
Abstract
Introduction
I
Usually, PBM uses low-powered laser light in the range of mW, at wavelengths from 600 up to 1000 nm, and at a low fluence around 1–5 J/cm2 or less, 2,11,12 to stimulate a biological response. PBM works on the principle that when light hits certain molecules called chromophores, the photon energy causes electrons to be excited and to jump from low-energy orbits to higher-energy orbits. PBM is not an ablative or thermal mechanism, but it is rather a photochemical effect comparable to photosynthesis in plants whereby the light is absorbed and exerts a chemical change. 2
Scrupulous researchers support the idea that, in animal cells, PBM stimulates components in the mitochondria, 13 increasing redox capacity, protein electrochemical potential, 14 oxygen consumption, phosphate potential, and energy charge, 15 stimulating the proton pumping activity, 16 or activating cytochrome c oxidase (complex IV). 17,18 Moreover, increased ATP production in both irradiated normal and dysfunctional cells has been proved. 14,19
Recently, a variety of diseases have definitively been demonstrated to depend on mitochondria functions: This then makes it possible that PBM with mitochondria can affect their pathophysiological function, and may perhaps be useful in therapy. 13
However, despite many reports of positive findings from experiments conducted in vitro, in animal models and in randomized controlled clinical trials, PBM remains controversial because of conflicting effects by a large number of operating parameters such as wavelengths, fluences, or laser powers. 2,20 –22
Authors report how in laser therapy, the PBM contradictory results (no-effect, photobiostimulation, or photobioinhibition) are the consequences of an existing biphasic dose response. A “biphasic curve,” similar to the Arndt-Schulz curve described by Sommer et al., 23 can be used to illustrate the expected dose response to light at a subcellular, cellular, tissue, or clinical level. Simply put, it suggests that if insufficient energy is applied, there will be no response (because the minimum threshold has not been met). If more energy is applied, then a threshold is crossed and biostimulation is achieved; however, when too much energy is applied, the stimulation disappears and is replaced by bioinhibition instead. 2 However, if significant effort has been made to clarify parameters of deposited energy density that will effectively promote positive change in individual cells without negative effects, there are evident limits to pass from cellular to organismic level.
Actually, laser energy density and distribution at the tissue surface are poor predictors of deeper tissue distribution. Several problems complicate the adoption of a standardized protocol. The biostimulatory effect of laser energy is experienced at a cellular level, whereas the energy is applied macroscopically to large volumes of tissue in a non-uniform manner. As energy passes through tissue, part of it is absorbed, so each successive depth of cells is irradiated differently. Beer's law is usually used to define this relationship. This is inadequate, since the dominant form of interaction at wavelengths between 600 and 1400 nm is scattering. 