
Abstract
Introduction
P
The clinical problem of artificial insemination is challenging and, recently, many studies have shown that PBM can be used in this field. 8,11,12
Many studies have shown the potential effect of PBM by low-power laser irradiation on spermatozoa in increasing their motility and raising the adenosine triphosphate (ATP) amount in the cells. 8,11,13 Subsequently, there is evidence base that the effect of He-Ne laser irradiation (632.8 nm) on active living human sperms can improve their motility and speed. 14 –16 Also, it has been shown that PBM with light-emitting diodes has improved the sperm motility in men with asthenozoospermia. 17
Shahar et al. 18 showed in their study a significant increase in the hyperactivated motility of human sperm, irradiated with 400–800 nm light by a mechanism mediated by reactive oxygen species (ROS) production and activation of the intracellular Ca2+ influx. A recent study 19 reported that He-Ne laser irradiation has a different action on PBM of turkey, chicken, and pheasant spermatozoa; the suggestion was a possibility to use PBM for restoration of motility of cryopreserved avian spermatozoa.
There are only a few studies that demonstrate the effect of PBM on oocytes. 11 Soares et al. 20 have used the He-Ne laser (632.8 nm) to irradiate bovine oocytes. These authors have found in their studies that PBM is capable of modulating events in granulosa cells, which may lead to changes in oocytes. In contrast, Ocana-Quero et al. 21 have used low-power laser settings to treat immature oocytes. They found in their studies that He-Ne laser irradiation with doses of 0.40 and 2 J/cm2 has had a significant negative effect on the maturation process and has caused nuclear damage (see also Karu's review 12 ).
A novel hand piece for PBM applications has been recently introduced to the market. This instrument, “named AB2799” (Doctor Smile–LAMBDA SpA–Vicenza, Italy), is a flat-top hand piece, which, in contrast to a hand piece with Gaussian profile output, provides a homogeneous irradiation over a 1 cm2 surface. It has the same irradiation area (spot size) from contact to 105 cm distance from the target tissue. Thus, it provides a homogenous energy density by using relatively high-power densities in less time and with less risk of thermal damage. 2,7
Our previous studies have shown that irradiation with an 808 nm diode laser with 64 J/cm2 of fluence and power of 1 W in continuous wave (CW), with a flat top hand piece (AB2799), induced PBM on Paramecium primaurelia (Ciliophora, Protozoa) and mammalian mitochondrial activities without generating thermal damage. 2,3,7,22,23
Further, the same PBM has stimulated the Paramecium swimming speed and the food vacuole formation, to suggest accelerated cilia beating as a consequence of increased Ca2+ influxes. 3 As the cilia structure, genes, proteins, and functions are highly conserved throughout evolution from protozoa to humans 24 and because of increased energy observed in the irradiated Paramecium cells, the 808 nm with 64 J/cm2 with 1 W (CW) parameter applies as proven therapy for gamete PBM.
However, if interest in the contribution of PBM effects on the artificial insemination of animals and human has increased, 8 the knowledge of the effect of PBM on the embryo/larval development by irradiated gametes or irradiated embryos has not been clearly understood.
This study investigated the PBM effect of the diode laser (808 nm laser light) irradiation on a bioethical compatible model of embryogenesis, by using higher fluence (64 J/cm2–1 W or 192 J/cm2–3 W) and AB2799 flat top hand piece.
Many studies have demonstrated the reliability of the Mediterranean Sea urchin Paracentrotus lividus for studying the mechanisms driving fertilization, development, and differentiation in higher organisms, including mammalians. 25 –28 Therefore, the authors chose this model.
The sea urchin has more than 70% genome homology and metabolic similarity to the higher vertebrates, including humans. 29 Such a model represents a number of other features, such as high number of eggs, transparent envelope, synchronous development, and the normal development, which makes it an optimal and suitable nonsentient model organism. 30 –32 In this regard it fully respects the 3Rs objectives, Reduction, Refinement, Replacement of animal experiments, 33 which would eventually lead to a reduction in vertebrate use for toxicity testing. It is one of the few organisms that is considered as a promising model by the European Agency for Alternative Models. 30,34
Materials and Methods
Experimental condition of irradiance
The irradiation was performed by using an 808 nm diode laser (Doctor Smile–LAMBDA SpA–Vicenza, Italy) with flat top hand piece (AB2799, Doctor Smile–LAMBDA SpA–Vicenza, Italy) (Table 1). A power meter was used to check the real power output of the laser (PM10 Coherent- Ca-Usa).
