
Abstract
Introduction
R
At this point, however, despite all these years of inquiry, there has been only one published report exploring the effect of R-NIR on the isolated enzyme CCO. In 2000, Pastore et al. published a study of the effect of helium-neon laser (632.8 nm) on the kinetics of CCO. 23 Increases in the observed kinetic constant for the reaction of CCO with cytochrome c were seen at high enzyme/substrate ratios when the enzyme was pre-irradiated. In contrast, a lowering of the kinetic constant occurred at low enzyme/substrate ratios. A mechanistic interpretation of these results was not offered. Similar results were obtained for the reaction of oxygen with CCO.
This study only involved a few combinations of CCO and cytochrome c concentrations, and was performed at a wavelength not as commonly associated with R-NIR-PBM as others. In addition, pre-irradiation of the enzyme presumes some sort of persistent effect; although most putative biological R-NIR-PBM effects also presume the same. Our current work was undertaken in the hope of confirming Pastore's results, widening the number of wavelengths used, and incorporating real-time irradiation during assays. In addition, we hoped to obtain data suitable for a more mechanistic explanation of the activation of CCO by R-NIR. Errede et al. have published a detailed study of CCO kinetics, 24 with an analysis of the results in light of several proposed mechanisms. The deduced rate equations for the reaction are complex, and include many parameters relating to various steps in the proposed mechanisms. A study of the kinetics along these lines could reveal specifics of the effects of R-NIR, and lead to mechanistic insights. In particular, it might be possible to relate the phenomenon of R-NIR-PBM to specific steps in the catalytic cycle. Our primary hypothesis was that R-NIR irradiation would have an effect on the kinetics parameters of the reaction of cytochrome c with isolated CCO, and that the magnitude and direction of these changes could be interpreted in the context of the reaction schemes proposed by other authors.
Preliminary to the spectrophotometric assay of cytochrome c oxidation, it was also deemed necessary to obtain new values for the milimolar extinction coefficients (ɛred or ɛox) of reduced and oxidized cytochrome c, and the reduced-oxidized difference (δɛred-ox). Values of ɛred (mM L−1 cm−1) at 550 nm reported or used in the literature range from 26.1 25 to 32.27, 26 whereas values for δɛred-ox range from 18.5 27 to 21.84. 26 Details of the determination of these values are not generally supplied; therefore, it is impossible to independently evaluate the quality of any given value. As precise values are necessary for both the determination of cytochrome c concentrations and the analysis of the reaction course data, it was believed to be necessary to determine these numbers in our own laboratory and under our own conditions. Values were desired for other wavelengths as well.
Materials and Methods
CCO was prepared from bovine heart according to the method of Soulimane and Buse. 28 Protein concentrations were determined by the DC Protein Assay (Bio-Rad, Hercules, CA). CCO concentrations were calculated using a molecular mass of 204,696. 23 The preparation was stored in small aliquots at −80°C in 10 mM Tris-HCl, 300 mM NaCl, and 0.1% Triton X-100, pH 7.6.
Horse heart cytochrome c was purchased from Sigma-Aldrich (St. Louis, MO, #C7752). For the determination of molar extinction coefficients, a stock solution of 1.97 mg cytochrome c in 10 mL deionized water was prepared in a 10 mL volumetric flask. Ten separate dilutions were prepared using calibrated pipettes to give expected absorbance values within the range of a Genesys 6 UVVIS spectrophotometer (Thermo Scientific, Waltham, MA). To each 1 mL sample in a cuvette, 5 μl 100 mM ascorbic acid was added to effect reduction. After 10 min, the sample was scanned from 350 to 600 nm at 2000 nm/min. To this sample, 100 μL of 10 mM potassium ferricyanide was then added to oxidize it. Again, a scan was performed after 10 min. Samples were tested in triplicate. Absorbance values were adjusted for volume changes and blank values obtained from the reagents. The molar extinction coefficients were obtained by fitting the data to Beer's Law using a linear least squares analysis. Both the absorbance and the concentration data were weighted using standard propagation of error techniques for all known or estimatable error sources.