24 Thus, as energy enters tissue, its density decreases rapidly. The output of most clinical lasers is Gaussian in special profile. Therefore, cells directly in the center of the beam are irradiated at a very high fluence, whereas those on the periphery of the incident beam receive a very low dose. As a result, cells at the center of the beam may be overstimulated far above the scientifically recommended range of 3–10 J/cm2 and inhibited; whereas those on the periphery receive insufficient cellular energy to produce any effect.
A further complicating standardization is the issue of beam divergence. Fiber-delivered laser energy exits the fiber with a significant divergence, usually on the order of 8°. The applied energy is, therefore, distributed over an increasing area as the tip-to-tissue distance increases dramatically, affecting energy density at a cellular level. At currently reported beam divergences, energy density can be diminished by 90% with only 3 mm of tip-to-tissue distance. This makes the repeatable application of an appropriate energy density extremely technique sensitive and operator sensitive.
Recently, to have deeper tissue penetration, the irradiation procedures have been simplified; to generate a more homogenous irradiation, a modern flat-top handpiece has been developed, which enables to irradiate a 1 cm2 area with the same power density and allows laser therapy by high-fluence (64 J/cm2) and high-power [continuous wave (CW)] (1 W) (AB2799; Doctor Smile) (Figs. 1 and 2). Our preliminary results show that this higher-fluence and higher-power laser therapy induces a precocious fission rate in Paramecium primaurelia cells for 10 successive generations, 25 increases P. primaurelia oxygen consumption 26 and ATP production, 27 and affects Paramecium swimming behavior and food vacuole formation, 27 and in association with exercise therapy, higher fluences appear to be effective in the treatment of nonspecific chronic low back pain. 28 Recently, we showed that the parameter 64 J/cm2–1 W more than the 3 J/cm2–100 mW makes P. primaurelia mitochondria respiratory chain activity more efficient. 29
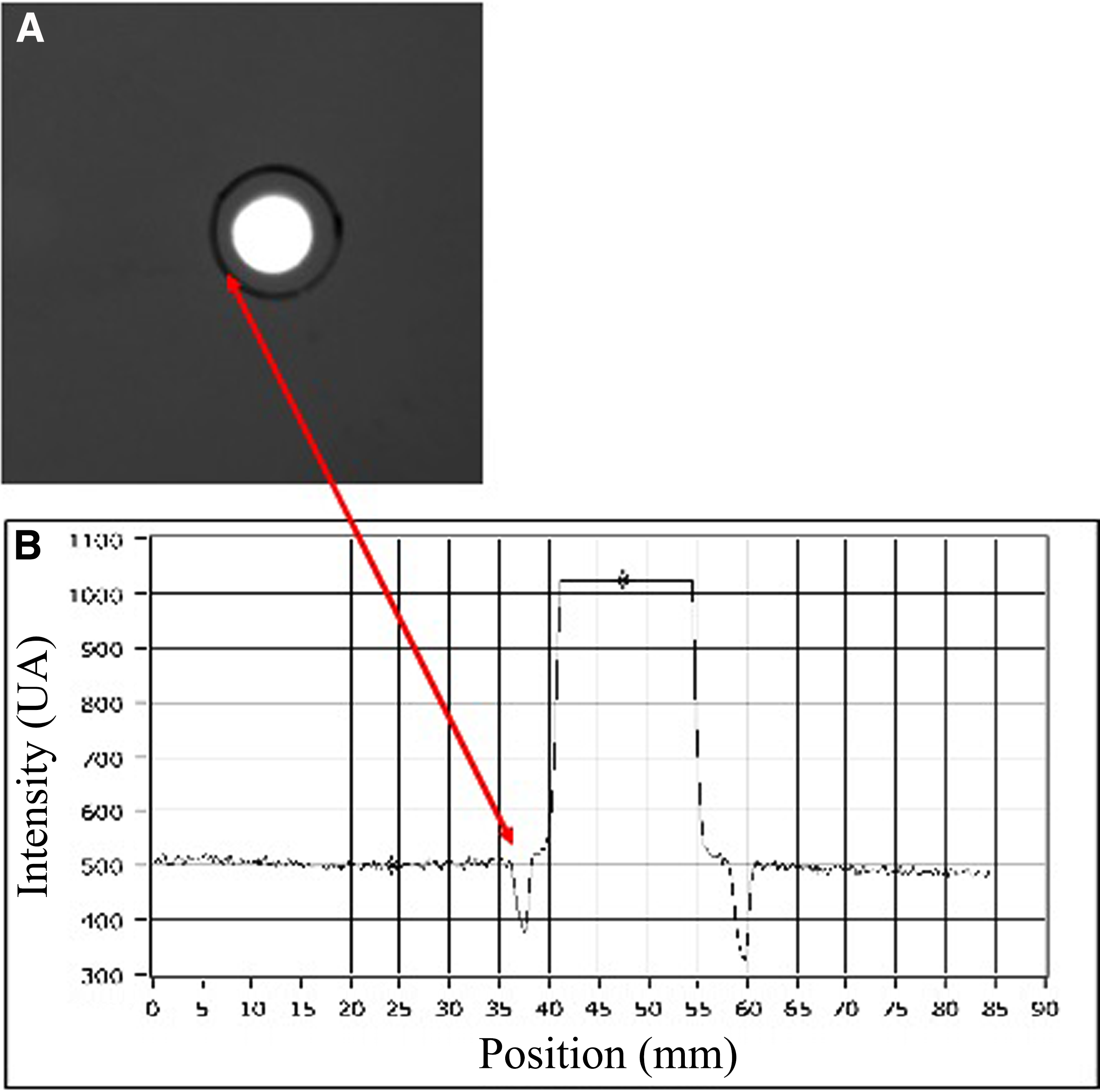
Flat-top profile of 808 nm diode laser. Laser beam profilers are devices that measure the energy level of each point of the cross-section of the laser beam and give a measure of it. This picture shows a flat-top distribution of the laser radiation [spot size
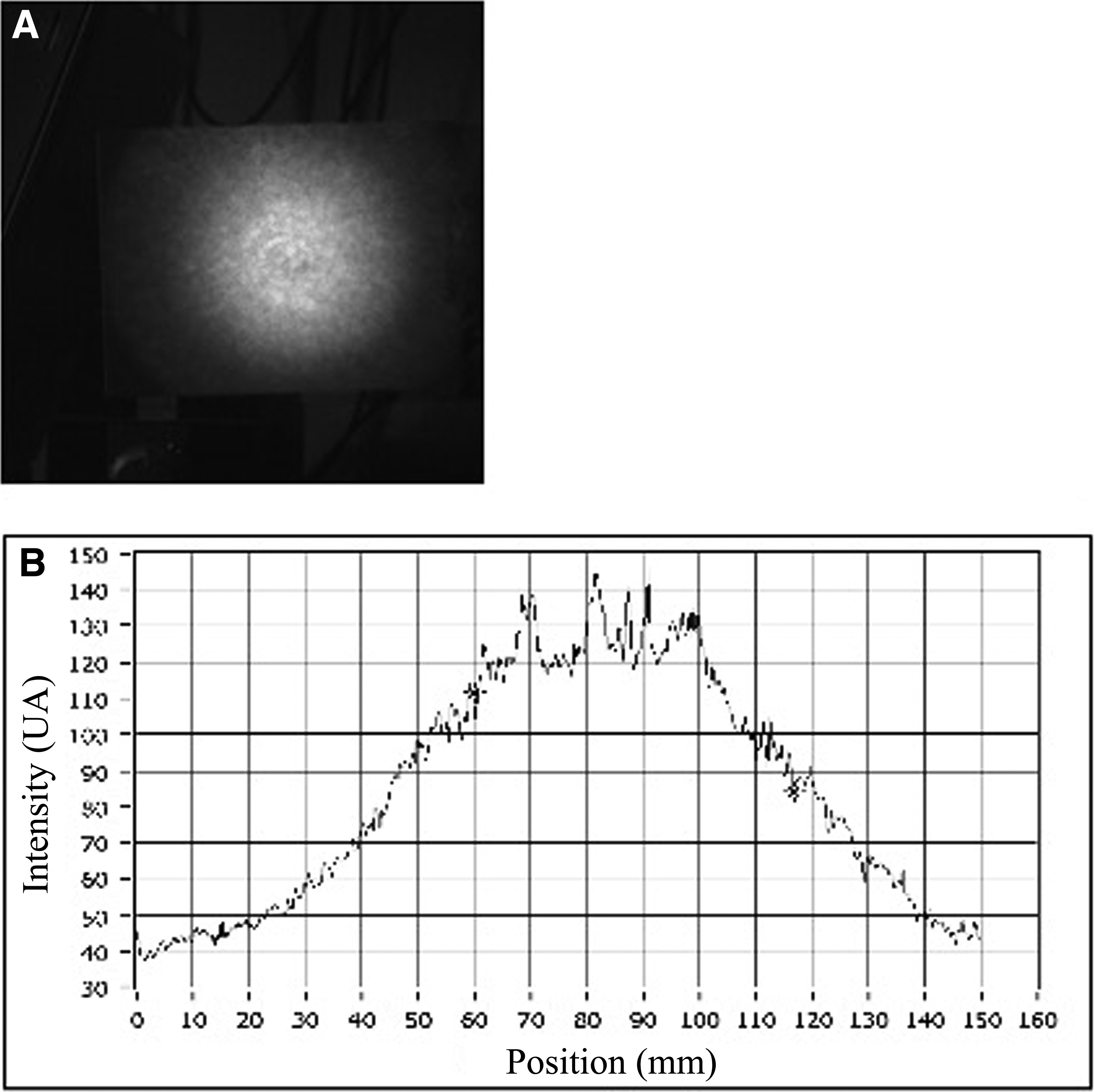
Wave profile of a conventional 808 nm defocused handpiece. The earlier picture refers to a circular laser beam
On this basis, to move from a unicellular eukaryotic organism model to a mammalian cellular model, in our work, we detected the photobiomodulatory effect of an 808-nm diode laser (1 W–64 J/cm2) irradiated by a flat-top handpiece on mammalian's mitochondria activities, such as oxygen consumption, activity of mitochondria complexes I, II, III, and IV, and cytochrome c as well as ATP synthesis.
Materials and Methods
Materials