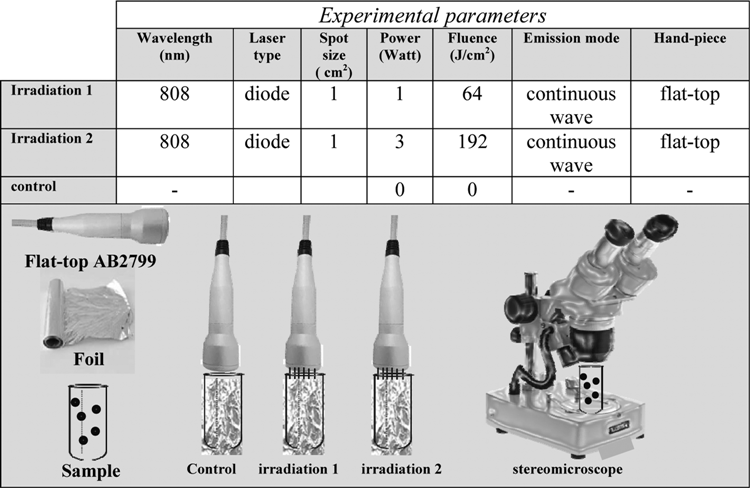
As the flat-top hand piece provides homogeneous irradiation over a 1 cm2 surface, 23 the authors performed the irradiation into an Eppendorf tube or a chamber of a multiwell plate, which is about 1.10 cm in diameter. During the process of irradiation or nonirradiation (control), the flat-top hand piece was kept in contact and perpendicular to the surfaces of the tube and the chamber, which were wrapped with a foil; the surface was not covered with foil (Table 1: control, irradiation 1, and irradiation 2).
In our experiments, the power settings were 1 and 3 W, respectively (Table 1). The irradiated cells were exposed to fluence of 64 J/cm2 using power of 1 W in a CW (Table 1, irradiation 1) or to a higher fluence of 192 J/cm2 by using a higher power setting of 3 W in a CW (Table 1, irradiation 2). The irradiated samples and the control sample were monitored by stereomicroscope (Table 1) as reported in the following subsection of Materials and Methods.
Sea urchin life cycle and experimental setup
