
Abstract
Introduction
D
Although there are a number of antifungal agents available for topical and systemic treatment of dermatophyte infections, nail infections are particularly difficult to treat, with recurrence reported in up to 25–40% of cases. 5 It is currently unknown if the fungal recurrence is the result of inefficient clearance of the infection, or re-emergence of disease; at present, terbinafine is considered the most powerful treatment. 6
An alternative treatment strategy for dermatophyte infections is the use of photodynamic antimicrobial chemotherapy (PACT), which involves the use of photosensitizer chemicals and irradiation with specific wavelengths of light. Smij and Schuitmaker 7 demonstrated the inactivation of T. rubrum using the photosensitizers 5,10,15-tris(4-methylpyridinium)-20-phenyl-[21H,23H]-porphine trichloride (Sylsens B) and deuteroporphyrin monomethyl ester (DP mme) in conjunction with broadband white light irradiation. More recently, Rodrigues et al. 4 demonstrated successful PACT inactivation of both T. mentagrophytes and T. rubrum microconidia using novel phenothiazinium photosensitizers and red light.
Although the PACT approach requires the use of both photosensitive chemicals and light, it has also been found possible to photoinactivate a wide range of microorganisms using violet-blue light from the visible spectrum, without the use of exogenous photosensitizers, with comparative doses being safe for mammalian cell exposure. 8 –13 Microbial inactivation by violet-blue light is accredited to the photoexcitation of intracellular porphyrin molecules within microorganisms, which have an absorption maxima in the region of 400 nm, 14 which causes the production of reactive oxygen species (ROS). 15,16 Cell death has been attributed to oxidative damage to cell components, including DNA and membranes. 9,12 It has previously been established that 405 nm light has antifungal effects, as Murdoch et al. 17 demonstrated the inactivation of the fungal species Saccharomyces cerevisiae, Candida albicans, and dormant and germinating spores of Aspergillus niger.
The current study highlights the fungicidal activity of 405 nm violet-blue light against the dermatophytes T. rubrum and T. mentagrophytes, and the high sensitivity of these fungi to 405 nm light was compared against the saprophyte A. niger, with results opening up the possibility of the development of 405 nm light treatments against dermatophytic infections.
Materials and Methods
Fungal strains and conidia preparation
The dermatophytic fungi used in this study were T. rubrum MUCL 11954 and T. mentagrophytes MUCL 9823, obtained from the Mycotheque de l'Universite catholique de Louvain Culture Collection in Belgium. The saprophytic mould fungus A. niger MUCL 38993 was also used in comparative light sensitivity studies with the two dermatophytic species.
T. rubrum and T. mentagrophytes spores were obtained by fungal cultivation on sabauroud dextrose agar (SDA) plates (Oxoid, UK) at 28°C for 14 days. Following incubation, 9 mL phosphate buffered saline (PBS) (Oxoid Ltd, UK) containing 0.01% Tween 80 was added to the dish, and an L-shaped spreader used to agitate and release the microconidia. Agitation was performed for 2 min. The resulting suspension was stored at 4°C.
To obtain A. niger spores, A. niger was inoculated onto a SDA slope and incubated at 26°C for a minimum of 7 days, after which a conidial suspension was obtained by agitation in an aqueous 0.01% Tween 80 PBS solution. Agitation was performed for 5 min. The population density of the spore suspensions was enumerated using an Improved Neubauer haemocytometer (Weber Scientific International, UK), and suspensions were diluted as required prior to light exposure.
Light transmission through conidial extracts
For light transmission tests on conidial extracts, the centrifuged pellets of conidia of A. niger and T. rubrum were extracted with 100% ethanol, and the light transmission spectrum of the ethanol extracts was determined using a Biomate 5 UV-Visible Spectrophotometer (Thermo Spectronic).