Substrates and the other analytical-grade chemicals were purchased from Sigma–Aldrich. Ultrapure water (Milli-Q; Millipore) was used. Safety precautions were taken in carrying out the experiments posing chemical hazards. Ampicillin (100 μg/mL) was added to the solutions. Sterile experimental conditions were employed where needed.
Mitochondria-enriched fraction isolation
Mitochondria were purified from bovine liver. 30 –32 Briefly, liver washed in PBS was homogenized in a buffer, containing 0.25 M sucrose, 0.15 M KCl, 10 mM Tris–HCl pH 7.4, 1 mM EDTA, and 0.5% BSA. The homogenate was centrifuged at 800 g for 10 min. The supernatant was filtered and centrifuged at 12,000 g for 15 min. The pellet was resuspended in another buffer, containing 0.25 M sucrose, 75 mM mannitol, 10 mM Tris–HCl pH 7.4, and 1 mM EDTA. The final supernatant was centrifuged at 12,000 g for 15 min, and the mitochondrial pellet was resuspended in the second buffer.
Irradiation of mitochondria
The mitochondria samples were irradiated in Eppendorf tubes by an 808-nm infrared diode laser (50 sec, 64 J/cm2, 1 W) (Wiser; Doctor Smile) with a flat-top handpiece (1 cm2 of spot) (AB2799; Doctor Smile), whereas the non-irradiated mitochondria were considered as controls.
Oxygen consumption measurements
Oxygen (O2) consumption was assayed in a thermostatically controlled oxygraph apparatus equipped with an amperometric electrode (Unisense–Microrespiration). For each sample, 50 μg of mitochondria proteins was used, treated or not with laser. The samples were incubated in the respiration buffer composed of: 120 mM KCl, 2 mM MgCl2, 1 mM KH2PO4, 50 mM Tris–HCl, pH 7.4, and 25 μg/mL Ampicillin. To observe the respiration rates in coupled conditions, first, 5 mM pyruvate plus 2.5 mM malate was added, to stimulate the complexes I + III + IV pathway; whereas 20 mM succinate was added to stimulate the pathway composed by complexes II, III, and IV. To observe whether the respiration was coupled with the ATP synthesis, 0.2 mM ADP was added after the respiring substrates, to induce the respiration state 3. To verify that O2 consumption was dependent on the activity of the OXPHOS proteins, 40 μM rotenone (complex I inhibitor) or 50 μM antimycin A (complex III inhibitor) was added. 32
Spectrophotometric assay of respiring complex
The activity of the four electron transport chain (ETC) complexes was assayed on 50 μg of total mitochondria proteins, treated or not with laser. Complex I (NADH-ubiquinone oxidoreductase) was assayed after the reduction of ferricyanide at 420 nm. Complex II (Succinic dehydrogenase) activity was measured at 600 nm, in 2 mM EDTA, 0.2 mM ATP, 20 mM succinate, 0.5 mM cyanide, 80 μM dicloroindophenol (DCIP), 50 μM decylubiquinone, 40 μM antimycin A, 10 μM rotenone, and 10 mM phosphate buffer, pH 7.2. Complex III (Cyt c reductase) activity was measured after the reduction of oxidized Cytochrome c (Cyt c) at 550 nm. Complex IV (Cyt c oxidase) was assayed after the oxidation of ascorbate-reduced Cyt c at 550 nm. 32 To evaluate the effect of laser on Cyt c, the complex III assay was repeated in the presence of cytochrome c that was previously treated with laser on untreated mitochondria.