The P. lividus life cycle includes the planktonic (embryo and larva) and the benthic (adult specimen) phases. The gametes from the adult sea urchin rapidly produce zygotes that become blastula within about 12 h postfertilization (hpf). The embryos reach their late gastrula stage corresponding to about 24 hpf, while at 48, 72, and 96 hpf they reach early pluteus, pluteus, and late pluteus, respectively. 35 These larvae grow by planktonic feeding and ultimately undergo metamorphosis and grow into adults. 36 In this study, sea urchin gametes, zygotes, embryos, and larvae (up to 96 hpf) were irradiated as summarized in Fig. 1. The fertilization rate and the early developmental stages were investigated. The experiment was stopped at 96 hpf, as after that time, larvae starvation may become a limiting factor. 37
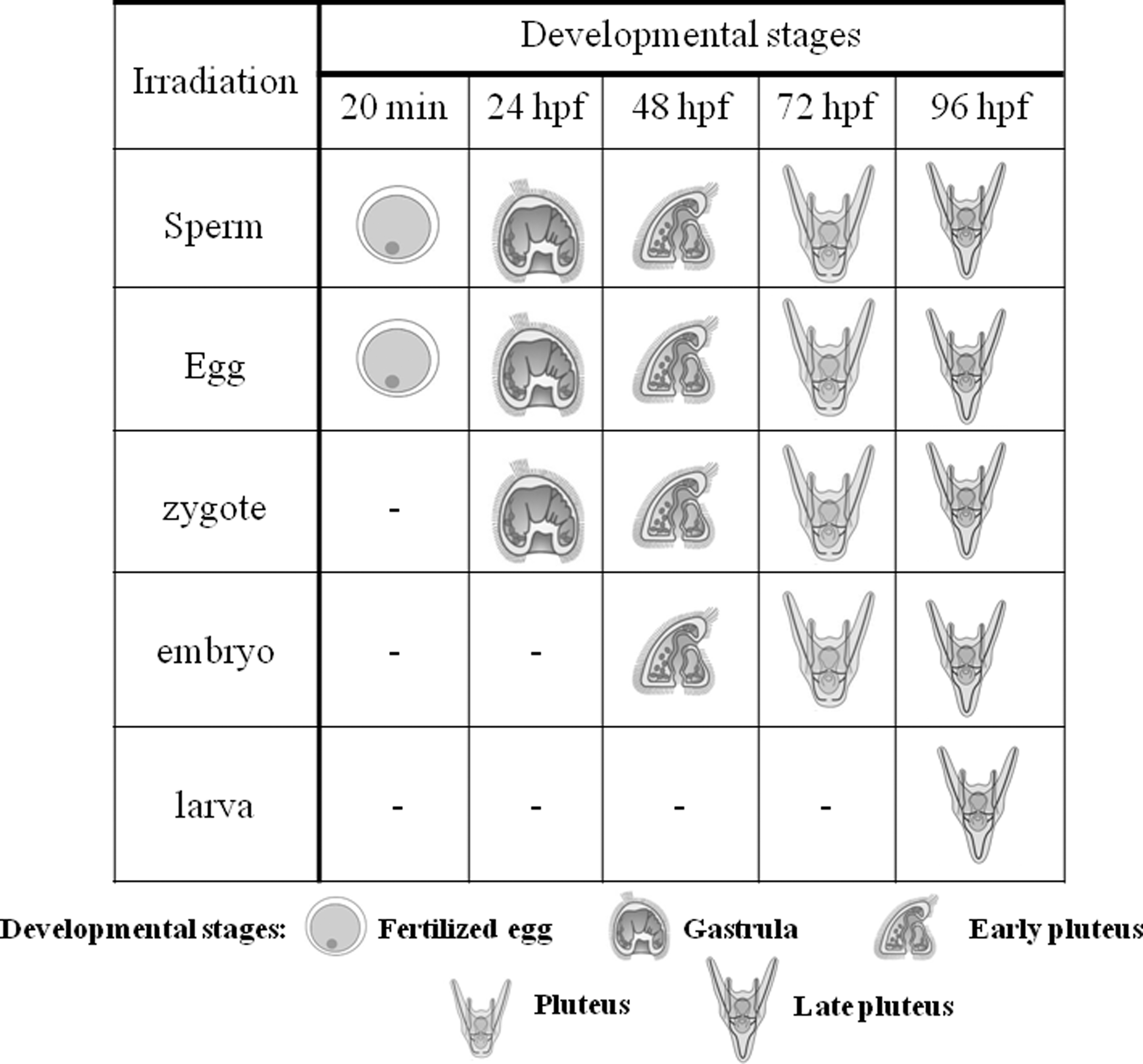
Shows the timing of the developmental and stage-specific laser exposure experiments. For the experiments, sperm, egg, zygote, 24-h embryo, and 72-h larva were irradiated. After the exposure, the fertilization success and the developmental stages defined as gastrula (24 hpf), early pluteus (48 hpf), pluteus (72 hpf), and late pluteus (96 hpf) were investigated. hpf, hours postfertilization.
