
Abstract
Introduction
B
The formation and activation of osteoclasts are dependent on the presence of receptor activator of nuclear factor kappa-B (NF-κB) ligand (RANKL) and macrophage-colony stimulating factor (M-CSF). 4 RANKL plays a critical role in the differentiation and activation of mature osteoclasts derived from monocyte/macrophage hematopoietic lineage cells. The currently known intracellular signaling cascades that are activated after RANKL binds to its receptor include the nuclear factor of activated T cells, mitogen-activated protein kinases (MAPKs), Tumor necrosis factor (TNF) receptor-associated factors, and reactive oxygen species (ROS). 5,6 ROS not only act as signaling molecules but also exert cytotoxic effects such as peroxidizing lipids and causing oxidative damage to DNA and proteins. 7 Cells have several protective mechanisms against these cytotoxic effects. 8 One protection mechanism against cellular stress involves heat shock proteins (HSPs), which are molecular chaperones expressed in cells in response to a variety of stimuli such as temperature, stimulation of membrane-bound receptors by hormones/cytokines, and other chemical factors that help cells cope with oxidative injuries. 9,10 Thus, HSPs are an integral part of mammalian development. 11,12 The mammalian HSPs are oligomeric polypeptides related to α-crystallin. When expressed in mammalian cells, they have a strong protective activity against oxidative stress. 13 HSPs are divided into several families according to the similarity of their amino acid sequences. 14 Some studies reported that HSPs act as molecular chaperones by maintaining stabilizing intracellular proteins following a pathological insult such as ischemia, hyperthermia, and oxidative injury. 14,15
Recently, irradiation with light-emitting diodes (LEDs) has been applied to many clinical fields. 16,17 Some investigators have suggested that LED therapy (LEDT) may modulate wound healing, cell proliferation, differentiation, collagen synthesis, and osteoblast development and bone formation. 18,19 Whelan et al. announced that most of the light photons at wavelengths between 630–800 nm could travel 23 cm through the surface tissue and muscle. 20 Therefore, LEDT could improve not only the superficial wound but also deep injured tissue healing such as bone fracture for the clinical application. In addition, therapy with LEDs instead of a laser has been proposed for conservative treatments while having similar biological and clinical effects. 21 A previous study indicated that 635-nm LED irradiation promoted the scavenging of intracellular ROS, mitigated oxidative stress, and alleviated several pathological conditions. 22,23 However, the mechanism by which LED light modulates oxidative stress during the osteoclastogenic process remains unclear.
The final goal of this study was to investigate the effects on LED irradiation on the signaling pathway in bone marrow-derived macrophage (BMMs) cells in an in vitro RANKL-induced osteoclastogenesis model. Considering the inhibitory mechanism of 635-nm LED irradiation in osteoclastogenesis, the primary objectives were to investigate whether or not 635-nm LED irradiation modulates the RANKL-induced osteoclastogenesis via the HSP signaling pathway in association with alleviation of oxidative stress. In addition, we determined whether or not 635-nm LED irradiation led to inhibit the differentiation and activation of RANKL-induced osteoclast in HSP-silenced cells and analyzed the functional cross talk during osteoclastogenesis.
Materials and Methods
Cell culture and experiment procedure
Eighteen of male BALB/c mice (4 weeks old; Orient Bio Co., SeungNam, Korea) were cared at our facilities according to protocols approved by the Animal Care Committee (CIACUC2014-A0023) of Chosun University. Bone marrow cells were obtained by flushing the femurs and tibia from each mouse with alpha-minimal essential medium (α-MEM; Welgene, Gyeongsan, Korea) containing 10% fetal bovine serum (Gibco, Grand Island, NY) for 1 day. Nonadherent cells were collected and further cultured in the presence of 30 ng/mL M-CSF (R&D Systems, Minneapolis, MN) for 3 days. Cells at this stage were considered BMMs and used as osteoclast precursors. For osteoclast differentiation, BMMs (50,000 cells per cm2) were cultured with M-CSF (30 ng/mL) and RANKL (37.5 ng/mL; R&D Systems) in α-MEM without mix with BMMs from other mice. 18,24 The experimental procedure is presented in Fig. 1.