F0–F1 ATP synthase assay
ATP synthesis was evaluated on 50 μg of mitochondria proteins, treated or not with laser. Samples were incubated for 10 min at 37°C in: 100 mM Tris/HCl (pH 7.4), 100 mM KCl, 1 mM EGTA, 2.5 mM EDTA, 5 mM MgCl2, 0.2 mM di(adenosine-5′) penta-phosphate, 0.6 mM ouabain, ampicillin (25 μg/mL), and 5 mM KH2PO4. As respiring substrates, 5 mM pyruvate plus 2.5 mM malate or 20 mM succinate was added. After the incubation, to induce ATP synthesis, 0.1 mM ADP was added to the mixture. Sample ATP concentration was measured in a luminometer (Lumi-Scint; Bioscan) by the luciferin/luciferase chemiluminescent method with ATP standard solutions between 10−9 and 10−7 M for calibration. 31,32
Statistical analysis
The experiments were repeated 10 times, and the statistical analysis was performed using a two-way ANOVA followed by the Tukey–Kramer multi-comparison test (GraphPad InStat 3) to discriminate statistically significant results.
Results
Evaluation of ETC activity in mitochondria after photoirradiation
To evaluate the effect of the photoirradiation on the mitochondria ETC, the oxygen consumption was evaluated. Figure 3 shows the amperometric traces of mitochondria photoirradiated (black line) or not (gray line) in the presence of pyruvate/malate and succinate, to induce the pathway formed by complexes I + III + IV or complexes II + III + IV, respectively. As it is possible to observe, the irradiated mitochondria consume more oxygen compared with the control (p < 0.001), in both conditions. Moreover, it is important to note that the addition of rotenone or antimycin