Cytochrome c for activity assays was prepared fresh for each experiment by dissolving ∼3 mg/mL cytochrome c in a buffer containing 0.1 M 2-(N-morpholino)ethanesulfonic acid (MES), 10 μM ethylenediaminetetraacetic acid (EDTA), pH 6.0. A total of 10 μL 100 mM ascorbic acid/ml cytochrome c solution was added to effect reduction. After 15 min incubation, excess ascorbic acid was removed by three successive ∼15/1 dilution and concentration cycles with this buffer using an Amicon Ultra-15 centrifugal filter (Merck Millipore Ltd., Co. Cork, Ireland) spun at 5000g for 20 min for each cycle. The resultant concentrate, ∼600 μL, was then diluted to a working strength of ∼200 μM using the same buffer. The concentration of this stock solution was determined spectrophotometrically at 550 nm, and appropriate volumes for the desired assays were calculated.
Activity assays at room temperature (23°–24°C) were performed by the method of Errede et al.
24
Appropriate volumes of cytochrome c stock were added to an assay buffer of 0.1 M MES, 10 μM EDTA, and 0.1% n-Dodecyl β-D-maltoside (Sigma-Aldrich, #D4641) pH 6.0. The reaction was initiated by the addition of an aliquot of CCO concentrate, and followed at 550 nm for 5–20 μM cytochrome c, or 415 nm for 1 μM cytochrome c, for 3 min in the Genesys 6. Each assay was performed in triplicate at six CCO concentrations per cytochrome c concentration. The concentration of reduced cytochrome c was calculated at 1 sec time points by the equation:
Values for the reduced-oxidized milimolar extinction coefficients δɛ550red-ox and δɛ415red-ox are reported in the Results section. The observed first order rate constant kobs was calculated from the slope of a plot of ln(ferrocytochrome c) versus time. The second-order rate constant k′ was calculated from a plot of kobs versus (CCO) using a weighted linear least squares analysis (PSI-Plot, Poly Software International, Pearl River, NY). Statistical significance was tested using a two tailed Student's t test, with significance set at p < 0.05.
Pre-irradiated R-NIR experiments were conducted by irradiating the CCO concentrate aliquot in a Hamilton syringe for 80 sec under 50 mW/cm2 irradiation from a 670 or 830 nm LED array (Quantum Devices Inc., Barneveld, WI) for a total exposure of 4 J/cm2. The CCO was then added to the assay buffer immediately to initiate the reaction in the normal manner. No significant heating of the enzyme was observed. Experiments were performed at 1, 5, 10, and 20 μM cytochrome c, and controls were performed for each experiment.
Real-time irradiation experiments were performed by initiating a reaction as described, setting the cuvette into the spectrophotometer, and placing a 660 nm LED (10 mm WP813SRC/F Super Bright Red, Kingbright, City of Industry, CA) powered at 19 V and 50 mA (full power) into the top of the cuvette, producing 4.6 mW/cm2 at the level of the detector beam. The LED was turned on immediately. Every 30 sec, the LED was turned off for 5 sec in order to obtain an absorbance reading, and the reaction terminated after 3 min. No heating caused by the R-NIR was observed. Experiments and controls were performed at the substrate concentrations given.
Results
Beer's law plots of absorbance at 550 or 415 nm versus cytochrome c concentration are shown in Fig. 1. Calculated milimolar extinction coefficients are: ɛ550red = 29.1 ± 0.4 L−1 cm−1, ɛ550ox = 8.60 ± 0.15 L−1 cm−1, ɛ415red = 140 ± 2 L−1 cm−1, and ɛ415ox = 89.0 ± 1.1 L−1 cm−1. Values of the calculated reduced-oxidized milimolar extinction coefficients are δɛ550red-ox = 20.5 ± 0.2 L−1 cm−1, and δɛ415red-ox 51.0 ± 2.0 L−1 cm−1.
