
Abstract
Introduction
A
Although employment of femtosecond lasers as medical tools opens new possibilities for surgery and therapeutic treatments, the impact of their use on genetic material is not yet fully known and investigated. Such knowledge is especially relevant to UV femtosecond lasers, because radiation in the UV range has the greatest mutagenic and carcinogenic potential. 2,14,19,20 Some studies performed two decades ago and at a later date 21 –23 prompted a view that conventional UV laser systems used in eye and skin treatment and surgery are safe enough for the health of patients and should not increase the risk of genetic disorders because of comparable low DNA damage level induced in model organisms or cultured cells. In our previous work, which was the first attempt to evaluate the possible genotoxic impact of the brand-new femtosecond laser Pharos, the unlooked-for damaging effect on the DNA of irradiation with the laser was detected. 24 Therefore, the purpose of present study was to extend our previous research and to compare the effect of femtosecond and nanosecond UV laser pulses on the genetic material. It was shown that cells of inner tissues were much more susceptible to UV radiation than cells of external tissues; 20 therefore, we used murine bone marrow cells to detect and to compare the DNA-damaging effect of irradiation with different UV sources in vitro. Comet assay, or single-cell gel electrophoresis, was used for DNA damage measurement. It is known that comet assay is a highly sensitive and valid technique for assessing DNA damage, and, in the alkaline version, it reveals the presence of single-strand breaks, double-strand breaks, and alkali-labile sites. 25 –27 This test is well suited to detecting UV-induced lesions, and is widely used for DNA damage assessment in UV-irradiated cells. 28 –34
Materials and Methods
Animals
Male BALB/C mice 10–12 weeks of age and weighing 20–25 g, were used. The animals were supplied by the Animal Facility Institute of Immunology, Vilnius University (Lithuania). Ethical approval was given by the Committee of Lithuanian Animal Care and Use.
Laser irradiation
Bone marrow cells from mice were chosen as a model sample. To compare the effects of femtosecond UV laser radiation with other UV sources, two different laser systems were employed. Femtosecond UV pulses (205 nm) were obtained by generating the fifth harmonic of the solid-state Yb:KGW laser Pharos (Light Conversion, Ltd., Vilnius, Lithuania) radiation. Nanosecond Nd:YAG laser NL301 (Ekspla, Ltd., Vilnius, Lithuania) equipped with fourth and fifth harmonic generation modules was used to produce 266 and 213 nm radiation. For positive control, we used the 254 nm line of mercury discharge lamp (Carl Roth, GmbH & Co. KG, Karlsruhe, Germany) because this type of radiation is known for having a strong DNA-damaging impact. 34 –36 Parameters (beam diameter and pulse energy) of femtosecond and nanosecond lasers were adjusted to achieve a constant energy per pulse per unit area (35 μJ/cm2) at different irradiation doses. The energy parameters of irradiation are summarized in Table 1. The exposure time for each laser source was calculated by dividing the dose by the average irradiation power density.
In dose-response experiments, it is important to ensure uniform irradiation of the sample containing bone marrow cells. To that end, the beam of the nanosecond Nd:YAG laser was expanded up to 25 mm in diameter, and the suspension of the bone marrow cells (drop diameter ∼10 mm) was placed at the center of the beam, where the energy density could be considered uniform. For the femtosecond pulses, the desired energy density per pulse was achieved by tuning the beam diameter to 2.2 mm (the pulse repetition rate of the femtosecond Yb:KGW laser is higher by a few orders of magnitude, whereas the pulse energy is correspondingly lower compared with the nanosecond laser). To ensure uniform sample irradiation, the femtosecond laser beam was scanned over the sample using galvanometric scanners HurrySCAN 10 (Scanlab, Puchheim/Munich, Germany). In this way, the high repetition rate UV pulses were evenly spread out over the entire sample area.
The average power of all UV light sources in the central part of the irradiated area was measured using power meter Nova II with thermal power sensor 3A (Ophir Optronics, Inc., Jerusalem, Israel). The sensing area of the power sensor was ∼0.7 cm2 (smaller than the irradiated area). The irradiation power density was determined by dividing measured power value by the sensing area. Spectrophotometer UV-3101PC (Shimadzu, Tokyo, Japan) was used to measure the UV absorption spectra.