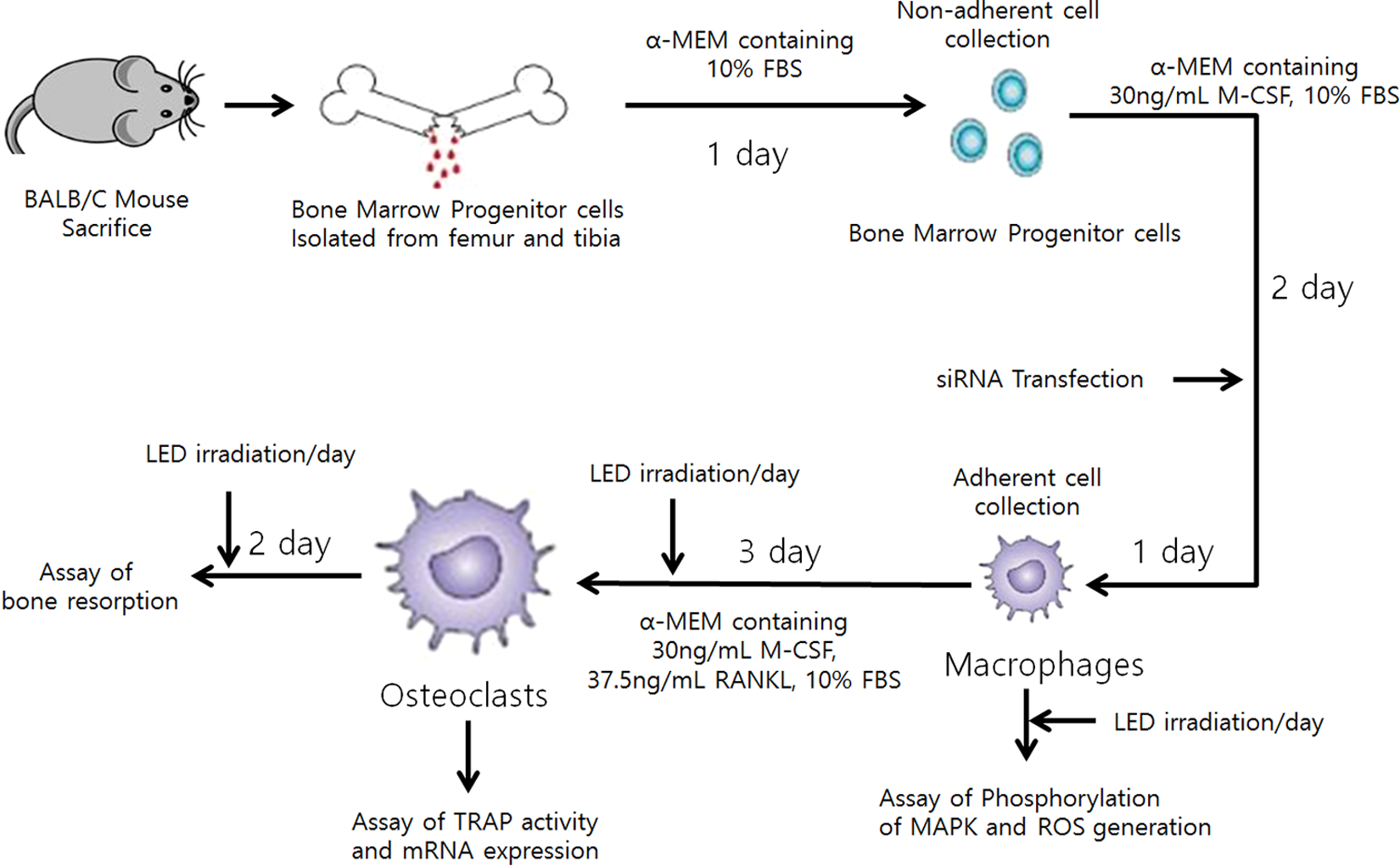
Schematic diagram of BMM isolation and experimental procedure. The assay of bone resorption, TRAP activity, mRNA expression, phosphorylation of MAPK, and ROS generation was carried out in indicated stages, respectively. BMM, bone marrow-derived macrophage; MAPK, mitogen-activated protein kinase; ROS, reactive oxygen species; TRAP, tartrate-resistant acid phosphatase.
Light source and irradiation
An LED irradiation at 635 nm with continuous wave was carried out with a power density of 25 mW/cm2 at surface of light source, 5 mW/cm2 at target cells, and a dose of energy density 18 J/cm2 on the cultured BMMs in the 9-cm Petri dish in the presence of M-CSF and RANKL for 1 h/day during osteoclast differentiation (Fig. 2). The details of the LED irradiation conditions are presented in Table 1. The manufactured LED irradiation tool (Biophoton Co., Gwangju, Korea) using 72 mW of 25 bundle LED ramp (U-Jin LED, Goyang, Korea) was built in a 5% carbon dioxide (CO2) incubator kept at 37°C, and the irradiated area covered to a 9-cm Petri dish. 18,25

An LED irradiation (635 nm) tool kit was built in a CO2-humidified chamber.
LED, light emitting diode.
HSP27 silencing by small interfering RNA
The sequence of HSP27 small interfering RNA (siRNA) corresponds to the mouse HSP27 site (Santa Cruz Biotechnology, Inc., Santa Cruz, CA). A control siRNA (Santa Cruz Biotechnology, Inc.) was used as a control for RNA interference experiments. Cells were plated at a density of 4000 cells per 1.9 cm2, and, 48 h later, cells were transfected with each siRNA (5 nM) for 24 h using the HiPerFect Transfection Reagent (Qiagen, Hilden, Germany). In this experiment, there were four groups: (1) control siRNA treated, (2) control siRNA treated and then 635-nm irradiated, (3) HSP27 siRNA treated, and (4) HSP27 siRNA treated and then 635-nm irradiated.
