
Abstract
Introduction
A
To this end, we present two clinical trials, one related to pain relief and the other where PBM was clinically effective in providing dental anesthesia, followed by discussion of the evidence that a neural basis underlies PBM effectiveness.
Clinical trial evidence
In a clinical trial of PBM for chronic neck pain, Chow et al. demonstrated that 830 nm PBM at 300 mW, 9 J/point, ED: 20 J/cm2, provided statistically significant and lasting pain relief. 1 The trial of 90 patients was randomized, placebo-controlled, and double blind. There were no side effects and specifically no adverse effects on sensation.
The second clinical trial examined the effectiveness of PBM for dental anesthesia in people requiring tooth extraction before orthodontic procedures. 2 Again the trial was randomized, double-blind, and placebo-controlled, and PBM was delivered by pulsed Nd:YAG at 0.2 W,15 Hz, 60–87 mJ energy pulse; PD: 0.3–0.45 J/cm2; energy density, 73–107 J/cm2; total energy, 211–312 J. PBM effectiveness was compared with the topical anesthetic cream EMLA and was statistically significant in providing more effective pulpal anesthesia than EMLA. There was also a concurrent but significant decrease in pulpal sensitivity after PBM as measured by subjective electric pulp testing. A follow-on morphological and histological study of all extracted teeth showed that there was no PBM-related damage and no significant temperature increase. 3 The anesthetic effect of PBM was reversible and as in the Chow et al. trial, there were neither side effects nor evidence tissue/nerve damage. In both trials, PBM was delivered transdermally to the ectoderm or gingiva both characterized by dense beds of C fiber endings with penetration of the dermis. The underlying tissue including the Aδ axons was well within the penetration depths of the laser at the wavelengths and parameters used (Fig. 1)
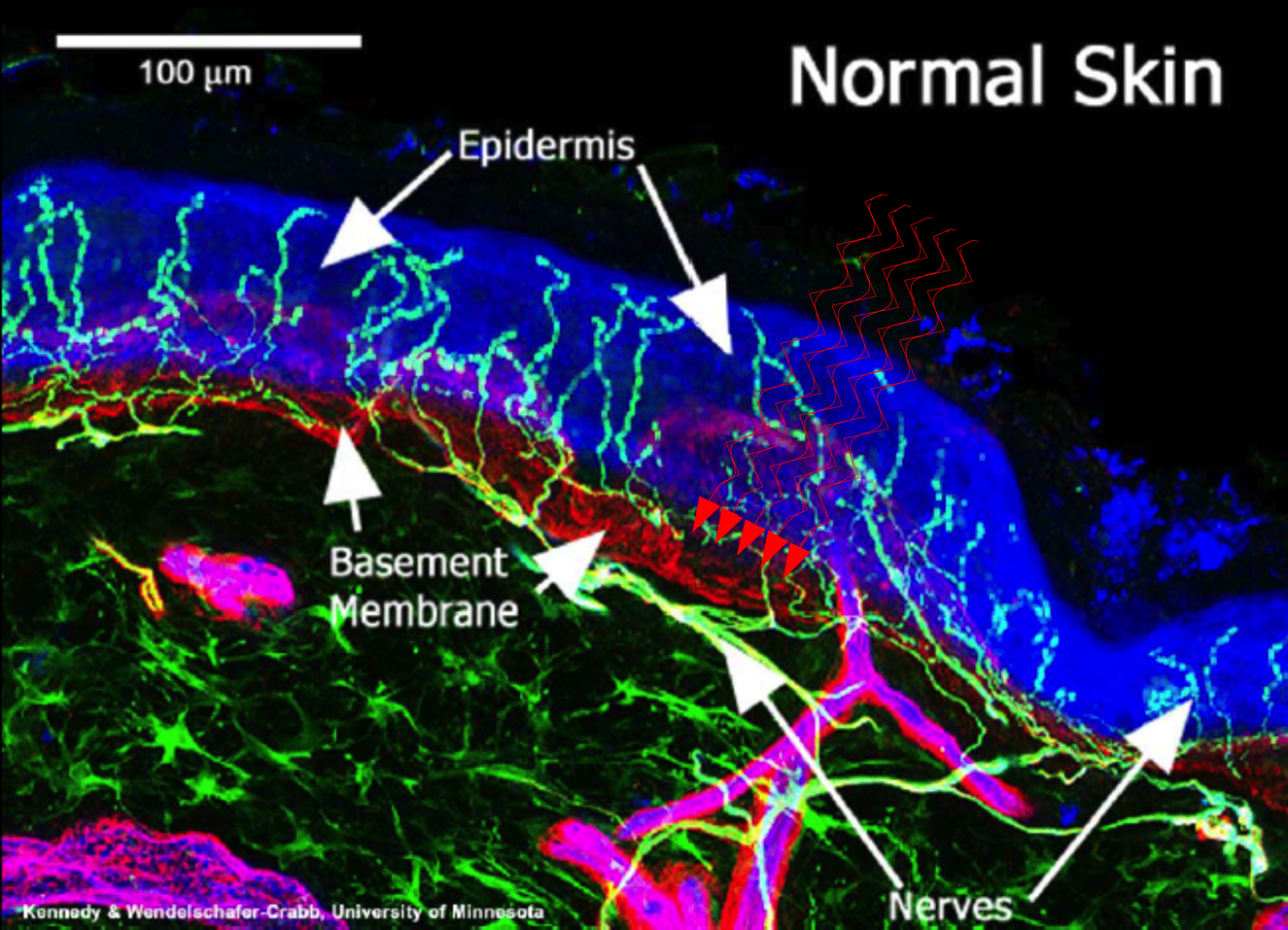
Normal human epidermal and papillary dermis innervation. Nerves are localized with antibody to PGP 9.5 and basement membrane demarcated with antibody to type IV collagen. Epidermal nerve fibers appear aqua and lie within the blue epidermis (E). The subepidermal neural plexus (SNP) appears green or yellow. The dermal-epidermal junction appears red. Capillaries (C) appear magenta. Nerve fibers (green and aqua) course in bundles through the dermis and branch in the papillary dermis to form the SNP. Fibers arise from this plexus and penetrate and epidermal-dermal basement membrane to enter the epidermis. Epidermal nerve fibers are abundant and uniformly distributed in normal human skin.
These two very different clinical trials demonstrate the nexus between PBM and pain relief with statistically significant confirmation of a neural basis for PBM effectiveness acting at least in a major part through neural inhibition/conduction block. Scientific studies discussed hereunder provide further strong evidence of PBM effectiveness in neuro-electrophysiology data from studies in humans and animals in vivo, in situ, ex vivo, and in cell culture studies.