Light source and irradiance measurements
An indium-gallium-nitride 99-DIE light-emitting diode (LED) array (OptoDiode Corp, CA) was used to generate high-intensity 405 nm light with a bandwidth of 14 nm. The LED array was powered by a DC power supply, and a cooling fan and heat sink were attached to the array, allowing heat to dissipate from the source, thereby minimizing heat transfer to the fungal samples. Irradiance was measured using a radiant power meter and detector (L.O.T.-Oriel ltd, UK). The dose of light exposure (J/cm2) was calculated as the product of the irradiance (mW/cm2) multiplied by the exposure time (seconds). Doses selected for use in this study were between 500 and 1500 J/cm2, as these were within the region of those used in previous fungal inactivation studies. 16
Light exposure of Trichophyton and Aspergillus conidia
The inhibitory effects of 405 nm light on conidia were assessed using surface-irradiated and liquid-irradiated exposure conditions. For surface irradiation tests, 10 μL conidial suspension of the test fungus was spot inoculated onto the center of an SDA plate. The test plate was exposed to 405 nm light, at an irradiance of 35 mW/cm2 for 1 and 4 h, giving doses of 126 and 504 J/cm2. Identical control samples were prepared and left exposed to normal laboratory lighting. Plates were incubated for 3 or 10 days for Aspergillus and Trichophyton, respectively, before being analyzed for characteristic differences between the test and control. Colony diameters were measured across the broadest section of the colony on the SDA plate using a ruler. The results were also recorded photographically for illustrative purposes using a Sony Cybershot DSC-T2 digital camera (Sony, Japan).
For liquid irradiation comparisons, a 3 mL volume of spore suspension of test fungi was transferred into the well of a 12-well multidish with the LED housing array then placed ∼3 cm above. The suspension was exposed to 50 Mw/cm2 405 nm light, for 2, 4, and 8 h, giving doses of 360 J/cm2, 720 J/cm2, and 1.44 kJ/cm2. Control samples were held under the same conditions, but exposed to normal laboratory lighting.
Light exposure of A. niger hyphal suspension
Following 24 h incubation of A. niger on an SDA slope, the top layer of fungal growth was removed and then fragmented in 50 mL PBS for 5 min using a stomacher (Colworth, UK), and 3 mL of the hyphal suspension was pipetted into one well of a 12-well multidish and exposed to 405 nm light, as described, for conidia suspension tests.
Plating and enumeration
For suspension experiments, post-exposure, samples (50, 100, or 500μL) were inoculated onto SDA and spread using an L-shaped spreader. Plates were then incubated for 1 or 5 days (A. niger and T. rubrum, respectively), with each sample being plated at least in triplicate. Following incubation, the plates were enumerated and recorded as colony-forming units per milliliter (CFU/mL). Data presented in this article represent the mean results of two or more independent experiments. Significant differences in fungal population were calculated at the 95% confidence interval (p < 0.05) using one-way analysis of variance (ANOVA), with Minitab statistical software package version 16 (Minitab Inc., PA).
Results
The inhibitory effects of 405 nm light on the growth of surface irradiated T. rubrum and T. mentagrophytes spores are shown in Fig. 1. The results demonstrate that after seeding the conidia onto SDA plates, and exposure to 126 J/cm2, followed by incubation for 10 days, a substantial reduction in growth was observed with T. rubrum, with the diameters of the non-light-exposed colonies and light-exposed colonies measuring 43 ± 1 and 12 ± 1 mm in diameter, respectively (Fig. 1 A,B). Following exposure to a dose of 504 J/cm2, both T. rubrum and T. mentagrophytes were completely inactivated, and failed to develop colonies, with the unexposed controls developing colonies of 21–22 mm diameter (Fig. 1 C–F). Exposure of surface deposited conidia of A. niger to the same dose of 504 J/cm2, followed by incubation for 3 days, demonstrated that complete inactivation of the spores was not achieved, with substantial conidial growth observed following incubation: light-exposed colonies grew to 38 ± 1 mm diameter, compared with 33 ± 1 mm for unexposed colonies (∼13% reduction in size; p = 0.049).