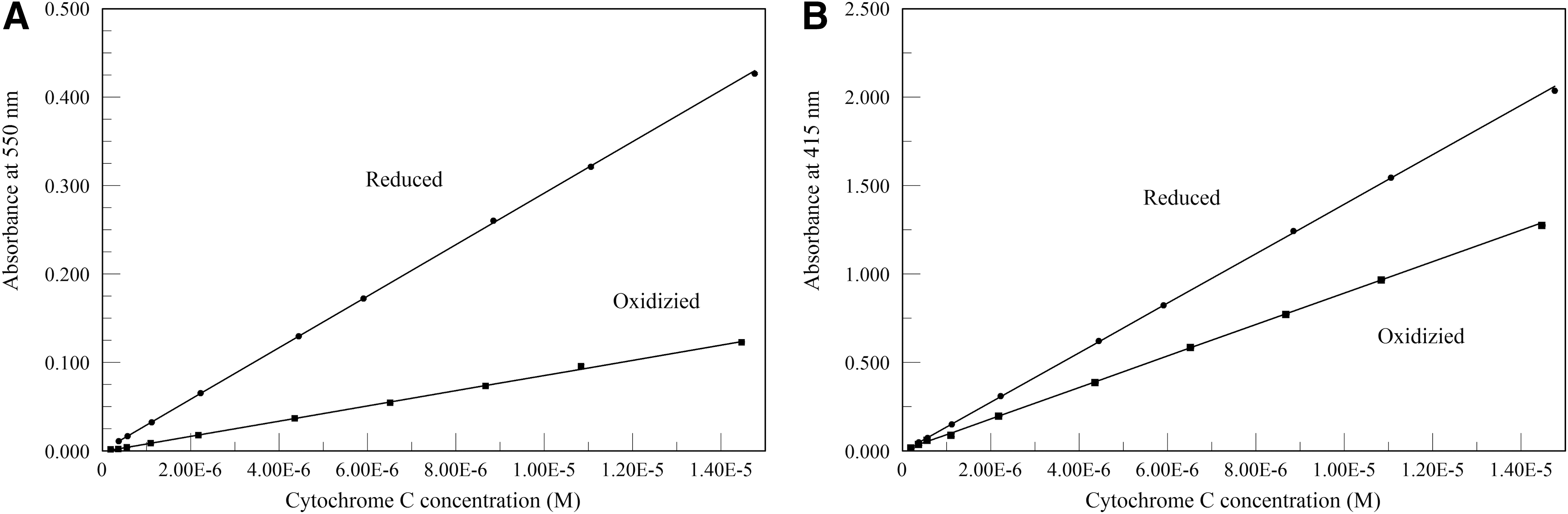
Beer's law plots of absorbance at 550 nm
CCO shows pseudo-first order kinetics throughout all substrate levels studied.
29
The observed first order rate constant (kobs) for any particular run can be calculated from:
Plots of ln(ferrocytochrome c) versus time were in all cases linear (data not shown). The second order rate constant k′ can be calculated from:
Therefore, the slope of plots of kobs versus [CCO] at any particular substrate concentration will give the rate constant k′, which relates the turnover number (TN) of the enzyme reaction to CCO and total cytochrome c concentration. This relation is given as:
Plots of kobs versus (CCO) for the 670 and 830 nm pre-irradiated experiments are shown in Fig. 2. Separate controls were run for each wavelength and substrate concentration set. The k′ values from the slopes of