Bedside to bench–electrophysiology studies
It is unusual to have bedside data such as the clinical trials as the starting point for bench studies to explain the concept that PBM acts through neural inhibition. Although it is well established that anesthetic and analgesic agents cause conduction block, it is the electrophysiology studies such as those discussed hereunder that confirm that PBM like these agents acts on the nervous system causing conduction block and is reversible. The concept that light can directly affect nerve conduction is not a new phenomenon. One of the earliest electrophysiological studies of light effects on nerves showed that 490 nm LED irradiation induced neural inhibition in the abdominal ganglion neurons of the marine mollusc Aplysia Californica. 4 Since then, many studies have reported further evidence from studies in mammals that PBM induces neural inhibition or conduction block and a number of these are discussed in the next section.
Electrophysiology and PBM—studies of human nerve
Clinical electrophysiology on human nerves reports measurements of the time from stimulation to the onset of the action potential—latency and the amplitude—a measure of compound action potentials reflecting conduction velocity. These clinical studies can include somatosensory-evoked potentials (SSEPs) and/or compound muscle action potentials (CMAPs), one representing sensory nerve responses and the other the neuromuscular junction—more correctly the tripartite synapse—response. The majority of PBM electrophysiological studies unfortunately report only the latencies and do not include the amplitudes. Amplitudes are particularly important in such studies as they provide the conduction velocity data, whereas increased latency is a nonspecific indicator of impaired or delayed neural conduction. Decreased or dispersion of amplitude traces directly represent slowing of conduction velocities, which at levels of 25–30% are clinically considered to reflect conduction block or inhibition, thus the importance of reporting latency and amplitude. In clinically based electrophysiological studies used for diagnostic purposes such as for the diagnosis of carpal tunnel syndrome, increased latencies are considered to be an indicator of demyelination. In experimental studies using laser, outlined hereunder, increased latencies immediately after PBM can be directly attributed to the effects of light on nerve physiology and not demyelination. This is particularly so as the return to normal latencies and conduction velocity occurs within a short time frame, which is not consistent with demyelination and remyelination.