Sea urchin sampling and gamete emission
The adult specimens of P. lividus were collected along the coast of Genoa (Italy). The mature specimens were induced to spawn by intraoral injection of 1 mL of 0.5 M KCl solution. 38,39
Sperm irradiation
The sperm of three different specimens was mixed. The experiments were carried out in triplicate. Ten microliters of “dry” sperm was diluted in 1 mL of fresh sea water (FSW). For the test, 5 μL of diluted sperm was added to 0.5 mL of FSW into the Eppendorf vials and irradiated at the two different power settings: 1 and 3 W. Negative controls (nonirradiated sperm) were performed. Ten microliters of the sperm from each vial was added to multiwell capsules containing ∼300 eggs/mL in 10 mL of FSW. Under normal conditions, the egg activation process occurs within 60 sec of the addition of sperm. The test was performed following the methods that were suggested by Annamaria et al. 40 The 2 mL of water containing ∼600 zygotes was collected from each well 20 min after sperm was added. This process was observed under the microscope (Zeiss) to check the successful percentage of fertilization. 41 The remaining zygotes were left to develop at 18°C in a thermostatic room and were monitored at 24, 48, 72, and 96 h, corresponding to gastrula, early pluteus, pluteus, and late pluteus stage, respectively. These stages were then fixed in cold pure methanol and 20% polyethylene glycol (Sigma). The state of embryos and larvae was checked under a light microscope (Zeiss) and classified according to the morphology and synchronicity of development compared to the controls, according to the studies by Gambardella et al. 42 and Mesarič et al. 43 In detail, the following morphological markers were considered: (1) developed embryos and larvae; and (2) undeveloped embryos and larvae. The first category included organisms that showed normal development, with well-structured archenteron and migratory cells infiltrated into the coelom, while the undeveloped organisms were defective embryos and/or larva, with typical signs of asymmetrical migration of primary mesenchyme cells, as recorded by Gambardella et al. 44 This latter category was represented by irregular embryo shapes showing loss of cell–cell adhesion at the gastrula surface, embryos, and had not reached the pluteus stage, as well as anomalous larvae characterized by the lack of skeleton or showing skeletal rod damages and defects.
Therefore, the state of the embryos and larvae was checked by analyzing such morphological markers (i.e., developed and undeveloped gastrulae and plutei).
Egg irradiation
The 300 eggs from three different specimens were mixed in an Eppendorf vial, which was diluted with 0.5 mL of FSW and then irradiated by two different average power settings: 1 and 3 W. The experiments were carried out in triplicate. The negative control consisted of a group of nonirradiated eggs. The eggs from each vial were added to multiwell capsules containing 9.5 mL FSW, and then, 5 μL of nonirradiated diluted sperm was added. The fertilization's ability and the early developmental stages were monitored after 20 min, 24, 48, 72, and 96 h, as described in the previous paragraph.
Zygote, embryo, and larval irradiation
The 300 zygotes were placed into an Eppendorf vial, which was diluted with 0.5 mL of FSW and then irradiated by two different power setting; 1 and 3 W. The experiments were carried out in triplicate. The negative control consisted of nonirradiated zygotes. Zygotes from each vial were added to multiwell capsules containing 9.5 mL FSW. The early developmental stages of this experiment were monitored after 48, 72, and 96 h, as described in the previous paragraph.
The same procedure was used for irradiating 24-h embryos and 72-h larvae, corresponding to the gastrula and pluteus stages, respectively.
Statistical data analysis
All experiments were repeated three times during a nominal breeding phase (approximately from the end of spring to early Autumn) and were carried out in triplicate (each sample included 300 samples per replicate). All data are expressed as mean ± standard error (SE). The significant differences between the controls and the irradiated samples were determined by using one-way ANOVA (Bonferroni nonparametric post hoc tests). A p value lower than 0.05 (p < 0.05) was considered significant. The significant differences among the controls and the irradiated samples are reported in the figure legends.
Results
Fertilization rate
The egg fertilization by the P. lividus sperm was demonstrated by the cortical reaction of the egg that occurred in response to the first sperm–egg contact. In these experiments, the controls showed 99% ± 1% of fertilized eggs (Fig. 2). The fertilization ability was not affected by the sperm/egg irradiation (Fig. 2). This was observed at an early development of the zygotes after sperm was irradiated with 1 and 3 W in comparison to the controls. This percentage was significantly different from the control (p < 0.05). The same early development of the zygotes was found in eggs irradiated with 3 W, although in this case it was not significantly different from the control.