Treatment and preparation protocol
Animals were euthanized and both femurs were dissected from each animal. Bone marrow was collected by washing the femurs with 2 mL Dulbecco phosphate-buffered saline solution (DPBS; Biochrom AG, Berlin, Germany) into a plastic Eppendorf microtube, and 2 mL more of DPBS was then added and the marrow mixed to obtain a fine suspension. A 60 μL aliquot of suspension was transferred onto each of six microplates and spread over an area of 1 × 1 cm to irradiate by each irradiation source. To evaluate the possible effect of medium on the extent of DNA damage, the absorbance of irradiation light by distilled water, DPBS, and cells' suspension was measured (Fig. 1).
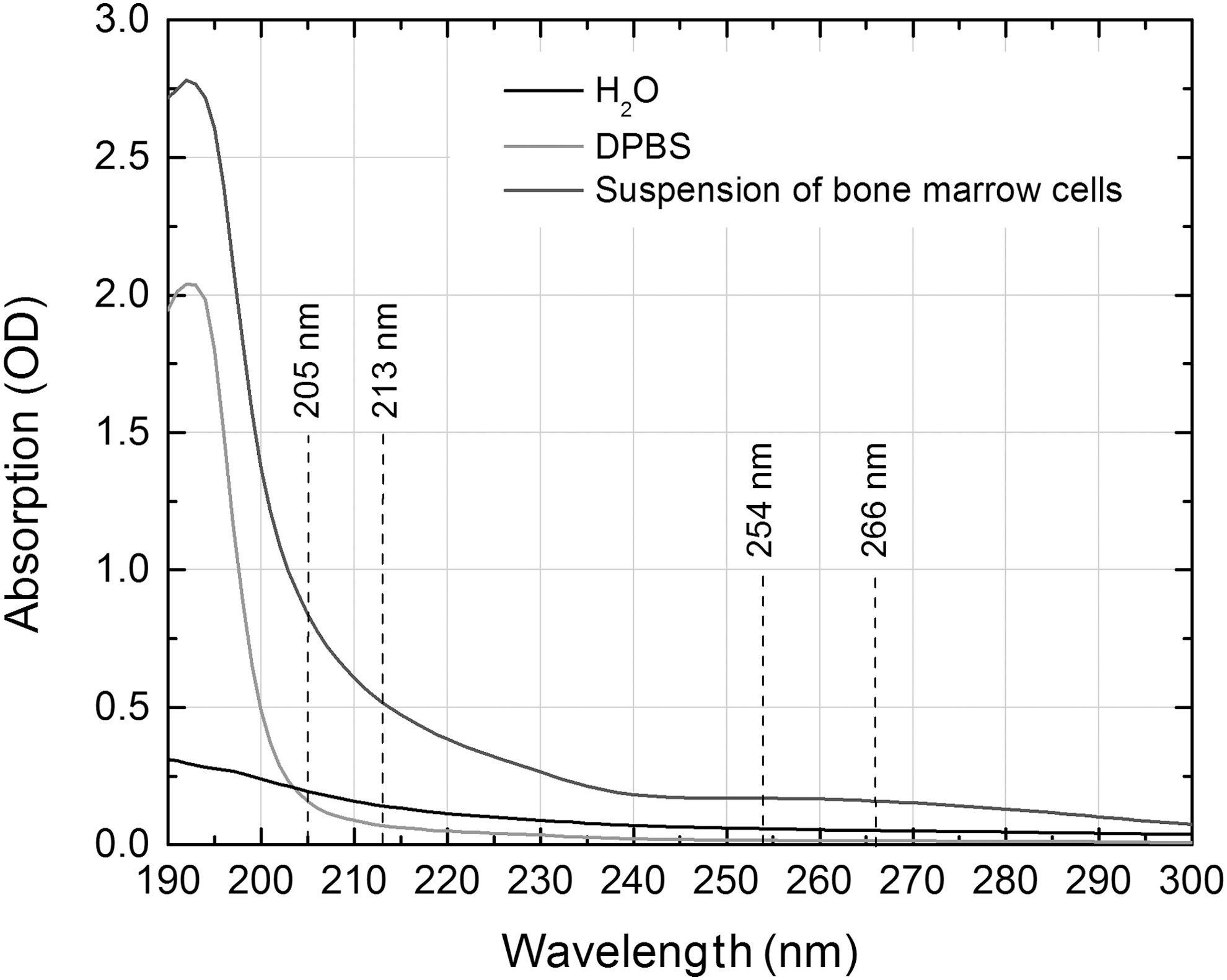
Absorption spectra of H2O, Dulbecco phosphate-buffered saline solution (DPBS), and suspension of bone marrow cells in 4 mL DPBS. The absorption of radiation by DPBS in the 205–266 nm wavelength range was weak and almost constant, whereas the absorbance of shorter wavelength light by cells' suspension was 3–4 times higher than the absorbance of 254–266 nm light.