A, inhibitors of complexes I and III, induces a decrement in the respiration, demonstrating that the oxygen consumption is really due to the ETC. Considering that both the pathways increase the respiration rate after the photoirradiation, it is possible to presume that the photoirradiaton may have an effect on either complex III or complex IV.
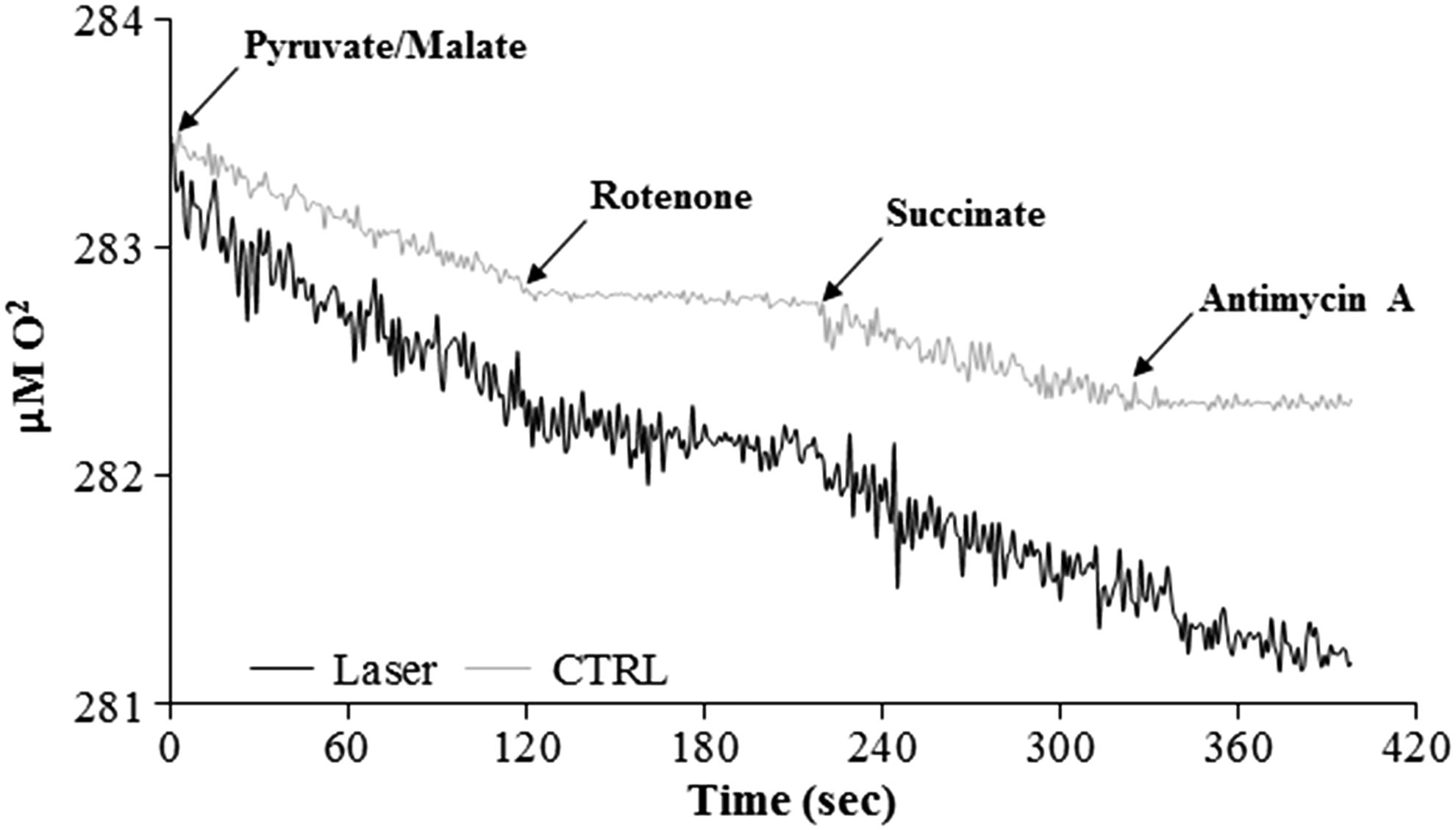
Oxygen consumption in photoirradiated mitochondria. The graph represents the amperometric trace of oxygen consumption in irradiated mitochondria (black line) and in not-treated mitochondria (gray line), used as a control. The respiration was measured in the presence of pyruvate/malate and succinate as respiring substrates and of rotenone and antimycin A, to inhibit complex I and complex III, respectively. The figure is representative of at least five measurements.
For this reason, the activities of each respiring complex were evaluated. In Fig. 4, it is possible to observe that complex I (A) and complex II (B) are not influenced (p > 0.05) by the photoirradiation; by contrast, both complex III (C) and complex IV (D) show an increment in the activity after the irradiation. In particular, complex III activity increases by about 33% (p < 0.001), whereas complex IV activity increases by about 70% (p < 0.001). This could suggest that the laser effect is directed on the cytochromes that compose the two respiring complexes or on cytochrome c, which transports the electrons from complex III to complex IV.
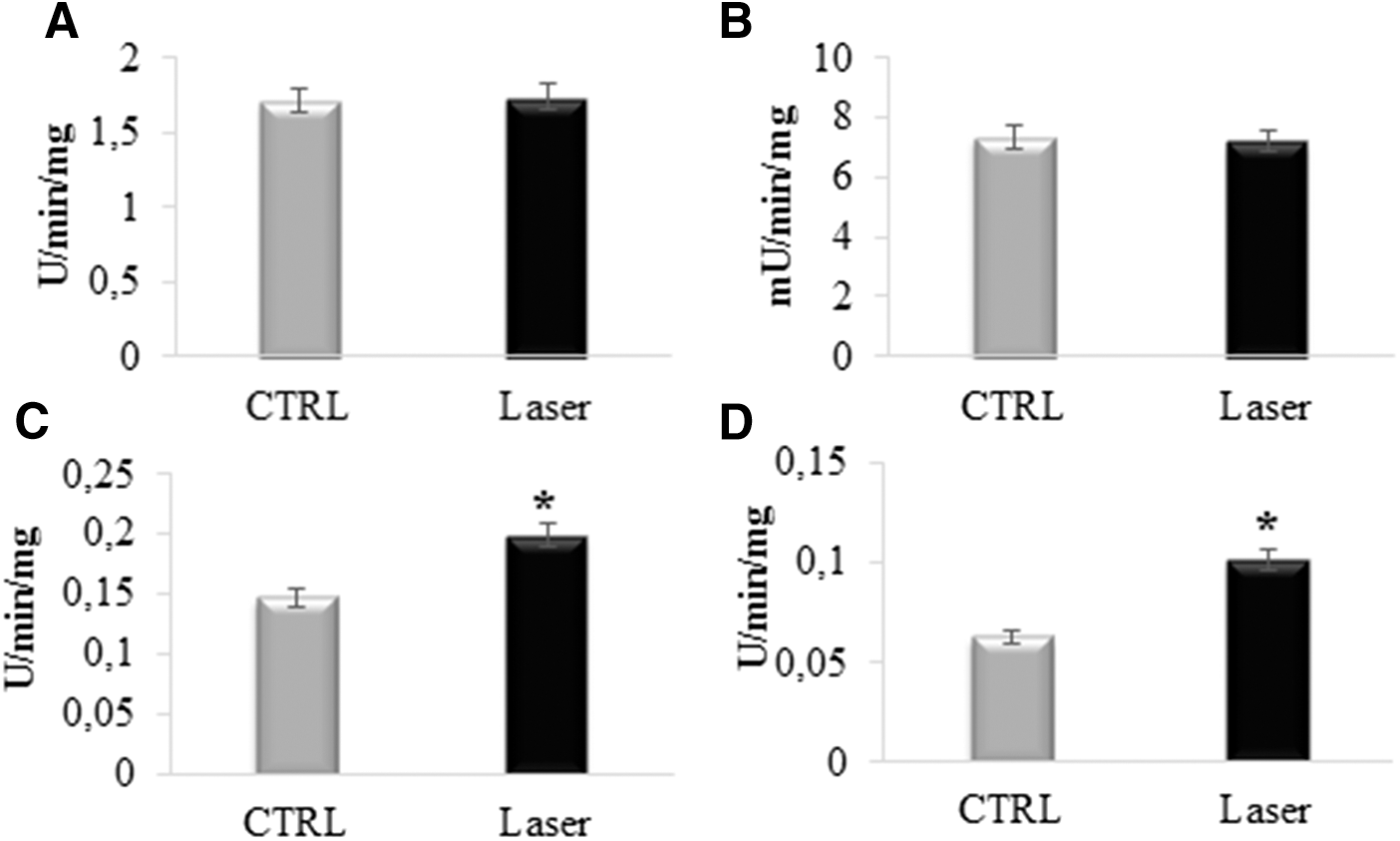
Assay of activity of respiring complexes in photoirradiated mitochondria.