these figures are given in Table 1. In no case did the second order rate constant show a statistically significant difference from the corresponding control value. Similarly, the 660 nm real-time irradiation values also show no statistically significant difference from the controls (Fig. 3 and Table 2).
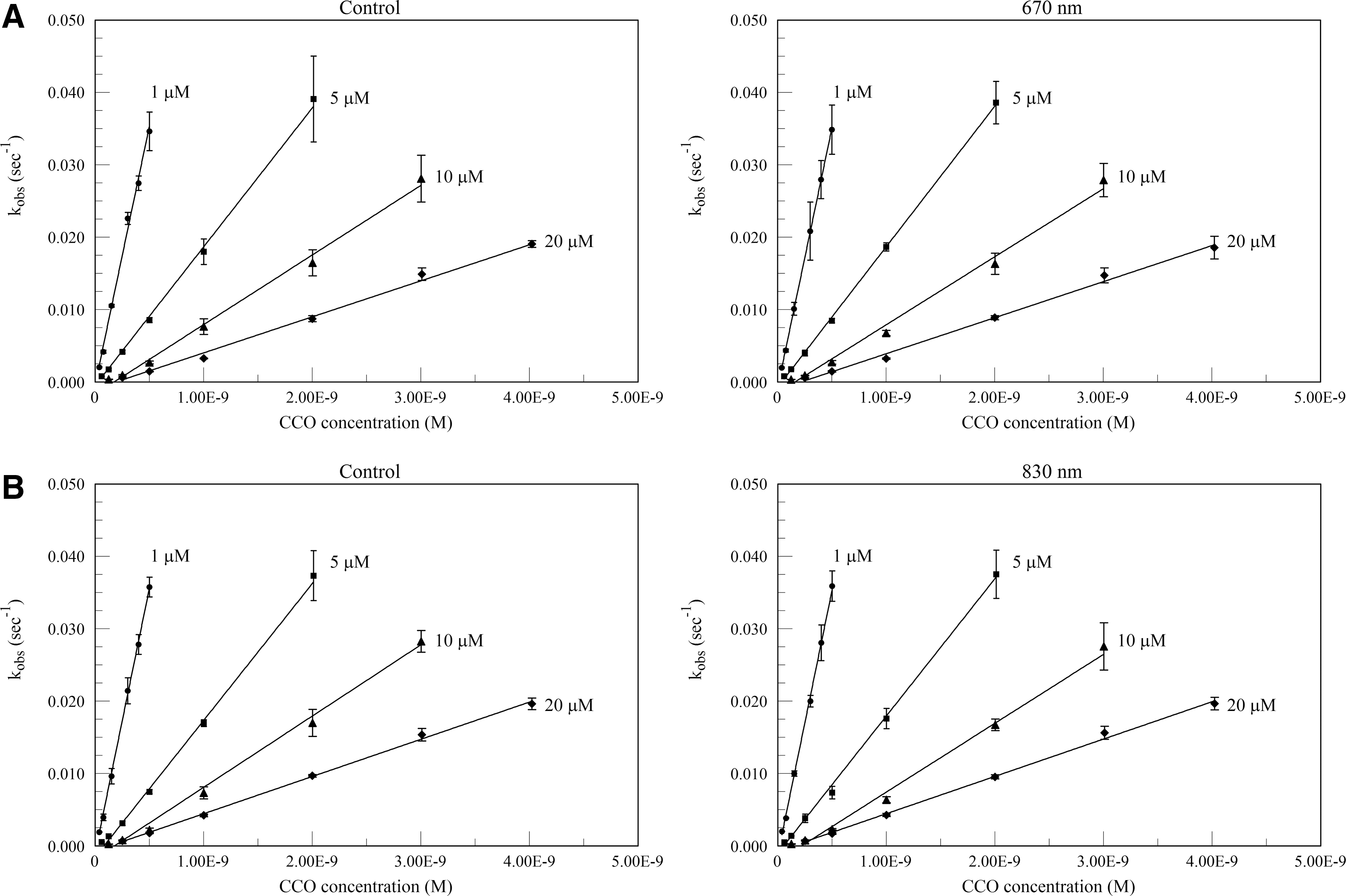
Plots of the pseudo-first order rate constant kobs versus cytochrome c oxidase (CCO) for control (no near infrared [NIR]) and pre-irradiated CCO at 1–20 μM substrate concentrations. Panel
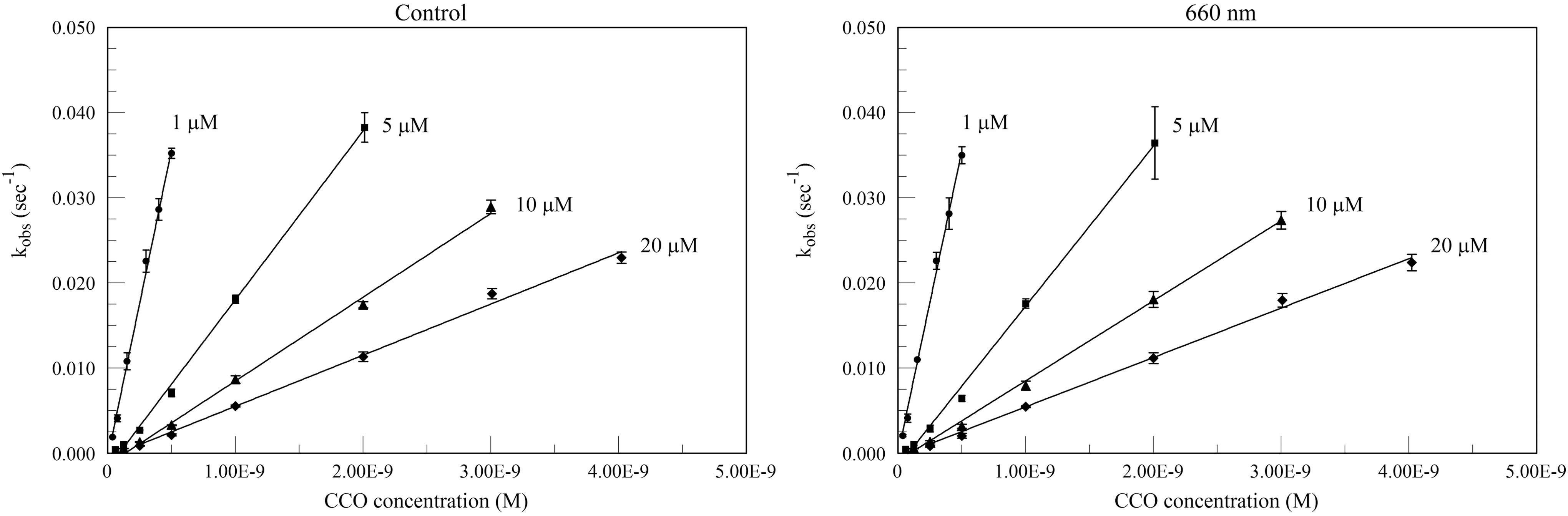
Plots of the pseudo-first order rate constant kobs versus cytochrome c oxidase (CCO) for control (no near infrared [NIR]) and real-time irradiated CCO at 660 nm. Assays were run for each substrate concentration at a variety of CCO concentrations, with unirradiated controls run at the same time.