Bone marrow cells on separate plates were exposed to different doses of irradiation: 0.00035, 0.00175, 0.0035, 0.0175, 0.035, and 0.105 J/cm2. Immediately after irradiation, 20 μL of each sample suspension was taken to assess cell viability by trypan blue exclusion assay, whereas 40 μL of the suspensions were transferred into separate Eppendorf microtubes
In brief, 40 μL of suspension from each microtube was transferred onto a microscope slide precoated with 1% standard agarose (Thermo Fisher Scientific, Vilnius, Lithuania) in DPBS and mixed with 40 μL of 1% low melting-point agarose (Carl Roth GmbH & Co. KG, Karlsruhe, Germany), and a cover glass was placed on top. The slides were refrigerated at 4°C until the gel layer solidified. The cover glass was removed, 80 μL of 1% low melting-point agarose in DPBS was added, and a cover glass was placed on top. The slides were refrigerated again at 4°C until the gel layer solidified. The cover glass was removed and the slides were placed into a jar containing lysis solution (pH 10) consisting of 2.5 M NaCl (Fluka, Riedel-de Haën, Germany), 100 mM ethylenediaminetetraacetic acid (EDTA), 10 mM Tris, 1% Triton X-100 (Carl Roth GmbH & Co. KG, Karlsruhe, Germany), and 10% DMSO (Sigma-Aldrich, Munich, Germany) for 90 min. The slides were then transferred into alkaline electrophoresis solution (pH >13) (1 mM EDTA and 300 mM NaOH [Fluka, Riedel-de Haën, Germany]) for DNA uncoiling and expression of alkaline-labile sites, where they remained for 20 min. Electrophoresis using the Comet Single Cell Gel Electrophoresis System (Scie-Plas, Ltd., Cambridge, UK) was performed at 15 V and 300 mA (0.73 V/cm) for 30 min. The slides were then placed in neutralizing solution (pH 7.5, 0.4 M Tris) for 30 min., stained with 2% ethidium bromide (Carl Roth GmbH & Co. KG, Karlsruhe, Germany) for 5 min., rinsed in distilled water, and covered with cover glasses. The analysis of the slides was performed the next day and 100 cells on each slide per treatment were scored. The computerized image analysis system Lucia (Laboratory Imaging Ltd., Praha, Czech Republic) was used to determine the percentage of DNA (%DNA) in the comet tail, as the most useful and validated index of DNA damage. Each experiment was performed 10 times using cells from the bone marrow of different mice.
Statistical analysis
The data were processed using the InStat V2.02 (GraphPad Software, CA) statistical package. Statistical analysis was performed using unpaired Student's t tests. The mean and standard error of the mean (SEM) of the average of the DNA damage score of 10 gels per experimental condition were calculated. Pearson's determination coefficient (R 2) was used in order to evaluate the dosage-effect relationship. p < 0.05 was chosen as the level of significance.
Results
The cytotoxicity assessment of irradiation of different UV irradiation sources as an integral part of the comet assay was performed using trypan blue exclusion. As is apparent from the Table 2 and Fig. 2, the effect of various parameters' UV irradiation on the viability of bone marrow cells differed, and was mostly dependent upon the wavelength.
