differed and varied from 16% to 80%, up to their complete disappearance. 17
Many authors noted the antitumor effect of red, yellow, green, and blue light, as well as different wavelengths in the near-IR spectral range. 21 Therefore, based on the literature, it can be suggested that the antitumor effect of the combined polychromatic VIS and IR light used in our work can be induced with red, yellow, and green light, but not with blue light, as the short-wave component of the VIS spectrum in our VIS-IR radiation (480–3400 nm) was absent. The repeatedly reported significant antitumor potential of near-IR irradiation allows understanding of why inhibition of tumor growth is achieved in the case of the combined VIS-IR irradiation, at lower doses more than with VIS light alone.
However, inhibition of tumor growth rate under action of the low-power irradiation conditions in vivo does not elucidate the mechanisms of the detected phenomenon. Our studies were performed in a model in which cultured hepatoma MH-22a cells were irradiated and then transplanted into C3HA syngenic mice. 22 This model has allowed us not only to evaluate the effect of VIS-IR and VIS irradiations, but also to identify possible mechanisms of antitumor effects induced by polychromatic light.
Irradiation of transplantable murine hepatoma cells with polychromatic VIS-IR and VIS light at non-damaging doses did not increase frequency of tumor formation and did not accelerate their growth for 2 months after cell transplantation to syngenic mice. Moreover, transplantation of hepatoma cells irradiated with VIS-IR light was accompanied by a decrease in their proliferation/transplantability and tumor growth rate and by an increase in the animal life span compared with control animals. The polychromatic VIS light (385–750 nm) at a dose of 9.6 J/cm2 did not produce a similar antitumor effect, and a fourfold increase in this dose led to inhibition of tumor growth. From this it follows that the antitumor effect of the combined VIS-IR radiation is provided by its VIS component, but in combination with IR irradiation, the antitumor effect is enhanced. This result depended on the dose and type of irradiation, and was realized after transplantation into mice of different numbers of irradiated cells (5×104 – 2×105 cells). In the present work, the maximum number of light-irradiated cells was transplanted into the mice, and this indicated that the higher the number of irradiated cells that were transplanted, the higher the antitumor effect of light.
The tumor growth inhibition after irradiation with VIS-IR light can be the result of several things, for example the suppression of tumor cell proliferation. Studies of the effect of VIS and IR light on the proliferation of various tumor cell lines using different modes of irradiation (wavelengths and doses) indicate that there are still several remaining questions regarding the effect on tumor cell proliferation post-irradiation. 10,13,23 Our work demonstrated that irradiation with VIS-IR light did not lead to a change in the MH-22a cell proliferation after 24–72 h, nor did it cause cell death; hence neither cytotoxic nor cytostatic effects were observed. Inhibition of tumor growth rate had no direct correlation with a change in the tumor cell proliferation.
In the work of Karu and coworkers, 24 visible monochromatic light was shown to increase the capability of cells both for adhesion to each other and to glass. Because the adhesive properties of any cell and their capacity for metastasis of tumor cells depend on the structural state of their actin cytoskeletons, 18,19 we analyzed the effect of VIS-IR and VIS light on the actin cytoskeleton of hepatoma MH-22a cells.
At evaluating the actin cytoskeleton of unirradiated MH-22a cells (controls), three cell populations were revealed: cells with evidently pronounced actin microfilaments (55% of the total cell number) (A); cells with reorganized actin localized predominantly in the cytoplasm (40% of the total cell number) (B), and cells with reorganized actin most pronounced near the membrane (5% of the total cell number) (C). After irradiation of MH-22a cells with VIS light at 9.6 J/cm2, their tumorogenicity did not change in any of the three populations. However, when irradiating with VIS-IR light at 4.8 and 9.6 J/cm2, tumorogenicity was downregulated, and the number of cells in population C sharply increased (five times). Additionally, total disappearance of tumors occurred when the cells were irradiated with VIS light at 38.4 J/cm2, and the portion of cells in population C increased 7.5 times. Our data allude to the existence of a relationship between a decrease in the tumorigenity of MH-22a cells after their irradiation with polychromatic light, and an increased number of tumor cells with reorganized actin associated with membrane (population C).
Literature is scarce regarding the action of the entire spectrum of VIS and IR irradiation on the actin cytoskeleton of tumor cells. Irradiation with laser GaAlAs (808 nm; 5.85 and 7.8 J/cm2) led to a disturbance of the regular structure of microtubules of human hepatoma cells. 13,16 They ascribe destruction of actin cytoskeleton to the inhibition of proliferation of these cells. 16 The structure of actin microfilaments is changed when interacting with natural killer (NK) cells that are effectors of the system of natural antitumor resistance and are capable of lysis of the large spectrum of tumor cell targets without their prior sensitization. Several studies 25,26 indicate that actin stress fibrils and “disorganized” actin localized in the cytoplasm can interfere with the cytotoxic process. Our data indicate that the total number of cells in the population with clearly pronounced stress fibrils are not altered, and that the number of cells with disorganized actin is the result of irradiation with VIS-IR light.
Conclusions
The results of the present study argue in favor of the oncological safety of polychromatic VIS-IR light (480–3400 nm), as the direct irradiation of tumor cells at non-damaging doses and their subsequent transplantation to mice do not stimulate tumor progression. In this aspect, VIS light is less effective. Taking into account that both polychromatic VIS and polychromatic IR irradiation are the main components of the solar spectrum, and the fact that artificial light sources are widely used in current photomedicine over the world, 3,4 the results of the present study may have an important practical contribution.
Footnotes
Author Disclosure Statement
No competing financial interests exist.