Values are mean ± SD. No significant differences (p < 0.05) between irradiated values and corresponding control values.
Values are mean ± SD. No significant differences (p < 0.05) between irradiated values and corresponding control values.
Discussion
Contrary to Pastore's previously published report, 23 in our study R-NIR showed no effect on the rate of oxidation of cytochrome c by isolated CCO. At a wide range of both substrate and enzyme concentrations, reactions with or without R-NIR were kinetically indistinguishable. Because pre-irradiation of the enzyme presumes some sort of persistent effect, assays were also run with R-NIR irradiation applied concurrently. Again, no effects were seen. This lack of kinetic modulation does seriously call into question the CCO activity model of R-NIR-PBM and its role as the primary cellular R-NIR receptor. 3
Although our present study fails to find an effect of R-NIR on the activity of isolated CCO, there have been reports of an effect on CCO activity in less homogenous systems. Silveira et al. have published studies of CCO activity assays on rat tissue homogenates after ulceration 18 and blunt force trauma. 19 In both cases, an increase in CCO activity that was attributed to NIR-PBM (904 nm) was observed. Unfortunately, this type of study cannot distinguish between increased expression and increased specific activity of CCO. In addition, it doubtful that an activated state can survive extraction, as the half-life of such an activated state is likely to be shorter than the lengthy extraction stages. It seems more likely that the observed results are the result of increased expression rather than specific activity.
Hu et al. have studied the effects of R-NIR (632.8 nm) on melanoma cell cultures, reporting a 200% increase in CCO activity. 22 The cells were lysed immediately after irradiation, centrifuged, and assayed. Thus, the time span between irradiation and assay is much shorter than the animal tissue studies, but the issue of activated state lifespan cited is still operative. Because of the low output of the laser employed (7 mW), irradiation time might also have been sufficiently lengthy to permit changes caused by expression and/or cell proliferation.
Houreld et al. have reported on irradiation (660 nm) of human fibroblast mitochondria concentrates, with an observed increase of ∼50% in CCO activity, 20 whereas Yu et al. have reported on rat liver mitochondria concentrates also irradiated at 660 nm, with an increase in CCO activity of ∼80%. 21 These experiments come nearer to answering the question of whether CCO itself is activated by R-NIR. These isolated mitochondria, when irradiated, likely eliminate expression confounding factors, and should give information pertinent to the local environment of CCO. If CCO is activated in mitochondria by R-NIR, but not when irradiated in isolated form, we could conclude that the local environment within the mitochondrial membrane is such that it sensitizes the enzyme molecule to R-NIR energy, and that this activation is both persistent and resistant to subsequent extraction and solubilization steps. Such an effect, likely to involve a conformational change rendering electron transfer steps quicker, would therefore result in a structural rearrangement of a nearly permanent nature. We have no evidence of such a rearrangement at this time.