Inhibitory effects of 405 nm light on the growth of Trichophyton rubrum (
Suspensions of T. rubrum conidia, A. niger hyphal fragments, and A. niger conidia were exposed to 405 nm light at an irradiance of 50 mW/cm2 over time periods that delivered a dose of 360 and 720 J/cm2 and 1.44 kJ/cm2. Following exposure to a dose of 360 J/cm2, complete inactivation of T. rubrum conidia was achieved (∼2.3-log10 CFU/mL). The results in Fig. 2 demonstrate that A. niger hyphae are more sensitive to 405 nm light than their corresponding conidia, with complete inactivation of a 103 CFU/mL hyphal suspension found after exposure to a dose of 1.44 kJ/cm2, whereas A. niger conidia demonstrated an ∼50% reduction following exposure to the same dose of 1.44 kJ/cm2 (Fig. 2.). Use of an increased irradiance or longer exposure time (meaning an increased applied dose) would lead to further decreases in the A. niger population, as previously reported. 16

Exposure of Trichophyton rubrum and Aspergillus niger conidial suspensions to 405 nm light using an irradiance of 50 mW/cm2 to deliver dose levels of 360, 720, and 1.44 kJ/cm2. Inactivation of A. niger hyphal fragments was included as a comparison. Surviving fungi were enumerated by mean colony-forming units per milliliter (CFU/mL) counts (± SD), and results were reported as the % log10 reduction compared with unexposed control samples. Asterisks (*) represent where a significant difference was detected between the exposed and unexposed samples, at 95% confidence level (p ≤ 0.05).
The light transmission through ethanol extracts of A. niger and T. rubrum spores was measured to determine the effect of spore pigments on the transmission of 405 nm light through suspensions of both A. niger and T. rubrum; in Fig. 3, the transmission spectra are shown alongside the emission spectrum of the 405 nm LED.
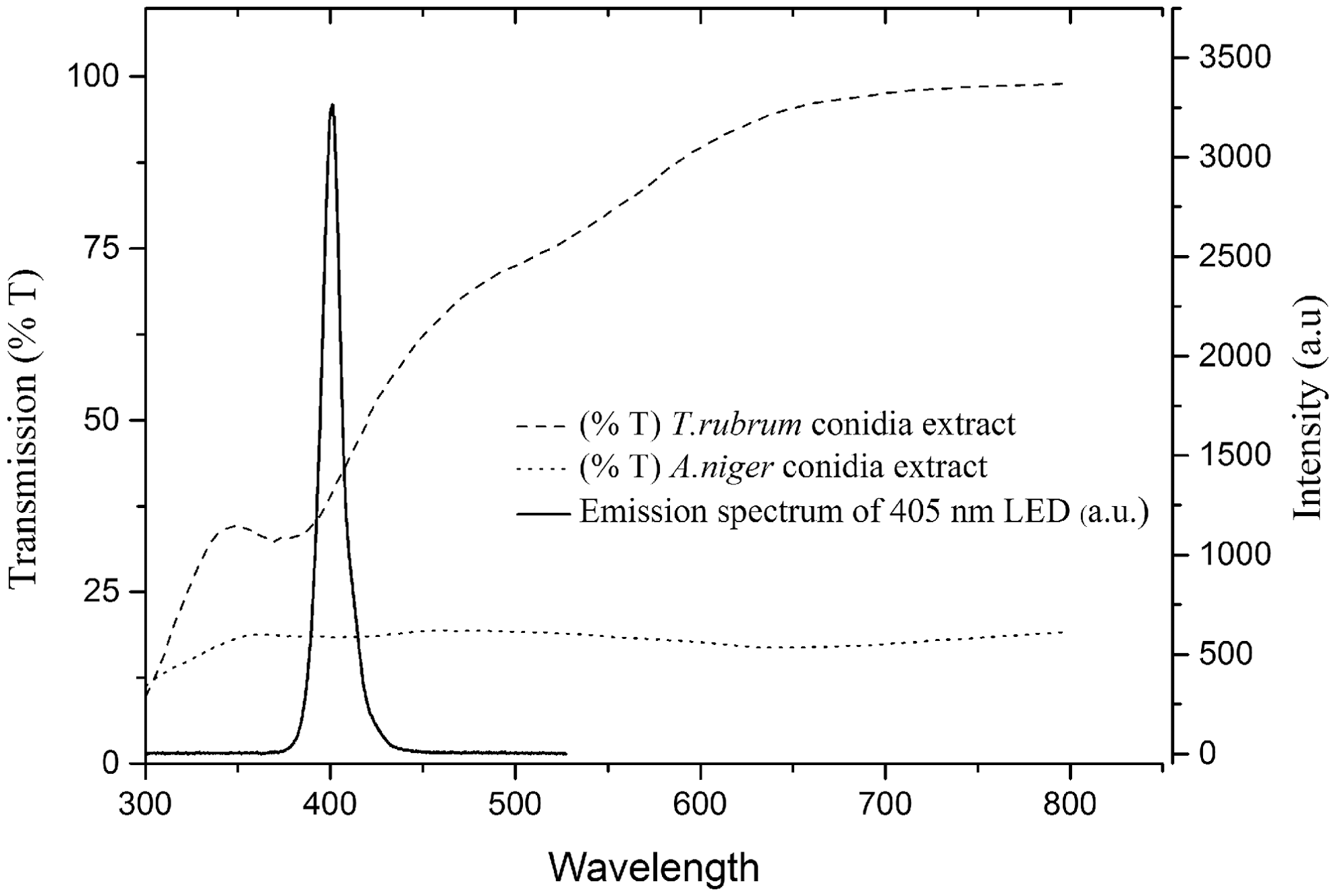
The transmission of light, over the wavelength range 300–800 nm, through ethanol extracts of the conidia of Trichophyton rubrum and Aspergillus niger. The emission spectra of the 405 nm light-emitting diode (LED) array, measured using a high resolution spectrometer (Ocean Optics Inc, USA), is included for reference.