Of particular interest are several relevant PBM electrophysiological studies undertaken on healthy individuals. Nerves studied include the median, radial, sural, and superficial radial nerves with PBM delivered transcutaneously or through gingiva to the maxillary branch of trigeminal nerve. These have been previously reviewed in detail in a systematic review. 5 Only one study measured both amplitudes and latency parameters simultaneously and reported both increased latency and decreased amplitudes. 6 A typical and early example was the randomized control trial by Snyder-Mackler and Bork. 7 Baseline nerve conduction studies of SSEPs were determined before He:Ne PBM at 632 nm or sham on 40 age- and sex-matched participants with no underlying pathology. There were statistically significant increased latencies of the superficial radial nerve—indicative of neural impairment or conduction delay. Another series of studies using a similar methodology with near infrared wavelengths (780–904 nm) in continuous mode similarly demonstrated increased latencies in the majority of studies. Of particular relevance to methodology and parameter reporting are two studies related to temperature, illustrating different outcomes. 8,9 In the Basford study, the arms of the participants were immersed in a water bath at 40°C while Baxter carefully controlled the ambient temperature between 21°–25C°. Basford et al. delivered 830 nm, continuous wave (cw) laser irradiation (ED 9.6 J/cm2) over the median nerve of healthy volunteers (n = 33), showing no evidence of amplitude change although a 3–4% decrease in latency was noted in the laser-treated group. Baxter et al. in contrast using the same parameters and experimental methodology (n = 51) showed increased latency. As nerve conduction is strongly influenced by ambient temperature, both studies show the importance of considering all the variables in the methodology and the parameters to compare outcomes. Of interest and importantly from an experimental methodology perspective was the recent study by Grandinetti et al. who applied PBM to people with different skin pigmentation. 10 There was no statistical difference between any wavelengths in skin temperature changes with different skin pigmentations.
Delivery of PBM in pulsed rather than continuous mode is a further variable with specific effects on neural inhibition shown in these studies. For example, the same methodology using with cw 830 nm laser irradiation (ED: 1–5 12.0 J/cm2) resulted in increased latencies, 9,11 whereas pulsed delivery of the same wavelength at similar energy densities did not cause a significant change. 12,13 The variability of outcomes was also demonstrated in a study of sural nerve, where pulsed wave 830 nm laser irradiation at 5.1 J/cm2 but not at 7.65 or 2.55 J/cm2 increased latencies. 14 Pulsed wave 904 nm (73 Hz) laser irradiation slowed conduction velocity (CV) of the superficial radial nerve with 120 sec irradiation per point, but 20 sec irradiation per point produced no effect on nerve conduction. 15 Interestingly, there is only a single study using LED, which at 950 nm shows inhibition of nerve conduction velocity in sural nerves with no increase in skin temperature. 16 Of note is that the Chan et al. clinical trial using pulsed Nd:YAG at specifically relevant clinical parameters that did effect anesthesia, consistent with conduction block, was not associated with any temperature change.
Relevant to dental practice is a study where 632.5 nm PBM was delivered intraorally to the gingiva overlying the posterior superior branch of the maxillary nerve followed by electrical stimulation over the ipsilateral infraorbital foramen. 17 Somatosensory trigeminal-evoked potential (STEP) amplitudes were measured before and after laser application. Within 2 min after PBM over the maxillary nerve, STEP amplitudes were reduced by up to 72% lasting for 20 min. Such conduction block may well be of clinical importance in explaining PBM effectiveness in dental applications and in the treatment of trigeminal neuralgia. 18
Electrophysiology and PBM in animal studies in vivo, in situ, and ex vivo
In addition to the human electrophysiological studies are studies in several animal models. These series of studies in vivo illustrate the effect of laser irradiation on nerve conduction, predominantly velocity 19 –27 (Table 1) and ex vivo 28 –32 Table 2. Only a single study in rats systematically measured changes in SSEP and CMAP latencies and amplitudes simultaneously at baseline and after transcutaneous PBM or sham PBM with 808 or 650 nm laser irradiation. 27 The study showed a statistically significant decrease in SSEP and CMAP latencies with both 808 or 650 nm wavelengths, applied at four equidistant points overlying rat sciatic nerve. This occurred by 10 min and lasted up to 20 min. Both wavelengths also statistically significantly reduced CMAP and SSEP amplitudes up to 32.6% consistent with conduction block. Conduction block is defined clinically as a reduction in amplitude of more than 30%. After PBM, the rats exhibited normal behavior with no evidence of peripheral nerve damage or impairment of nerve conduction. Electrophysiological measurements repeated 48 h later showed that nerve conductions had returned to baseline.