Western blot analysis
After incubation in the designated reagent for the indicated time with or without LED irradiation for just one time, the media were removed, and the BMMs were washed three times. The lysates were prepared in cold lysis buffer (200 μL, 50 mM Tris-HCl, 150 mM NaCl, 1% NP-40, 150 μg/mL PMSF, 20 μg/mL leupeptin, 2 μg/mL aprotinin, 1 μg/mL pepstatin A, and 0.02% sodium azide, pH 7.5). The cell lysates (30 mg) were separated on a sodium dodecyl sulfate gel and transferred to a polyvinylidene difluoride membrane (Amersham, Piscataway Township, NJ). With a blocking solution (5% skim milk in TBST buffer, pH 7.6 with 8 g/L NaCl, 2.42 g/L Tris-HCl, and 0.1% Tween 20), the membrane was blocked for 30 min and rinsed briefly. The membrane was incubated overnight at 4°C with the anti-HSP27 (1:1000; Cell Signaling Technology, Beverly, MA), anti-HSP70 (1:1000; Cell Signaling Technology), anti-HSP90 (1:1000; Cell Signaling Technology), anti-phospho-extracellular signal-regulated kinase (ERK1/2, 1:500; Invitrogen, Camarillo, CA), anti-phospho-c-Jun-N-terminal kinase (JNK, 1:1000; Invitrogen), anti-phospho-p38 (1:500; Invitrogen), anti-phospho-AKT (1:500; Invitrogen), anti-ERK1/2 (1:500; Invitrogen), anti-JNK (1:1000; Invitrogen), anti-p38 (1:1000; Invitrogen), anti-AKT (1:1000; Invitrogen), and anti-GAPDH (1:3000; Invitrogen) 1” antibodies in 5% nonfat milk. After rinsing with TBST buffer, the membrane was incubated for 1 h with a horseradish peroxidase-conjugated 2” antibody (1:2000; Invitrogen), respectively. Finally, the membrane was washed, immunoreactivity was detected using an enhanced chemiluminescence detection solution (Thermo Fisher Scientific, Inc., Waltham, MA), and the blot measured using ImageJ software (National Institutes of Health, Bethesda, MD).
Tartrate-resistant acid phosphatase staining
To assess the activity of iconoclasts, the osteoclasts were stained for tartrate-resistant phosphatase (TRAP) by incubation with a solution containing naphthol AS-MX phosphate in N,N-dimethylformamide (Sigma-Aldrich, St. Louis, MO) and 5-chloro-4-benzamido-2-methylbenzenediazonium chloride hemi (zinc chloride) salt, as previously described after treating BMMs with RANKL and M-CSF for 3 days with or without three times irradiation. 19 Images of the stained osteoclasts were monitored with a light microscope (Leica Microsystems, Tokyo, Japan). Multinuclear cells were counted manually.
Bone resorption assay
To assess bone resorption pit formation by osteoclast activity, BMMs were cultured on OsteoAssay Plates (Cosmo Bio Co., Ltd, Tokyo, Japan). BMMs were then incubated with M-CSF and RANKL until the mature osteoclasts resorbed the calcium phosphate-coated film. After 5 days with or without five times irradiation in culture, the attached cells were dissolved in sodium hypochlorite (5%), and images of the resorption pits were acquired with a light microscope (Leica Microsystems). The ratio of the resorbed area was determined using ImageJ software.
Total RNA isolation and quantitative real-time polymerase chain reaction
After treating BMMs with RANKL and M-CSF for 3 days with three times irradiation or not, total RNA was isolated from cells using the TRIzol reagent (Thermo Fisher Scientific, Inc.) according to the manufacturer's instructions to assess the related mRNA expression. The cDNA was synthesized by RNA (1 mg) and reverse transcription polymerase chain reaction (PCR) kit (Thermo Fisher scientific, Inc.). The cDNA was amplified using the primer sets listed in Table 2, and quantitative real-time PCR (qRT-PCR) was conducted using an Exicycler™ 96 (Bioneer Co., Daejeon, Korea) with a 20 μL reaction mixture containing 10 μL SYBR Green Premix (Thermo Fisher scientific, Inc.), forward primer (10 pmol), reverse primer (10 pmol), and cDNA (1 μg). The amplification parameters consisted of an initial denaturation step for 5 min at 95°C followed by 40 cycles of three-step PCR (denaturation for 1 min at 95°C, annealing for 30 sec at 60°C, and extension for 1 min at 72°C). Relative gene expression levels were normalized to that of the GAPDH gene, Gapdh.
DC-STAMP, dendrocyte expressed seven-transmembrane protein; OSCAR, osteoclast-associated immunoglobulin-like receptor; TRAP, tartrate-resistant acid phosphatase.
Detection of ROS formation
To assess the intracellular ROS generation, the 2′,7′-dichlorodihydrofluorescein diacetate (H2DCF-DA; Thermo Fisher Scientific, Inc.) method was carried out as described previously. 19 Cells were grown on four-well chamber slides (Thermo Fisher Scientific, Inc.), treated with RANKL, irradiated with LED for 1 h or not, and then immediately incubated with DCF-DA (10 mM) for 20 min. The cells were then washed with PBS and DCF intensity was detected using a confocal microscope (Leica Microsystems) set at excitation and emission wavelengths of 488 and 525 nm, respectively.
Statistical analysis
All experiments were carried out in triplicate. Data are expressed as the mean ± SD. The results were subjected to analysis of variance (ANOVA) using the SPSS program (SPSS V12.0K; SPSS, Inc., Chicago, IL) to analyze the differences and multiple comparisons were performed with Tukey's test. p < 0.05 was considered to be statistically significant.
Results
Effect of LED irradiation on HSP expression
To identify the HSPs affected by LED irradiation, LED-irradiated BMMs were analyzed using western blotting (Fig. 3a). HSP27 expression was significantly increased by LED irradiation, whereas that of HSP70 was only slightly increased. However, LED irradiation decreased HSP90 expression. Therefore, HSP27 was selected for subsequent experiments.