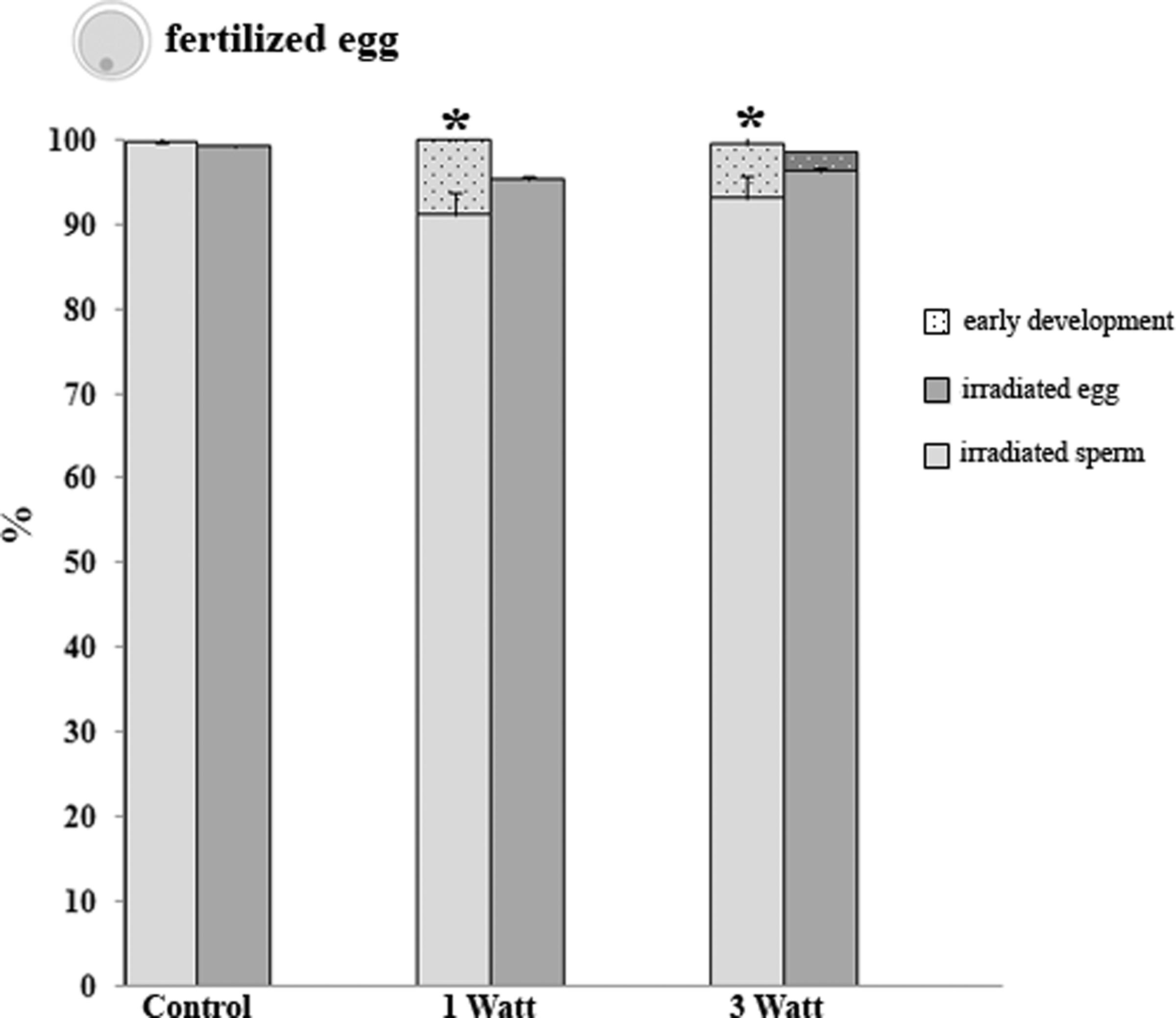
Shows the percentage of fertilization (full bars) and early development (tiny dotted bars) from the irradiated sperm (light gray bars) and the irradiated egg (gray bars). Data are mean ± standard error (M ± SE) of three replicates. *p < 0.05.
Gastrula (24 hpf)
The gastrulae were observed at 24 hpf from irradiated gametes and zygotes. This presented as a low number of undeveloped embryos (<10%), which showed that the laser exposure at 1 and 3 W did not cause damage to the offspring. No significant differences were observed in the percentage of both developed and undeveloped embryos compared with the control samples (Fig. 3).