λ, wavelength; CAP, compound action potential; CMAP, compound muscle action potential; CV, conduction velocity; cw, continuous wave; ED, energy density; ES, electrical stimulation; J, joules; mW, milliwatts; nm, nanometer; NR, not reported; P, pulsed; PD, power density; SSEP, somatosensory-evoked potential.
In an innovative and interesting ex vivo study, again related to dental procedures and neural inhibition, Orchardson et al. 32 inserted excised rat dorsal spinal nerves into the pulpal cavities of freshly extracted human teeth immersed in isotonic Krebs solution at 37°C. SSEPs were measured after supramaximal stimulation to the exposed part of the root before PBM delivered by Nd:YAG laser in scanning mode at one of two energy densities. This enabled the calculation of the effect of the PBM on the Aδ axons, which showed significant evidence of conduction block with return to baseline after 7 min. The study is particularly interesting as it used Nd:YAG at parameters similar to those used in the clinical trial of Chan et al., where dental anesthesia was obtained after PBM. What is particularly important in the Orchardson study is that both rat and human spinal nerves are made up of sensory neurons. Approximately 80% of these are small diameter unmyelinated C fibers conducting at ∼1–1.9 ms 33 with the remaining larger diameter thinly myelinated Aδ neurons conducting at ∼6.7–23.7 ms. Both neuronal subsets are nociceptive and convey noxious stimulation. Both studies are evidence of neural inhibition of the intradental pulpal nerve.
Another set of experiments by Orchardson's group examined both He:Ne and Nd:YAG effects on the intradental nerve using in situ teeth of anesthetized ferrets to determine the effect on dentine sensitivity. 34 Continuous electrophysiological recordings were made before, during, and after application of the laser. Responses of the intradental nerve were dose dependent with suppression of nerve excitability followed by recovery with no pulp damage with 1 W, whereas 2 W or more caused irreversible pulpal damage. Also, 632 nm 1 mW PBM in the same study did not show a change in intradental nerve activity after electrical stimulation. This is not unexpected as the low output power of the laser and lack of tooth penetration at this wavelength would not deliver sufficient energy to the nerve to alter conduction.
A further series of studies used Nd:YAG to determine effects on rat and cat dorsal roots and rat peroneal and sciatic nerve. 35 Although they report neural inhibition, and in particular selective inhibition of C fibers, they attributed the outcome to thermal effects as in most of the experiments; temperatures increased several degrees unlike those of Chan et al. As mentioned earlier, temperature change provides a note of caution regarding the reporting of temperature as a number of studies using Nd: YAG are confounded by significant temperature increases and irreversible damage to nerve when high intensities (>1 W) are used. This contrasts with studies using Class 3B devices or lower, where thermal responses are minimal or where PBM is delivered at lower powers (<500 mW) and showed that the electrophysiological effects were reversible.
PBM inhibits electrophysiological response to noxious stimulation
Neural inhibition/conduction block as reported in animal and human studies is indicative of the potential for PBM's pain-relieving effects. Several studies provide evidence of direct inhibition of neural activity in response to a variety of noxious and proinflammatory stimuli (Table 3). Specifically these include heat 36 ; chemical: formalin Shimoyama et al. 54 ; turpentine, 37,38 bradykinin 39 ; mechanical, 25,34 and electrical stimulation. 29,40 An example of these studies is that of Tsuchiya et al., where 830 nm laser to saphenous nerve significantly decreased SSEPs after noxious pinch, cold, and heat. 24,37 This was selectively nociceptive, as non-nociceptive brush stimulation transduced by Aβ fibers that mediate light touch was not suppressed.
APs, action potentials; DRG, dorsal root ganglion; EPs, evoked potentials; W, watts.
Tsuchiya et al. also demonstrated a specific anti-inflammatory effect as 830 nm laser applied to rat paw after the injection of the proinflammatory formalin or turpentine blocked the consequent nerve activity generated by the stimulus. Moreover, rats treated at birth with capsaicin that destroys C nociceptors did not respond to either turpentine injection or PBM, again demonstrating subtype selectivity. In another study using noxious heat to stimulate isolated nociceptors of rat tongue, 830 nm laser decreased the lingual nerve firing rate as assessed by single-fiber recordings, again demonstrating the nociceptor-specific effect. 36
Neurophysiological and morphological responses to PBM—ex vivo and cell culture experiments
In addition to electrophysiological studies, neurophysiological studies employing ex vivo, cell, and tissue culture experiments of peripheral sensory axonal responses to PBM are important additions to the armament required to define cellular responses both morphological and neurophysiological, resulting in neural inhibition.