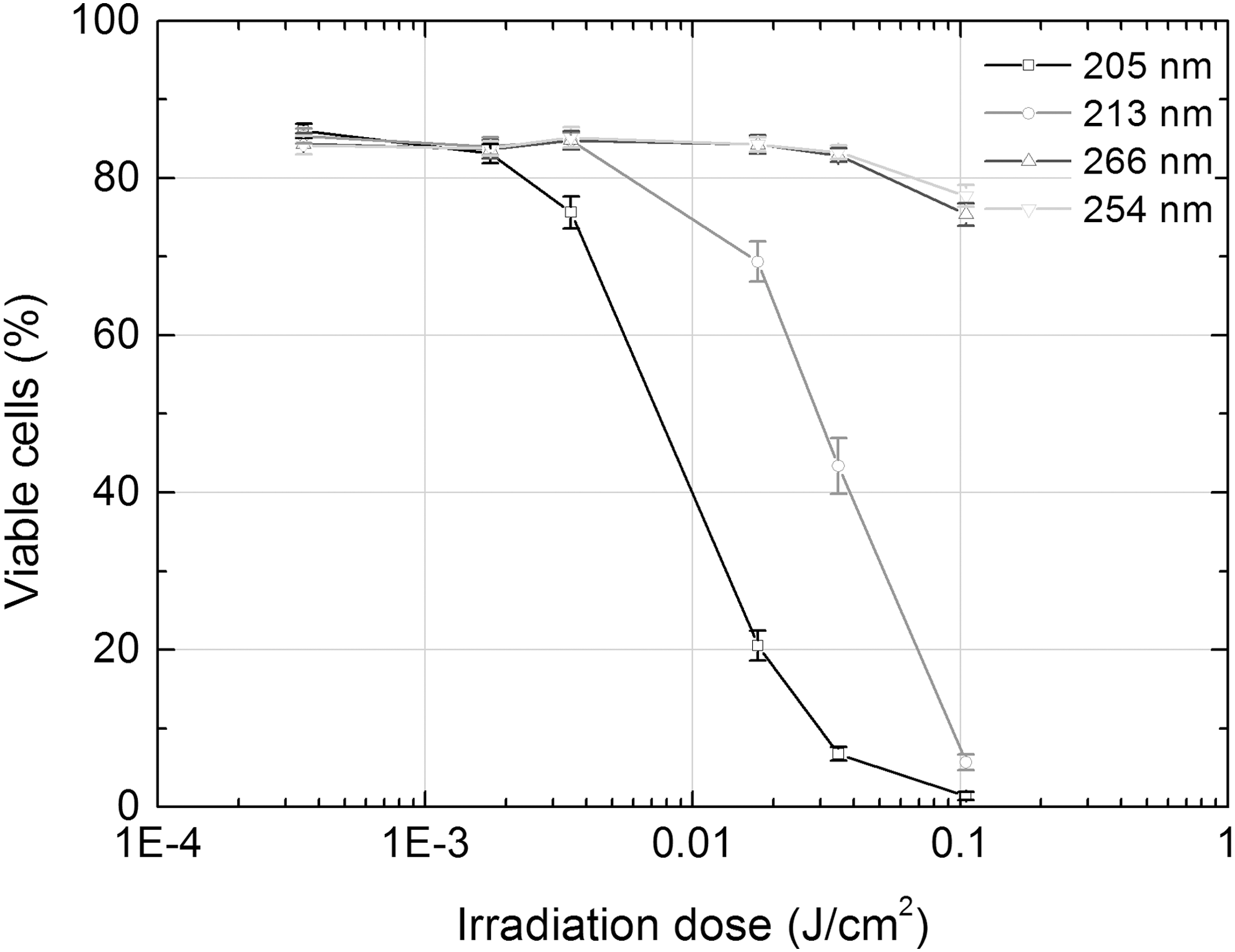
Viability of bone marrow cells exposed to UV irradiation of different parameters. The viability of cells was assessed by the trypan blue exclusion immediately after irradiation of cells' suspension by UV.
p < 0.0001, b p < 0.0005, c p < 0.05 when compared with unirradiated control.