So, to analyze the possible effect of photoirradiation on cytochrome c, this protein was irradiated before the assay of complex III. Data reported in Fig. 5 show that the complex III activity in the presence of irradiated cytochrome c is similar (p > 0.05) to that observed in the control condition, suggesting that cytochrome c does not change when treated with laser.
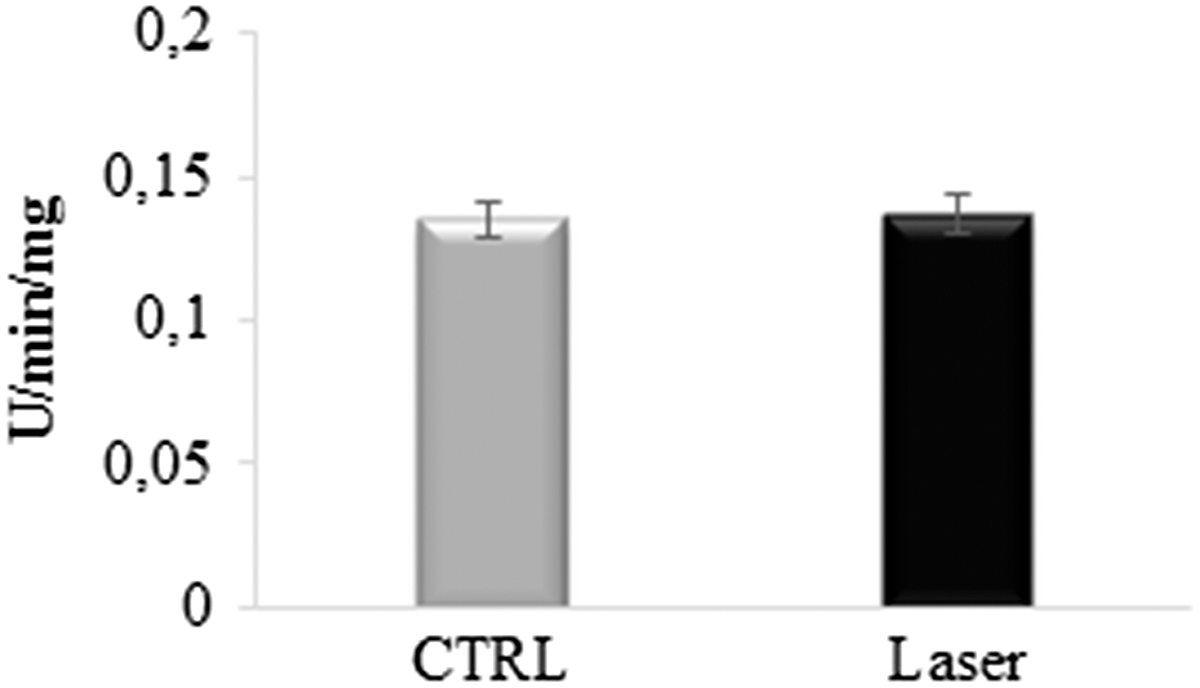
Complex III activity in the presence of photoirradiated cytochrome c. The graph shows the activity of complex III, measured with photoirradited cytochrome c. The data are expressed as mean ± SD, and the panel is representative of at least five experiments.
Evaluation of ATP synthesis in photoirradiated mitochondria
To understand whether the oxygen consumption increment is associated with a greater production of ATP, the activity of F0–F1 ATP synthase was evaluated in mitochondria, after the irradiation, compared with a not-treated control.
Figure 6 reports that the ATP synthesis in irradiated mitochondria (white square) is higher with regard to about 30% of the control (black circle) (p < 0.001), as yet observed for the oxygen consumption. This suggests that, in both conditions, the respiration is coupled with the energy synthesis, and that the laser does not induce an uncoupled status in the mitochondrial oxidative phosphorylation. To verify this aspect, the ratio between ATP produced and oxygen consumption, called P/O, was calculated. The data showed that in the presence of pyruvate/malate, the P/O is 2.3 ± 0.4 for the control and 2.6 ± 0.4 for the treated mitochondria.