Cell culture employs primary cells directly dissected from human or animal fetal tissue or more generally rodents such as rats and mice. Some primary neural cells and cell lines relevant for neural studies are now commercially available, although cell lines are genetically transformed. With respect to neuronal tissue cell lines, except for PC2 cell lines, they do not provide the unique organization of neurons with their specialized cell bodies and the lengthy axons (up to 1 M) of peripheral nerve nor the axons and dendritic branching of the central and autonomic neurons. Another proviso in assessing data from cell cultures is that there is no circulating blood supply or systemic delivery of molecules nor gas exchange, so that the combination of data from animal studies that can be benchmarked against the culture studies is ideally done in tandem. This is particularly relevant when considering the production of molecules released from nerves after injury or surgery, such as tumor necrosis factor (TNF), nerve growth factor (NGF), and bradykinin, which are both painful and proinflammatory.
A novel study by Jimbo et al. 39 demonstrates the importance of considering the complexity and interactions between the cell body and axon in vivo. They employed cultured murine dorsal root ganglion (DRG) neurons with cell diameters and responsiveness to the proinflammatory and stimulating molecule bradykinin, indicating that they were C and Aδ nociceptors. The culture system used was developed by Noda et al., 41 where the neuronal cell bodies were maintained in a central chamber with their axons growing through a separation barrier to the outer chamber. This separated the medium in each chamber. Bradykinin added to the axons of outer chamber evoked action potentials of the cell body of the inner chamber, measured by patch clamping. When this was followed by 830 nm PBM (16.2 mW) to the distal axonal region, the evoked action potential was inhibited. This distal delivery corresponds with the clinical delivery of PBM in the periphery over sites of inflammation and pain suppressing transmission of action potentials to the dorsal horn of the spinal cord. In vivo when tissue injury and inflammation occur, neurons become directly sensitized by the release of inflammatory chemicals such as bradykinin and histamine released locally by cellular disruption and degranulation of mast cells. Nociceptors become activated and pain is experienced. 42 This response is known as peripheral sensitization, the reduction of which, by laser or LED, would have significant effects in reducing nociceptive action potentials and neurogenic inflammation. 43 Also cultured primary DRG neurons–human, rat, or murine–are rarely purely neuronal even when obtained commercially. They contain Schwann cells and peripheral nervous system (PNS) fibroblasts, which are generally overlooked in understanding the interactions between neurons and other cell types. For example, bradykinin in vivo also stimulates Schwann cells, which ensheath all peripheral nerve axons, to release excitatory amino acids. Thus the mechanism of PBM-induced neural inhibition in vivo may well include the effects on molecular exchanges between the neuroglial Schwann cells and the axonal lengths they ensheath. 44
In cell culture studies using rat DRG. 45,46 830, 808, and 650 nm PBM irradiation resulted in reversible neurophysiological changes of significantly decreased mitochondrial membrane potential (MMP) and inhibition of fast axonal flow (FAF). Morphological changes included clustered mitochondria in regions of disrupted microtubule β-tubulin seen morphologically as axonal varicosities—clear indicators of impaired or delayed neural conduction. Varicosities have also been demonstrated in a murine DRG study by Chen et al. using the same wavelength. 47 This disruption would have immediate implications for the transport of ATP-bearing mitochondria from the nerve cell body along the axon where they rely on the fast anterograde transport by the kinesin 1 family motor protein KIF5. 48 In humans, axons may be more than 1 M long. With these fundamental alterations, both morphological and physiological, action potentials could not be generated. This scenario fits the human, animal ex vivo, and cell culture electrophysiological data already discussed, which demonstrate that PBM results in conduction block/neural inhibition.
An interesting possibility would be clinical studies of PBM using the refined technique of axonal excitability studies and threshold tracking. 49,50 This technique could be applied to people with painful conditions to define whether the alteration of function and organization of neural channels such as Na+ of the PNS could also underlie neural inhibition after PBM. This in vivo study would be important as it is well established that the perturbation of the axon–Schwann cell relationship alters both the complex architecture of the axon–Schwann cell internode and node and the distribution and abundance of K+ and Na+ channels in peripheral neuropathies. 51 This technique of axonal excitation could also lead to channel-based clinical trials.