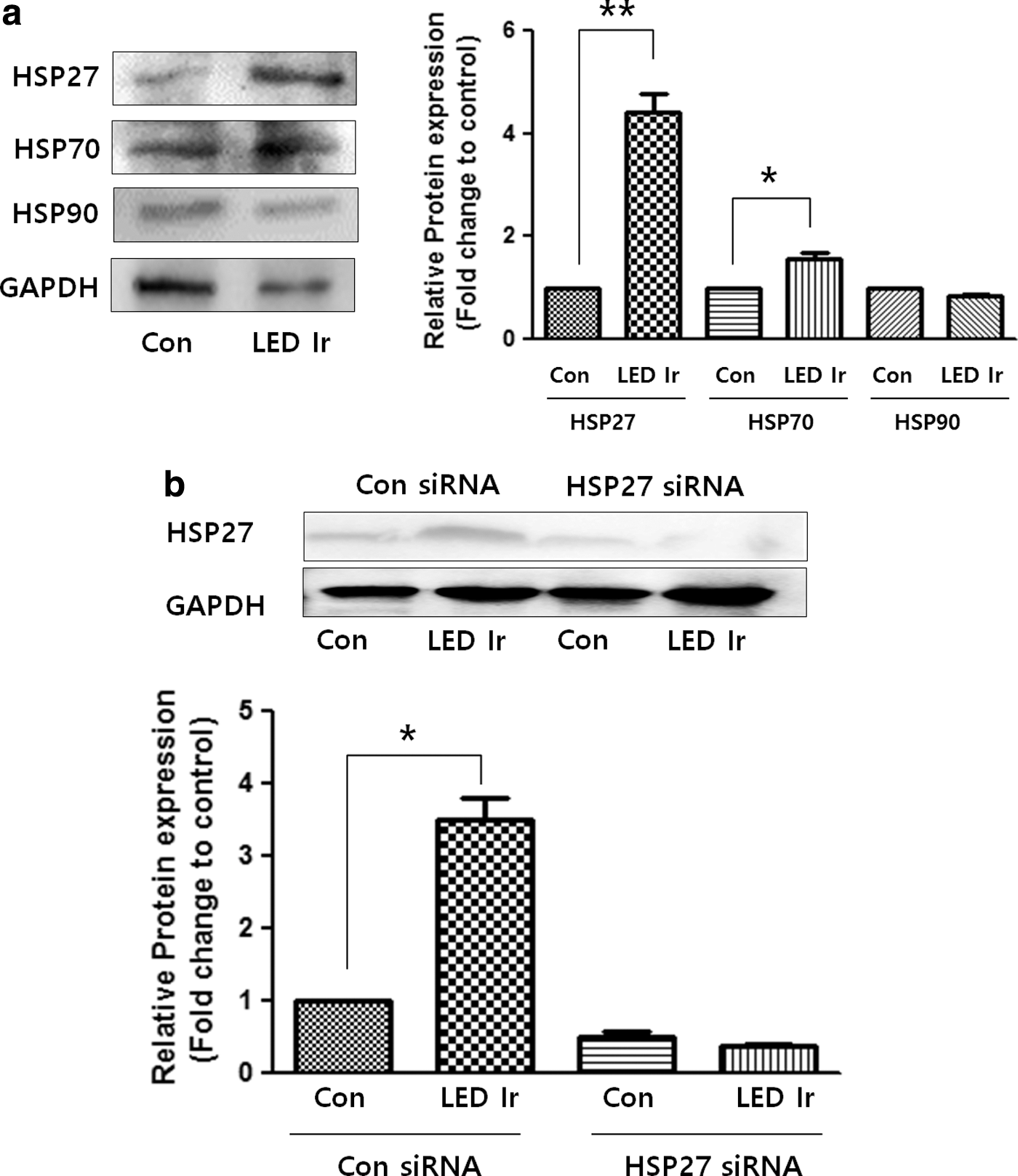
Effect of LED irradiation on HSP expression in BMMs.
To clarify whether HSP27 activation is involved in the effects of LED irradiation, HSP27 was knocked down in BMMs by siRNA. The western blot results confirmed the low expression levels of HSP27 in HSP27 siRNA-transfected cells compared with the controls. LED irradiation only induced an increase in HSP27 expression in the control siRNA-transfected cells (Fig. 3b).
Effect of LED irradiation on osteoclastic differentiation in HSP27-silenced BMMs
To evaluate the effects of LED irradiation on osteoclastogenesis, the TRAP assay was performed on control siRNA- or HSP27 siRNA-transfected primary BMMs with or without LED irradiation in the presence of RANKL (37.5 μg/mL; Fig. 4a, b). Nonirradiated control BMMs differentiated into mature TRAP-positive multinucleated osteoclasts. However, the number of mature osteoclasts was significantly reduced by LED irradiation of BMMs. Treatment of HSP27 siRNA-transfected cells with 635-nm LED irradiation had no effect on TRAP-positive cells. These results indicate that 635-nm irradiation decreases osteoclastogenic differentiation in a manner that is dependent on HSP27 expression.