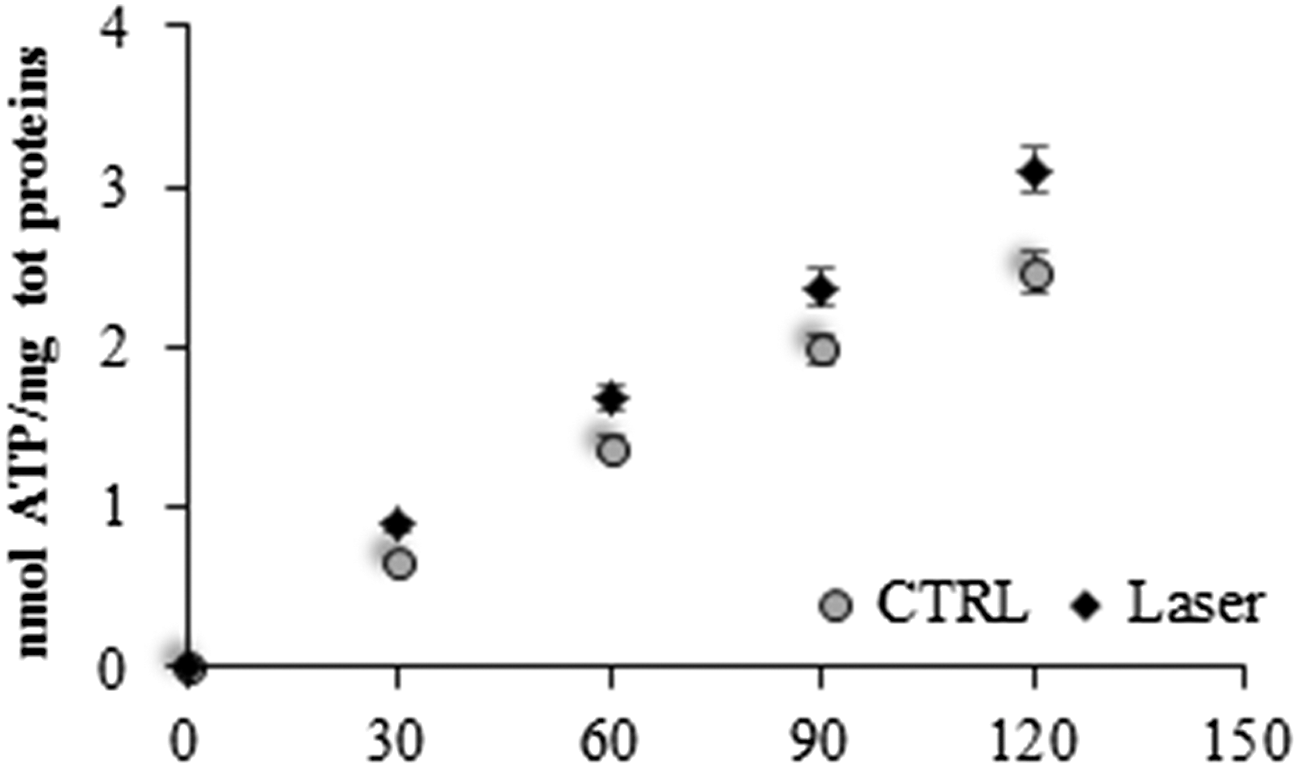
Assay of F0–F1 ATP synthase in photoirradiated mitochondria. The graph shows the rate of ATP synthesis in the first 2 min. White squares represent the activity in the photoirradited mitochondria, and black circle indicates the activity in the control. The data are expressed as mean ± SD, and the panel is representative of at least five experiments.
Discussion
The history of using light and color for health and healing (heliotherapy) can be traced back thousands of years in human history. The latter half of the 20th century witnessed a rediscovery of the science by researchers such as Gurwitsch (Russia), Mester (Hungary), Karu (Russia), Fröhlich and Popp (Germany), Passarella (Italy), and Hamblin (United States). There are now more than 40 years and 2000 published papers of formal research available in the area of PBM,
33
which established its merit as a genuine medical therapy. PBM is proposed as a non-linear process. Only the action of light at a low intensity and fluence is assumed to have stimulation on the cells; whereas higher light intensity and fluence generate negative effects, exhausting the cell's energy reserve as a consequence of a too strong stimulation.
34
Karu
35
observed that high fluences cause destruction of photoreceptors, which is accompanied by growth inhibition and cell lethality. Further, researchers have demonstrated that irradiation with fluences higher than 10 J/cm2 damages DNA.
36,37
To meet such evidences, Bensadoun and Nair
38
suggested the photobiostimulating optimal dose in the range of 2–3 J/cm2 for prophylaxis and not more than 4 J/cm2 for therapeutic effect and the application on a single spot on a lesion rather than a scanning motion over the entire lesion. The World Association of Laser Therapy (
However, our results show that the 808 nm higher-fluence (64 J/cm2), higher-power (1 W) diode laser therapy irradiated by the flat-top handpiece positively photobiomodulates mitochondria activities. Pastore et al. 16 have shown that 632 nm, low-fluence (2 J/cm2), low-power (10 mW) laser therapy increased the mitochondria oxygen consumption. Equally, our higher fluence acts on mitochondria, incrementing their oxygen consumption in the presence of pyruvate/malate or succinate. This result suggests that our therapy could affect the complexes III (cytochrome c oxidoreductase) and IV (cytochrome c oxidase) of the mitochondria respiratory chain. Our hypothesis is supported by our results of the higher fluence's effect on the single mitochondria complexes, which have an increment of about 33% of the complex III and 70% of the complex IV activity; whereas the activities of complexes I (NADH ubiquinone oxidoreductase) and II (succinate dehydrogenase) are not influenced.