Irradiation of shorter wavelengths (205 and 213 nm vs. 254 and 266 nm) had evidently stronger cytotoxic effect and obvious dosage-effect dependence (R 2 = 0.63 for 205 nm and R 2 = 0.96 for 213 nm irradiation). The 205 nm femtosecond irradiation was most cytotoxic: its last three doses decreased the viability of cells by 4.2, 12.7, and 60.8 times, respectively, as compared with control, whereas the same three doses of 213 nm nanosecond irradiation did it by 1.2, 2, and 14.9 times, respectively. Four doses of irradiation that did not decrease viability of cells >20% (except the fourth 205 nm irradiation's dose) were then used for the assessment of genotoxicity of different parameters' UV irradiation by comet assay. The results of the assessment are presented in Table 3 and Fig. 3.
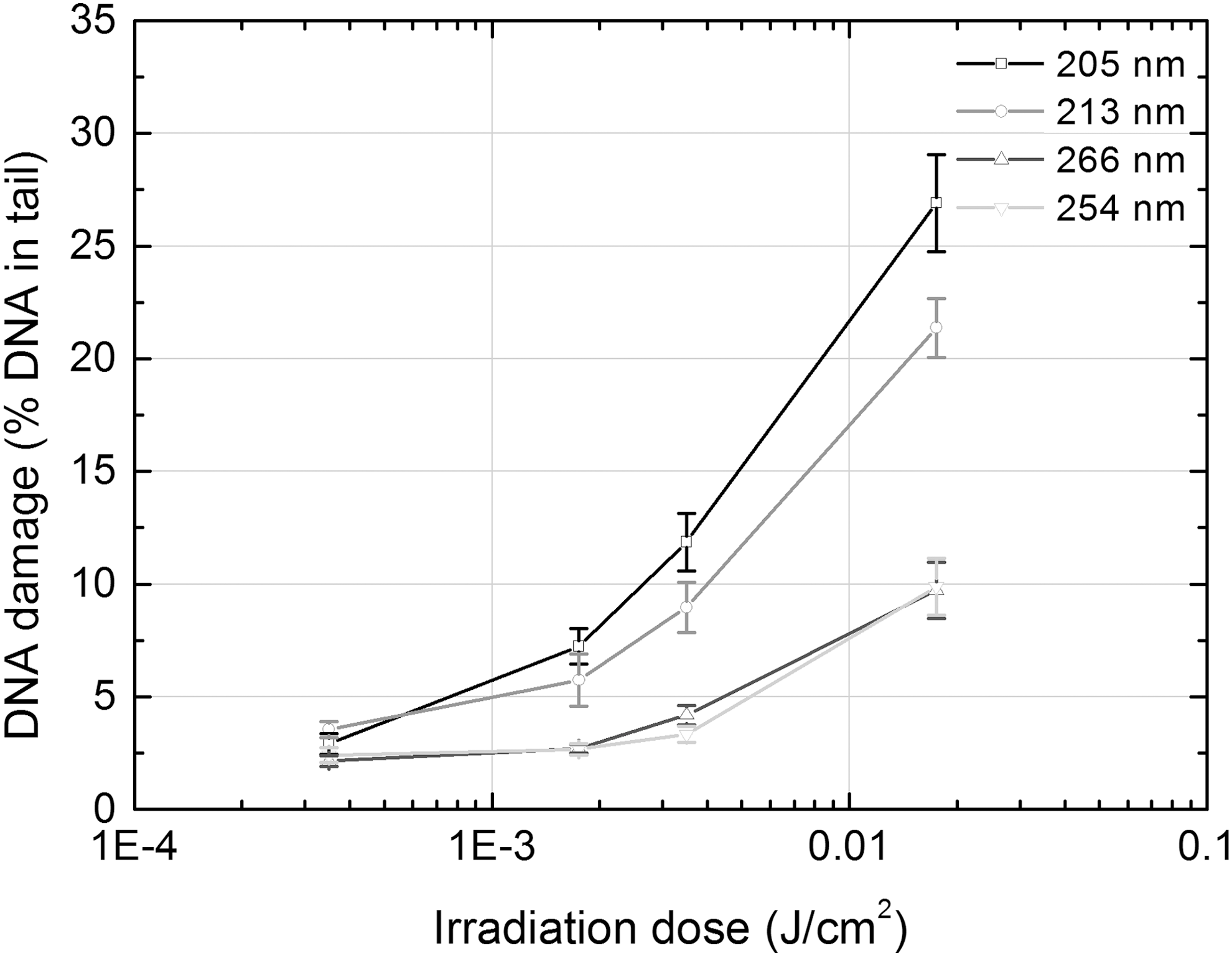
The DNA-damaging effect of UV irradiation of different parameters, assessed by DNA content in the comet tail in bone marrow cells. The cells were embedded on comet slides and comet assay was performed immediately after irradiation of cells' suspension by UV.