CNS-relevant studies
Like the PNS, the CNS is made up of neurons with neuroglia–oligodendrocytes, astrocytes, and microglia of hemopoietic origin. Although there are studies on effects of PBM on CNS neurons showing formation of varicosities 52 as in the PNS neurons as well as neurophysiological changes, 53 they are not directly relevant to analgesia. However, there are reports of PBM indirectly inducing CNS neural suppression at the spinal cord level that are relevant to analgesia.
A study by Kono et al. 23 recorded dorsal horn potentials (DCPs) evoked by electrical nerve stimulation to a distal portion of exposed sural nerve in anesthetized decerebrate cats. After low-power 632.8 nm, 1 mW PBM irradiation over the sural nerve, proximal to the electrical stimulation, the evoked DCPs were significantly suppressed (25.6% ± 2.5%) by PBM. Similarly, 632.8 nm PBM at 8.5 mW also suppressed single dorsal horn neuron action potentials when applied for 30 min before formalin injection into the skin of raw paw in the receptive field of the spinal cord level of rats. 54 These studies lend weight to the concept that anesthetic effects of PBM downregulate ascending signals from nociceptors to the spinal cord and thence to second-order neurons from the dorsal horn to the higher centers.
Mechanisms of PBM-induced conduction block/neural inhibition
Studies of individual PNS neurons, DRG cultures, and electrophysiology in whole animals indicate that PBM causes significant but reversible changes in neuronal physiology and morphology. The mechanism by which PBM achieves this effect for pain relief and in anesthesia has been somewhat unraveled by experiments on individual neurons as shown by Chow et al. 46 In these studies, cultures of dissociated DRG neurons were exposed to PBM and examined by real-time confocal microscopy and demonstrated morphological changes including significantly decreased MMPs, indicating decreased ATP level, disruption of axonal microtubule β-tubulin polymerization, and significant reduction of FAF consistent with conduction block reported in the electrophysiological studies. The confocal studies also showed significant decrease of movement and clustering of the mitochondrial along the axonal microtubules. All of these changes indicate that neuronal cell body renewal of high 100:1 ratio of ATP to ADP-bearing mitochondria 55 that drives neuronal function and is reliant on FAF will be compromised by PBM. Experiments by Jimbo et al. showed that the stimulatory effect of bradykinin on Aδ and C nociceptors could be inhibited by PBM, leading to decreased neuronal resting potential. This adds to the hypothesis of neural inhibition after PBM. The decrease in membrane potential will prevent an action potential arising in the distal axonal region being transmitted to the synapse with the first-order neurons of the spinal cord and so prevent synaptic neurotransmission at the spinal cord level. It is, therefore, not unexpected that the generation of an action potential will be inhibited as shown in the electrophysiological studies. Importantly, these neuronal changes both morphological and physiological are reversible after PBM.
The neurons responding to PBM include not only the peripheral nociceptors but also their equivalent postganglionic sympathetic neurons. 30 Action potentials from the periphery are transduced across the synapses with the second-order dorsal horn neurons, where the majority of axons cross contralaterally and run in the ascending spinothalamic tracts to synapse with the third-order neurons of the thalamus. These, in turn, synapse with the somatosensory regions of the cerebral cortex and translate into the experience of pain. Neural inhibition would, therefore, prevent synaptic transmission from the skin to the brain. 56 This leads ultimately to suppression of central sensitization 43 and is associated with long-term pain reduction. These pathways emphasize the interconnectivity of the peripheral and central nervous systems (Fig. 2).

Pain-modulatory pathways. This diagram depicts ascending (left) and descending (right) pain pathways that terminate within cortical and subcortical structures and are the basis for pain-modulatory pathways, including the amygdala, hypothalamic nuclei or hypothalamus, thalamus, midbrain PAG, and the RVM. Arrows indicate that PBM can modulate nociceptive signaling in peripheral nerves and modulate central pain pathways.
Summary
The complex phenomenon of pain is an increasing challenge in medicine. The importance of the search for nondrug modalities to relieve pain has been recognized in the International Association for the Study of Pain's Declaration of Montreal 2010. 57 PBM is such a modality. In this review, we have outlined the multiple studies showing that PBM modulates signaling in the PNS, which translates centrally as pain modulation. The combination of the clinical trial data and the experimental data provides a scientific basis for the understanding of one of the mechanisms of PBM in the treatment of pain, which should lead to better acceptance of PBM as a clinical treatment and its acceptance into mainstream medicine.
Footnotes
Author Disclosure Statement
No competing financial interests exist.