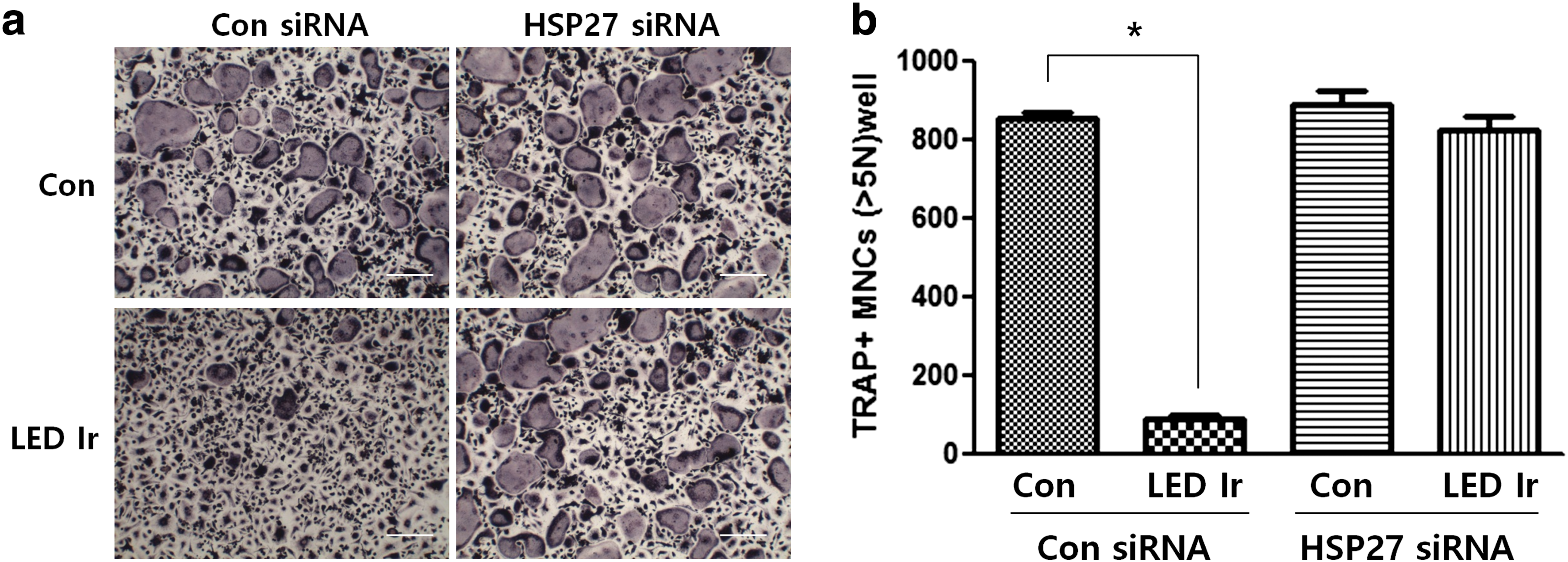
Effect of LED irradiation on HSP27-silenced osteoclast differentiation.
Effect of LED irradiation on osteoclastic bone-resorbing activity in HSP27-silenced BMMs
To determine whether LED irradiation adversely affects osteoclast function, we conducted calcium phosphate resorption assay. LED irradiation resulted in a reduced resorption area, from 49.5 ± 1.4 to 7.9 ± 3.9 mm2, in RANKL-induced BMMs (Fig. 5a, b). However, LED irradiation had no significant effect on bone mineral resorption pit formation (54.4 ± 4.8 to 47.1 ± 0.9 mm2), indicating that LED irradiation inhibits bone resorption by mature osteoclasts in an HSP27-dependent manner.
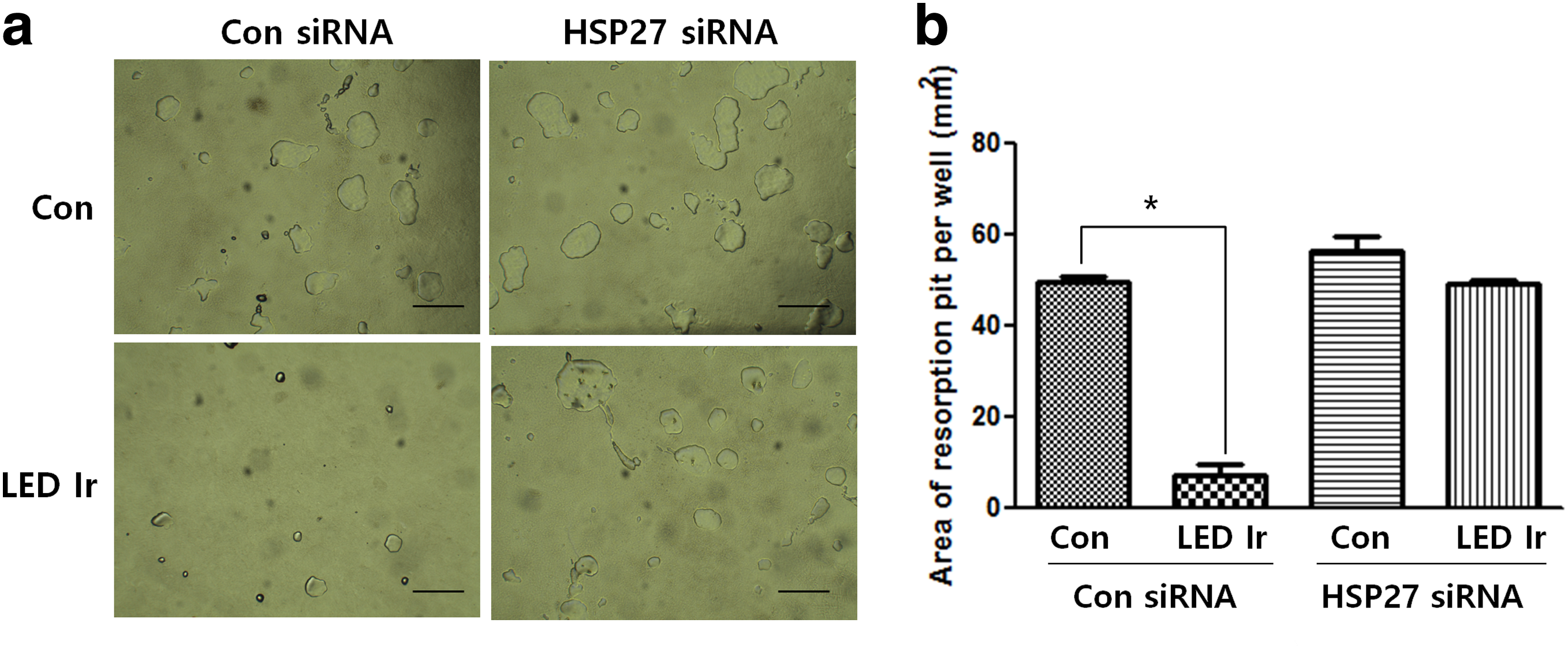
Effect of LED irradiation on bone resorption in HSP27-silenced BMMs.