p < 0.0001, b p < 0.005, c p < 0.05 when compared with unirradiated control.
As data in the Table 3 show, all doses of different parameters' UV irradiation except the lowest one (0.00035 J/cm2) caused a significant increase in the amount of DNA in the comet tail. All types of irradiation demonstrated strong or very strong dosage-effect dependence: R 2 ranged from 0.89 for 213 nm nanosecond irradiation to 0.99 for 266 nm nanosecond and 254 nm UV lamp irradiation (Fig. 3). It should be noted that the damaging effect of irradiation on DNA was mainly dependent on wavelength rather than on other parameters. Shorter wavelength irradiation (205 and 213 nm vs. 254 and 266 nm) showed a 2–3.5 times stronger genotoxic impact. Moreover, comparable wavelength irradiation that greatly differed in pulse duration and repetition rate (205 vs. 213 nm), and even in the mode of beam energy delivery to a target – that is, continuous or pulsed energy (254 vs. 266 nm) – induced an almost identical amount of DNA breakage in cells exposed at the same irradiation dose.
Discussion
Our study of the possible genotoxic impact of femtosecond and nanosecond UV laser pulses on the DNA of murine bone marrow cells in vitro revealed a similar damaging effect of both types of radiation. Genotoxicity (and cytotoxicity) of investigated UV radiation was obviously dependent on wavelength rather than on other parameters such as pulse duration, repetition rate, or even the mode of beam energy delivery to a target.
It was unexpected that radiation of both shorter wavelengths (205 and 213 nm) induced a considerably greater amount of DNA damage than 254 and 266 nm radiation, which are known as particularly genotoxic, and were used in the current study for comparison of results. There may be a few reasons for that. The most important is the comparatively small size of murine bone marrow cells (on the average 7.6 μm in our samples), and the consequently weak filtering effect of a thin layer of cytoplasm surrounding the cell's nucleus. 37 Even 205 and 213 nm radiation penetrated through the thin cytoplasmic layer, reached the nuclear DNA, and damaged genetic material to a larger extent than radiation from both longer wavelengths, because of the greater energy of photons in the shorter wavelength range. Another reason could be the possible effect of secondary radiation, which can be particularly significant in the range of far-UV radiation. 38 As the absorbance by suspension of shorter wavelength light was three- to fourfold higher than the absorbance of 254 and 266 nm light (Fig. 1), it is likely that the secondary radiation, emitted during and after the treatment of bone marrow cell suspension, enhanced the genotoxic effect of irradiation at shorter wavelengths. A further possible explanation of the comparatively high cytotoxicity and genotoxicity of 205 and 213 nm radiation can be found in the established fact that the absorption by molecular oxygen of light <240 nm initiates a cascade of photochemical reactions, which produces a mix of strong oxidants, collectively referred to as reactive oxygen species (ROS), in air and in aqueous phases. 39 –42 It was demonstrated that ROS have the potential to interact with cellular components including DNA bases or the deoxyribosyl backbone of DNA, producing abnormal base adducts or strand breaks, 43 –45 and thereby increasing the damaging effect of short-wave UV radiation. Finally, strand breaks measured by comet assay are a relatively minor form of DNA damage. 34 Other kinds of damage, including cyclobutane pyrimidine dimers, base oxidation, and DNA-protein cross-links, which are also induced by UV exposure, 46,47 were not assessed in this study. However, as some previous studies have demonstrated, single strand breaks are generated in all UV ranges, and these lesions tend to be even more frequent than pyrimidine dimers in the longer-wave UV region. 48,49 These findings enable the use of strand breaks as a fairly unbiased indicator for the assessment of the damaging effect of UV radiation on DNA.