Pastore et al. 17 established that cytochrome c oxidase is a specific target of PBM by 632 nm, 10 mW–2 J/cm2, laser therapy. Yu et al. 15 showed that a 660 nm laser (10 mW–0.6 J/cm2) increased the activity of the complexes III and IV and incremented the oxygen consumption in the presence of succinate (36%) or ascorbate/TMDP (41%), without affecting the complexes I and II. In the same paper, however, Yu et al. 15 indicate that as the fluence was increased up to 2.4 J/cm2, the oxygen consumption was not influenced by laser irradiation whereas the complex I increased; surprisingly, the activity of complex I was about 389% and those of complexes III and IV were 330% and 80%, respectively. This great increment in the activities of complexes I, III, and IV suggests that the 2.4 J/cm2 fluence 15 may be able to uncouple the ETC, with respect to F0–F1 ATP synthase, increasing free radical production and cellular damage. 40 For a long time, convincing evidence was provided that the potential sites of a side reaction with O2 are localized at complex I 41 and complex III. 42 Conversely, the more balanced increment of the activities of complexes III and IV proposes that the 2 J/cm2 fluence, 17 the 0.6 J/cm2 fluence, 15 and our 64 J/cm2 fluence photobiostimulate the mitochondria without uncoupling the system.
According to Pastore et al., 17 our results on cytochrome c show no alteration of its activity.
It is known in PBM that the photons of low-power visible light are absorbed by electronic bands belonging to some molecular photoacceptors or chromophores. A chromophore is a molecule, or part of a molecule, that imparts some decided color to the compounds of which it is an ingredient. Chromophores almost always occur in one of two forms: conjugated pi electron system and metal complex (chlorophyll, bacteriochlorophyll, flavoproteins, hemoprotein, sulfur protein, and porphyrins are examples of chromophores). 43 In animal cells, the majority of the cell chromophores are localized in the mitochondria; thus, it is not surprising, as a light-cell interaction is essentially a light-mitochondria interaction. 13
Getting back to the results, we suggest that our 808 nm, 64 J/cm2–1 W, diode laser therapy, applied by the flat-top handpiece, similar to the 632 nm, 10 mW–2 J/cm2, laser therapy 17 acts on the heme groups and/or the iron-sulfur protein of cytochromes that are associated with the mitochondria complexes III and IV; whereas the iron-sulfur protein and the flavoprotein of the complex I, the heme group and the succinic dehydrogenase activity of the complex II, and the electron shuttle molecules are not influenced by the laser. The data indicate that even for our high fluence, the heme-apoprotein or metal-protein micro-environment, rather than the heme group or metal ions per se, is responsible for the observed photobiostimulations.
Mitochondria play an important role in energy generation and metabolism. Mitochondria are sometimes described as a “cellular power plant,” because they convert food molecules into energy in the form of ATP via the process of oxidative phosphorylation. 2 Thus, an increase in the mitochondria respiratory chain and an increment in the mitochondria complex activities without augmentation in mitochondria ATP levels result in cell damage by oxidative stress. 44 The mitochondria irradiated by our 808 nm, 64 J/cm2–1 W, diode laser therapy increase the ATP synthesis with a phosphate/oxygen ratio (P/Olaser) = 2.6 with respect to the control (P/Ocontrol = 2.3). Thus, our therapy makes the mitochondria respiratory chain more efficient without uncoupling them and stimulates oxidative stress. In fact, it is known that if the P/O ratio in the presence of pyruvate/malate is around 2.5, the oxygen consumption is associated with the ATP synthesis, defying the coupled status, 27 which is present only when the mitochondria are intact and functioned. This hypothesis is also supported by our previous results on Paramecium cells, which after irradiation with the same 808 nm laser therapy by the flat-top handpiece showed a precocious fission rate for 10 successive generations without DNA damage, probably through an increase in metabolic activity, secondary to an energy transfer. 25
However, it is important to note that the irradiation may also affect other pathways involved in the energy production, such as Kreb's cycle and the pyruvate/malate shuttle, deserving a further investigation.
Conclusions
Our data show for the first time that even the higher fluences 64 J/cm2–1 W, similar to the low fluences, can photobiostimulate the mammalian's mitochondria respiratory chain without uncoupling them and can induce an increment in the ATP production. These results and our previous works 25 –27,29 suggest that the negative effect of higher fluences observed to date are not unequivocally due to high fluence per se but might be a consequence of the irradiation carried by handpieces with a Gaussian profile. In fact, thanks to the flat-top handpiece, expositions to higher-power energy (1 W) and higher fluence without ablative or thermal effects are now possible; irradiation in a short time (<60 sec) is possible on a target surface with a homogenous energy density.
Footnotes
Author Disclosure Statement
No competing financial interests exist.