Modulation of osteoclastogenesis-associated mRNA expression in HSP-silenced BMMs by LED irradiation
To further explore the influence of LED irradiation on RANKL-stimulated osteoclastogenesis, qRT-PCR was performed to examine the mRNA expression of various osteoclast differentiation markers [i.e., c-fos, NFATc1, ATP6V0d2, dendrocyte expressed seven-transmembrane protein (DC-STAMP0), TRAP, and osteoclast-associated immunoglobulin-like receptor (OSCAR)] in control siRNA- or HSP27 siRNA-transfected primary BMMs with or without LED irradiation in the presence of RANKL (37.5 μg/mL) after 3 days (Fig. 6). The mRNA expression of c-fos, Nfatc1, and Dc-stamp in BMMs transfected with control siRNA was significantly downregulated by LED irradiation, whereas of that in HSP27 siRNA-transfected BMMs was slightly decreased. The expression of ATP6V0d2, Trap, and Oscar was slightly decreased by LED irradiation in both control siRNA- and HSP27 siRNA-transfected BMMs.
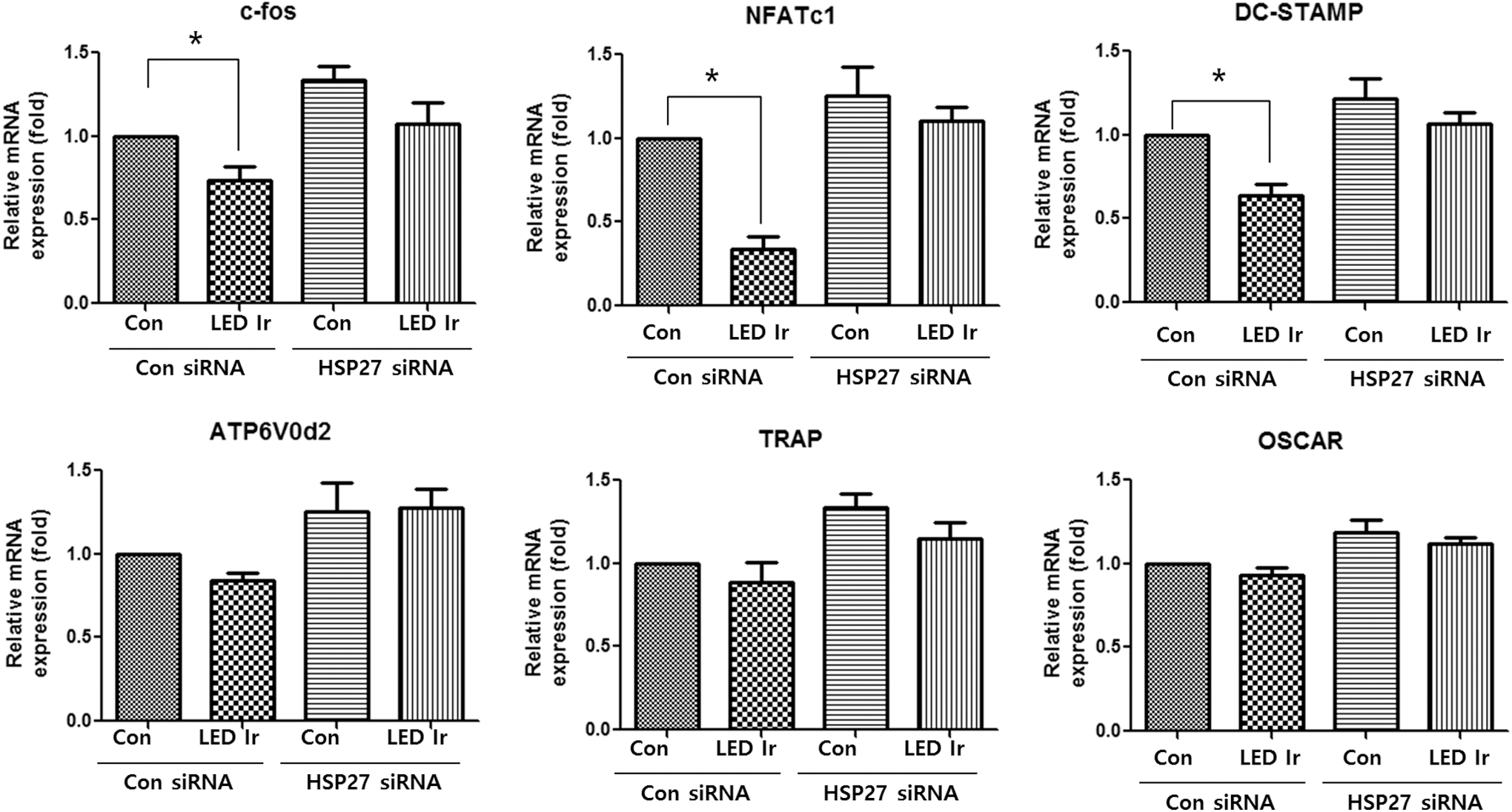
Effect of LED irradiation on osteoclast gene expression in HSP27-silenced BMMs. mRNA expression levels of Trap, c-fos, ATP6V0d2, Dc-stamp, Nfatc1, and Oscar in HSP27 siRNA-induced BMMs are shown. Bars and error bars indicate mean ratios and the SD (n = 3 independent experiments; *p < 0.05 vs. the indicated condition). Dc-stamp, dendrocyte expressed seven-transmembrane protein; Oscar, osteoclast-associated immunoglobulin-like receptor.