Discussion
The results shown in Fig. 1 demonstrate that a substantial reduction in microconidial growth of T. rubrum is achieved following exposure to 405 nm light at a dose of 126 J/cm2. Further, the results demonstrate that 405 nm light, at a dose of 504 J/cm2, can completely inactivate the microconidia of T. rubrum and T. mentagrophytes such that hyphal and colony growth do not occur. By contrast, exposure of surface deposited conidia of A. niger to a similar dose of 504 J/cm2 did not result in a substantial reduction in conidial growth; therefore, on subsequent incubation of the SDA plates colony growth occurred and the colony diameter achieved after 3 days of incubation was only marginally less than that observed with the control unirradiated plates (Fig. 1). Even after an increased light dose of 1.008 kJ/cm2, A. niger conidia were not completely inactivated, and colony growth occurred, although the extent of growth was considerably less than that of the unexposed control: colony diameter of 22 mm for light-exposed and 39 mm for control (not shown). These results demonstrate the higher susceptibility of the dermatophytic conidia of both T. rubrum and T. mentagrophytes to inactivation using 405 nm light, compared with A. niger conidia.
It is known that exposure of microbiological culture media to light can result in the formation of toxic compounds. 18 To ensure that the results obtained were caused by direct light-induced inactivation of the fungal conidia as opposed to an indirect media-induced toxic effect, experiments were also conducted using liquid suspensions of the fungal conidia and hyphae. Comparison of the susceptibility of T. rubrum conidia, the conidia and hyphae of A. niger, to 405 nm light at doses of 360 and 720 J/cm2 and 1.44 kJ/cm2, demonstrated the much higher susceptibility of T. rubrum to inactivation using 405 nm light than that of A. niger, with complete inactivation achieved at 360 J/cm2 (Fig. 2). The conidia of A. niger were much more resistant to 405 nm light, and although the CFU count decreased with increasing dose, complete inactivation was not achieved with the doses used in the present study. As we reported in a previous study. 11 complete inactivation of A. niger conidia, with higher populations of 105 CFU/mL, required a dose of 2.3 kJ/cm2. Although conidia of A. niger are highly resistant to 405 nm light, it was of interest to compare the sensitivity of A. niger hyphae with that of the conidia. The results, shown in Fig. 2, demonstrate that A. niger hyphae are more sensitive to 405 nm light than their corresponding conidia, with complete inactivation of a 103 CFU/mL hyphal suspension found after exposure to a dose of 1.44 kJ/cm2. It was interesting to note, however, that the A. niger hyphae demonstrated more resistance to the 405 nm light than the T. rubrum conidia.