The objective of this study was to assess and to compare the general genotoxic potential of UV laser irradiation of different parameters, rather than discussing possible mechanisms or pathways of the damaging effect of the irradiation on DNA. There are some reasons, however, why such assessment can be incorrect when an in vitro model is used. First, unequal absorbance of light by aqueous solution can cause differences in the damaging effect of irradiation. A wavelength not significantly affected by aqueous solution, for example, may provide more reliable and predictable treatment outcomes. 50,51 Second, because the medium used for in vitro experiments usually contains a certain part of water, higher amounts of ROS can be generated and photogenotoxic effect can be overestimated. 52 As is apparent from Fig. 1, the absorption by DPBS of radiation in our investigated range (205–266 nm) was weak and almost constant; therefore, outcomes of different wavelength irradiations could not be biased. Further, it must be noted that before and during surgical procedures in vivo when lasers are applied, different aqueous solutions in organs or tissues are instilled in the area of laser treatment to achieve a better quality of treatment and greater comfort for the patient. For example, in an in vivo rabbit model of corneal laser surgery, three drops of different aqueous solutions in the eye were instilled. 16 When irradiated, these administered solutions can be an additional source of ROS, which can enhance the photogenotoxic effect in vivo as well. Therefore, both the direct and indirect damaging impacts of irradiation on DNA must be measured for complete assessment of the genotoxic potential of the lasers used currently or considered for future medical procedures.
The results of the current study demonstrated an intensity-dependent genotoxic impact of both 205 nm femtosecond and 213 nm nanosecond laser pulses on the DNA of bone marrow cells. It must be noted however, that primary DNA damage resulting from laser irradiation does not necessarily lead to genetic disorders. Risk of mutations and, therefore, malignancy is greatest in cases in which laser irradiation is less cytotoxic but induces more primary damage to genetic material. 19 Therefore, the resulting mutagenicity can be viewed as a combined effect of the absence of cytotoxicity plus DNA damage. To roughly quantify this effect, one would have to multiply the estimation of genotoxicity (Table 3) by the relative amount of viable cells remaining (Table 2). The resulting numbers are provided in Table 3, in parentheses next to the data for primary DNA damage. Such estimates, although very approximate, suggest the mutagenic potential of different irradiations. From the numbers provided, one can conclude that a combined effect of cytotoxicity plus genotoxicity is uncorrelated with the genotoxic effect taken separately. In our experiment, 205 nm femtosecond pulses induced a comparable amount of DNA damage in bone marrow cells as irradiation with 213 nm nanosecond pulses of the same intensity, but showed considerably stronger cytotoxic impact. This feature could be considered as one of the advantages of 205 nm femtosecond laser pulses when applying UV radiation for medical use as a result of reducing the risk of the formation of mutations and tumors in irradiated tissues or organs.
Conclusions
In the current study, both 205 nm femtosecond and 213 nm nanosecond laser pulses showed comparable intensity-dependent genotoxic impact by inducing strand breaks in the DNA of murine bone marrow cells in vitro. The damaging effect on the DNA was obviously dependent on the wavelength rather than on other parameters of irradiation. Investigations of possible genotoxic and mutagenic effects of UV femtosecond laser irradiation on cells of such tissues as the cornea and skin epithelium are needed for a better assessment of the risk associated with the clinical use of lasers.
Footnotes
Acknowledgments
This work was supported by European Community's social foundation under Grant Agreement No. VP1-3.1-ŠMM-08-K-01-004/KS-120000-1756.
Author Disclosure Statement
Ms. Gabryte and Mr. Danielius are paid employees of Light Conversion Ltd. Mr. Danielius is a shareholder of Light Conversion Ltd. None of the other authors has a financial or proprietary interest in any material or method mentioned.