Inhibitory action of LED irradiation on MAPK phosphorylation in HSP-silenced BMMs
We examined the influence of 635-nm irradiation on MAPK activity in HSP27-silenced BMMs by measuring the phosphorylation levels of ERK, JNK, AKT, and p38 (Fig. 7). Exposure to LED light significantly diminished AKT and p38 phosphorylation in control siRNA-transfected BMMs in the presence of RANKL. However, no change in AKT and p38 phosphorylation was observed in HSP27 siRNA-transfected BMMS in the presence of RANKL with LED irradiation. Moreover, no differences in JNK and ERK phosphorylation were observed between control siRNA- and HSP27 siRNA-transfected BMMs. These results suggest that LED irradiation affects the phosphorylation of AKT and p38 in an HSP27-dependent manner.
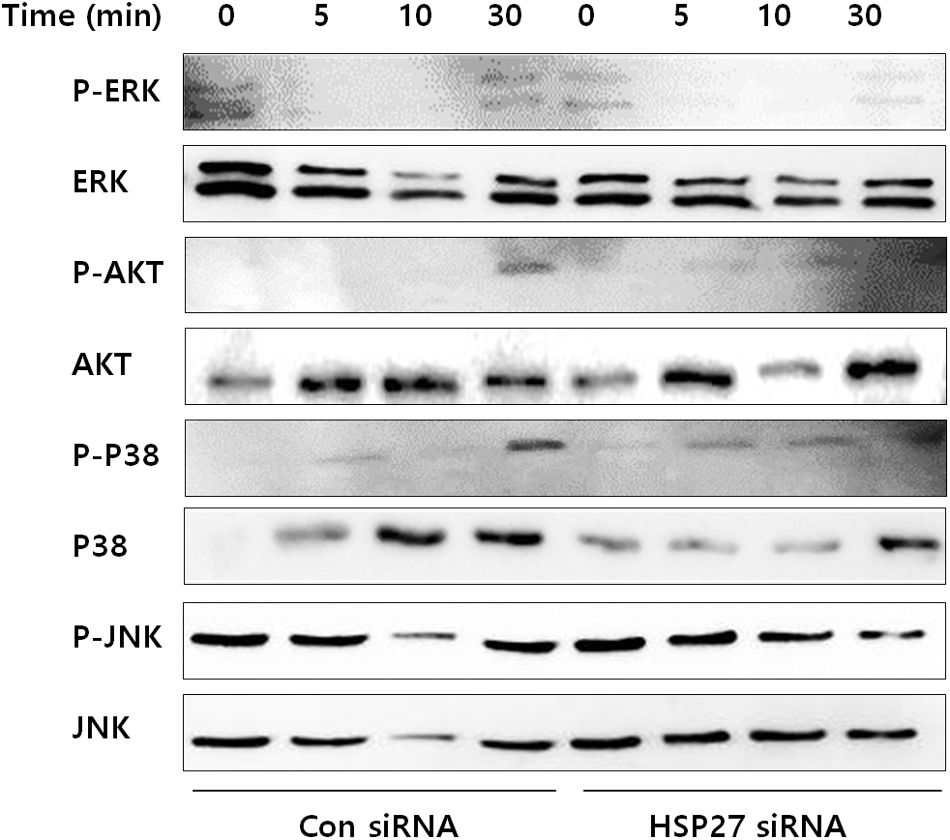
Effect of LED irradiation on the phosphorylation levels of AKT, ERK, p38, and JNK in HSP27-silenced BMMs. Similar results were obtained in three independent experiments. ERK, extracellular signal-regulated kinase; JNK, c-Jun-N-terminal kinase.
Effect of 635-nm irradiation on ROS generation in HSP27 siRNA-induced BMMs
The confocal microscopy results indicated that 635-nm irradiation led to a decrease in DCF, indicating the production of ROS in control siRNA-transfected BMMs in the presence of RANKL (Fig. 8a). However, HSP27-siRNA transfection resulted in an increase in DCF in BMMs. These results indicated that 635-nm irradiation did not affect the intracellular ROS levels in HSP27-silenced BMMs.

Effect of LED irradiation on ROS production in HSP27-silenced BMMs.