The mechanism of the antifungal effect mediated by violet-blue light occurs following exposure of the organism to light photons in the region of 405 nm. Endogenous porphyrins within the cells absorb these photons, resulting in their photoexcitation, and electron transfer via the type I or type II pathway, resulting in the production of ROS, most notably singlet oxygen (1O2). 15,19 The ROS produced then react with various cellular components, causing an imbalance in cellular homeostasis resulting in damage to cytoplasmic organelles and nucleic acids, and, consequently, cell death by apoptosis, necrosis, or autophagy. 20 This hypothesis is supported by a study by Baltazar et al., which demonstrated the photodynamic inactivation of T. rubrum, via increased levels of NO, ROS, and ONOO, using 630 nm light and the exogenous photosensitizer toluidine blue. 19
Fungi possess mitochondria and, although there are some enzyme differences when compared with mitochondria of mammalian cells, the production of the endogenous photosensitive protoporphyrin IX molecule has been demonstrated. 21 –23 Protoporphyrin IX may be activated by wavelengths ranging from UVA to the visible wavebands, with a maximum peak in the Soret band at 375–405 nm, and a lower peak at 630–633 nm. 21 –23 The presence of porphyrins in fungi indicates that both bacteria and fungi may be affected by a similar porphyrin photoexcitation and ROS-induced inactivation mechanism following exposure to visible light. 13,17 Further evidence that a similar underlying inactivation mechanism is involved is the finding 17 that light exposure under aerobic and anaerobic conditions, together with results obtained using oxygen scavengers, has revealed that 405 nm light inactivation in fungi involves an oxygen-dependent mechanism, which is also the case with bacteria. Although the inactivation mechanism may be similar, the physiological status of the organism is an important factor influencing the degree of susceptibility of the light-exposed cells, with bacterial and fungal spores being understandably more resistant than their vegetative counterparts, an innate resistance that rapidly disappears during spore germination. 17
Although most previous research on the use of light to inactivate fungi has involved the use of added photosensitizer chemicals, a previous study by Smijs et al. 21 demonstrated the ability of UVA light alone, at a dose of 40 J/cm2, to kill T. rubrum without the use of exogenous photosensitizers. In addition to this, irradiation with broadband visible light at a dose of 20–50 J/cm2 in the absence of exogenous sensitizers was found to produce oxygen-dependent lethal effects on the plasma membranes and mitochondria of Candida guilliermondii. 24 Within bacterial cells, porphyrin-mediated violet-blue light inactivation has been associated with severe cell wall damage and leakage of intracellular substances, presence of cytoplasmic vacuoles, and disruption of intracellular structures. 12,13
It is believed that pigments such as melanin, which are black or dark brown pigments, commonly occurring as wall components in fungal spores, have a protective role against photochemical damage. 25 To investigate the effects of spore pigment on 405 nm light transmission, ethanol extracts of A. niger and T. rubrum spore suspensions were prepared and the wavelength transmission spectra were compared (Fig. 3). Results demonstrate that the transmission of light across the measured spectrum (300–800 nm) is much lower for the A. niger than for the T. rubrum spore extract, with 18.4% and 41.3% transmission at 405 nm, respectively. The high resistance of A. niger spores to 405 nm light is most likely the result of possession of a multilayered pigmented spore coat containing aspergillin, a black melanin-like compound making the spores particularly difficult to inactivate when exposed to visible light, 26 and pulsed UV light. 27,28 The presence of the aspergillin pigment explains why A. niger conidia are more resistant to 405 nm radiation than the conidia of T. rubrum, and indeed to the A. niger hyphae which are unpigmented. Although T. rubrum also produces several melanin-type pigments, 29 these either do not occur in the conidia, or, at least, do not occur at sufficient levels to provide protection against 405 nm light irradiation.
Conclusions
The results of this study demonstrate that the microconidia of the Trichophyton spp tested are much more sensitive to inactivation by 405 nm light than the conidia of the saprophytic fungus A. niger. Although the resistance of A. niger conidia to light inactivation is not surprising given the dark pigment present, it is of interest that the Trichophyton microconidia were more sensitive to 405 nm light than the unpigmented hyphae of A. niger. Although Trichophyton microconidia are not the main transmissible agents of these dermatophytic fungi, they are regarded as the preferred fungal structure for dermatophytic antifungal susceptibility testing. 3,4 The findings of this study, demonstrating the relatively high sensitivity of Trichophyton microconidia to 405 nm light, are, therefore, of potential interest regarding the control and treatment of dermatophyte infections.
Footnotes
Acknowledgments
The authors thank the University of Strathclyde and The Robertson Trust for their support. The authors also thank Daniel Irving for technical support.
Author Disclosure Statement
No competing financial interests exist.