Several issues involving all of these studies do present themselves. As they do not directly involve isolated, purified CCO, numerous confounding factors are present. This can be both a problem and an opportunity, as the presence of differing environments can lead eventually to conclusions that are more pertinent to real life. The wide spread in reported specific activities makes it difficult to compare studies, whereas the lack of sufficient information on the CCO assay methods makes it hard to adequately evaluate the results presented in each study. In particular, it is hard to determine whether detergents were used in the mitochondrial assays. If no detergent is used, CCO is mostly inaccessible because of the mitochondrial membrane. If detergent is used, then the result would be the same as solubilized CCO, unless the hypothesis of a persistent, stable, activated state is correct. As no detergent was reported by Houreld 20 or Yu, 21 it is possible that the results are actually a measurement of the state of the mitochondrial membrane, 30 and that increases in activity represent membrane disruption.
Because these experiments were based on the kinetics methods of Errede, 24 and were intended to replicate and extend the R-NIR-PBM work of Pastore, 23 it may be instructive to compare the current results with the two previous studies. Table 3 contains a comparison of the second order rate constant and turnover numbers for these studies. In the case of Errede, k′ values were estimated from the figures, whereas for Pastore, k′ values represent single determinations. In all cases, our k′ values are nearly twice those of Errede's. Because Errede based his enzyme molar mass on a cytochrome a basis, and we have used the molar mass calculated by Soulimane, 28 which is based on two cytochrome a moieties per enzyme molecule, this is understandable and expected. When this difference is considered, our k′ and TN numbers are consistent with Errede. Pastore's values, however, are two to three orders of magnitude lower. Because values this low are not reported in the literature, one must postulate that there is some vital difference in the enzyme preparation used or the assay techniques employed. Perhaps the lack of any reported use of detergent needed for proper CCO solubilization and stabilization 30 in the assay mixture may have resulted in protein aggregation or a drastic loss of enzymatic activity. The reported results may, therefore, perhaps be experimental artifacts imposed on an extremely weak enzymatic reaction.
Values are mean ± SD.
Calculated from average of all six experiments pooled, whether control or irradiated.
Turnover number (TN) = k′(cytochrome c).
k′ estimated from the published figures.
k′ estimated from kobs/[cytochrome c oxidase (CCO)] values given.
There could still be a role for CCO in R-NIR-PBM. Although the activity may not be affected, it may be possible that R-NIR reception by CCO in some manner results in increased expression of the same enzyme. 13,14 It is difficult, however, to envision how such a result could be achieved. Perhaps CCO modulation by R-NIR does not affect the rate of cytochrome c oxidation, but rather increases its efficiency by increasing the proton pumping rate per electron transferred, as argued in a study of isolated rat liver mitochondria. 12 This could improve the production of adenosine triphosphate (ATP), with all of the implied cellular benefits. In addition, more attention is being given to the signaling roles of ATP, expanding the possible beneficial effects of increased ATP production. 31 Nitric oxide (NO) has also been implicated in R-NIR-PBM. Application of 670 nm light protects cardiomyocytes from reoxygenation injury in an NO- dependent mechanism, at least partially derived from non-NO synthase sources. 32 CCO has been shown to have an NO synthesis activity, and can be stimulated by light, in this case 590 nm. 33 Therefore, CCO could still be the receptor for R-NIR, whereas NO production and signaling pathways could provide the means by which photobiomodulation is effected. 4
We have only studied one part of the CCO reaction. In this work, the other substrate, oxygen, is considered to be saturating and not rate limiting. It is possible that the kinetics of the reduction of oxygen by CCO may be affected by R-NIR even if the cytochrome c oxidation may not. It is our intention to study oxygen uptake in a respiration chamber using the Clark electrode technique. 34 This would enhance our ability to measure CCO kinetics with real-time irradiation, as the respiration chamber is clear, and R-NIR irradiation would not interfere with detection, as it does in a spectrophotometer. Another possible avenue of exploration would entail the incorporation of CCO in lipid vesicles, 35,36 as this might afford a more physiologically relevant environment, without the multiplication of confounding factors when intact mitochondria are studied.
Conclusions
The oxidation of cytochrome c by isolated CCO has not been shown to be affected by R-NIR irradiation, whether applied prior to or concurrently with the enzymatic assays. This lack of effect by R-NIR calls into question the CCO activity model of R-NIR photobiomodulation. Further work on the effect of R-NIR on the reduction of oxygen is planned.
Footnotes
Acknowledgments
This work was supported by Hanger Orthopedic Group Inc. (to Dr. Whelan), the Bleser Endowed Chair in Neurology (to Dr. Whelan), as well as the Baumann Research Endowment (to Dr. Whelan). These funding sources had no involvement in the preparation of this article.
Author Disclosure Statement
No competing financial interests exist.