Discussion
LEDT has gained considerable attention for its ability to regulate bone remodeling properties by preventing bone loss in osteoporosis and other resorptive bone diseases. 26 –28 LEDT positively affects bone metabolism by increasing bone formation and suppressing bone resorption. 29 Considering the clinical uses of LED irradiation, LEDT was considered to treat the not deeper tissues but superficial wound. However, Whelan et al. announced that most of the light between 630–800 nm could travel into the muscle and bone. 20 Therefore, if the BMMs were exposed to over the 5 mW/cm2 (a fairly low dose of power density) as in our study, the penetrated light into the bone marrow hypothetically should inhibit osteoclast activity. In addition, in a previous study, 635-nm LEDT markedly reduced intracellular ROS generation and NF-κB activation, led to the suppression of TRAP expression and activation, and ultimately decreased osteoclastogenesis and disorganization of the cytoskeleton of osteoclast cells in RANKL-induced BMMs. 18,30 Another report indicated that He-Ne laser irradiation (632.8 nm) around 635-nm wavelength led the promotion of bone repair in the cortical bone of the rat tibia in vivo study. 31,32 In the present study, 635-nm LED irradiation significantly suppressed TRAP activity in RANKL-induced BMMs. Moreover, 635-nm irradiation suppressed the activation of osteoclasts, resulting in a decreased resorption pit area. To date, only a few studies have addressed the effect of light therapy on osteoclast differentiation or bone resorption, and the signaling pathway by which LED irradiation regulates osteoclastogenic responses has not been described until now. Therefore, an additional goal of this study was to explore the signaling pathways involved in the response to 635-nm LED irradiation and its underlying inhibitory effect on osteoclastogenesis.
HSP27 has been reported to be the factor affected by 635-nm irradiation, leading to the inhibition of RANKL-induced osteoclastogenic responses. HSP27 is an important modulator of intracellular ROS generation, and its response is affected by various intra/extracellular stimuli such as oxidative stress. 33 In a previous study, 635-nm LED irradiation was shown to attenuate LPS-induced NF-κB activation in vitro via HSP27, leading to the inhibition of intracellular ROS generation as well as the phosphorylation of IKK and IκB. 34 Various lines of evidence suggest that HSPs protect cells against the deleterious effects of ROS and contribute to the maintenance of redox homeostasis and mitochondrial stability. HSP25/27 synthesis can be induced by different conditions, including heat shock and other stress conditions, nerve injury, and differentiation. 14 The effects of HSP27 seem to be determined by its expression level. Our results show that 635-nm irradiation led to a significant increase in HSP27 expression among the HSPs expressed in BMMs, indicating that HSP27 could play a pivotal role in mediating the cellular response to LED irradiation. Because HSP27 silencing in BMMs leads to ROS generation in the presence of RANKL, increased HSP27 expression levels following LED irradiation play a critical role in the reduction of oxidative stress by reducing ROS levels. ROS are constantly produced under physiological conditions, and these oxygen side products, which are induced by oxidative stress, are potent regulators of cellular activities such as proliferation and differentiation. 35 In addition, exogenous ROS induce osteoclastogenesis; thus, oxidative stress affects the regulation of osteoclastogenesis both intercellularly and extracellularly. Therefore, oxidative stress due to increased ROS production is considered to play a major role in the differentiation of osteoclasts, and LED irradiation could inhibit ROS generation through HSP27.
To identify the signaling pathway activated in response to 635-nm LED irradiation during osteoclastogenesis, the activation of MAPK phosphorylation by 635-nm LED irradiation was assessed in HSP27-silenced BMMs in the presence of RANKL. MAPKs (JNK, ERK, and p38) are activated by RANKL stimulation and are associated with osteoclastogenesis. 36 p38 is important in the early stages of osteoclast generation because it regulates the microphthalmia-associated transcription factor, whereas dominant-negative JNK prevents RANKL-induced osteoclastogenesis. 37 Meanwhile, ERK is known to induce c-Fos for osteoclastogenesis, and inhibition of ERK reduces osteoclast formation. 38 In a previous study, we evaluated the effects of 635-nm LED irradiation on the activation of these MAPKs and found that LED inhibited the phosphorylation of p38 and ERK in RANKL-induced BMMs. 18 However, in the present study, 635-nm LED irradiation did not affect p38 phosphorylation in HSP27-silenced BMMs, indicating that LED irradiation decreases p38 phosphorylation in an HSP27-dependent manner.
HSP27 silencing altered the expression of genes associated with osteoclast differentiation and activation. Monocyte/macrophage precursors develop into functionally active mature osteoclasts via a process involving commitment, differentiation, fusion, and bone resorption, which is under gene regulation. 39 In our qRT-PCR analysis, the mRNA expression levels of c-fos, Nfatc1, and Dc-stamp were markedly decreased by LED irradiation in RANKL-induced BMMs, whereas the expression levels of the osteoclast-associated genes, ATP6v0d2, Trap, and Oscar, were slightly decreased in HSP27-silenced BMMs. This finding indicates that 635-nm irradiation could activate the expression of genes involved in the early stage of osteoclast differentiation such as c-fos, Nfatc1, and Dc-stamp through HSP27 and that HSP27 is essential for downregulating the expression of genes involved in osteoclast differentiation induced by 635-nm irradiation. Also, further in vivo studies are required to elucidate the mechanisms in HSP27 knockout mice, by which signaling cascades are involved in the inhibitory effects of 635-nm LED irradiation on the osteoclast differentiation and activation.
Conclusions
In conclusion, we report the novel finding that 635-nm irradiation attenuates RANKL-induced osteoclastogenesis in BMMs via HSP27, which leads to the inhibition of intracellular ROS generation; p38 and AKT phosphorylation; c-fos, Nfatc1, and Dc-stamp mRNA expression; TRAP activation; and osteoclastic bone resorption. These results indicate that HSP27 plays a crucial role in the decrease in osteoclastic differentiation caused by 635-nm irradiation by regulating RANKL-induced severe osteoclastogenesis. Thus, LEDT by targeting HSP27 could be applied to conservative treatment for deep tissue healing such as osteoporotic bone fracture as clinical usage.
Footnotes
Author Disclosure Statement
No competing financial interests exist.