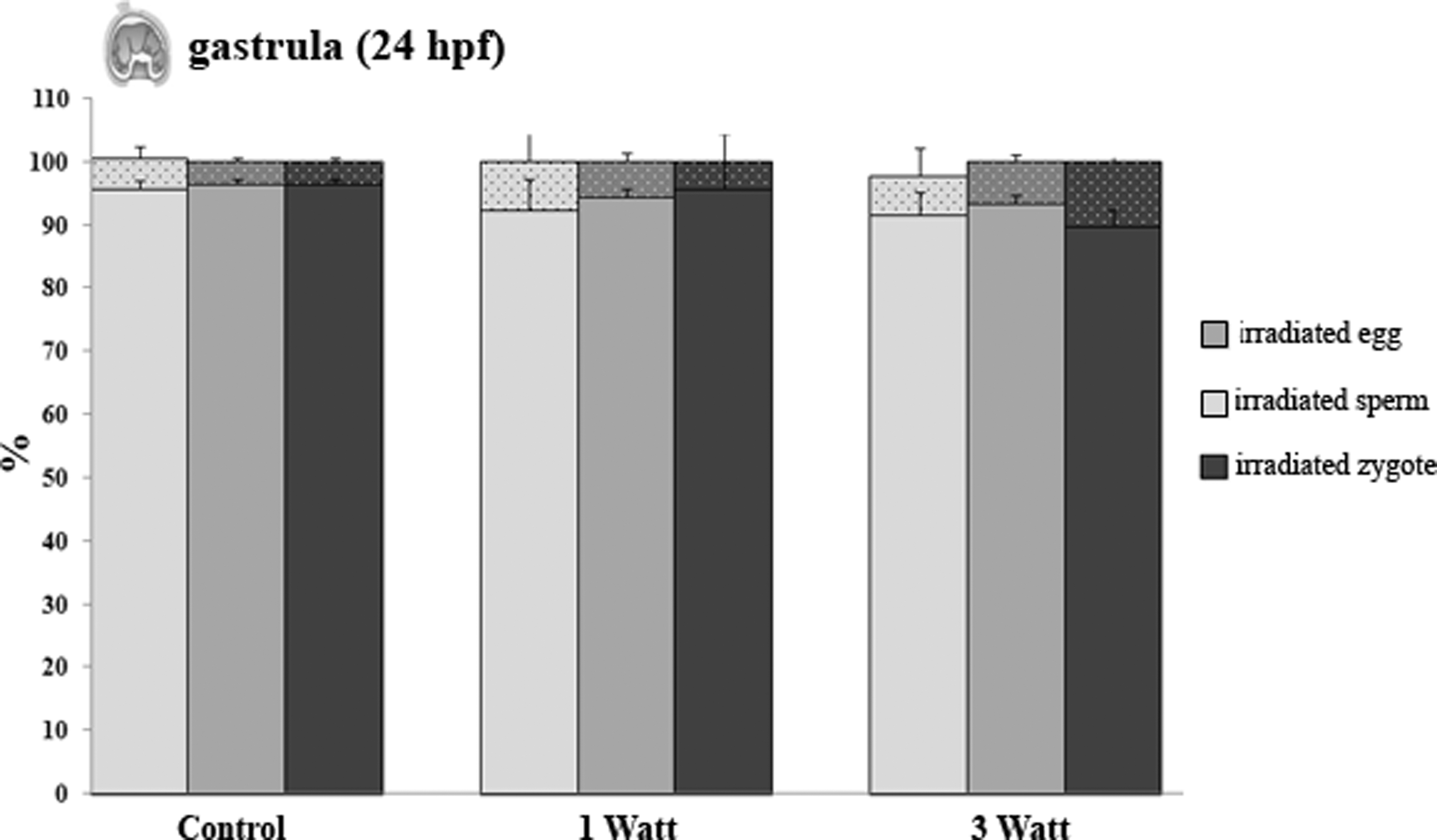
Shows the percentage of the developed (full bars) and not developed (tiny dotted bars) embryos from the irradiated sperm (light gray bars), egg (gray bars), and zygote (dark gray bars). Data are mean ± standard error (M ± SE) of three replicates.
Pluteus (larva) developmental stages
The larvae were monitored at 48, 72, and 96 h, corresponded to the pluteus stage of controls. This has presented in different types of anomalous development, including damage and defects of the skeletal rods and delayed specimens (Figs. 4 and 5). The delayed specimens included all the embryos that had not reached the pluteus stage.
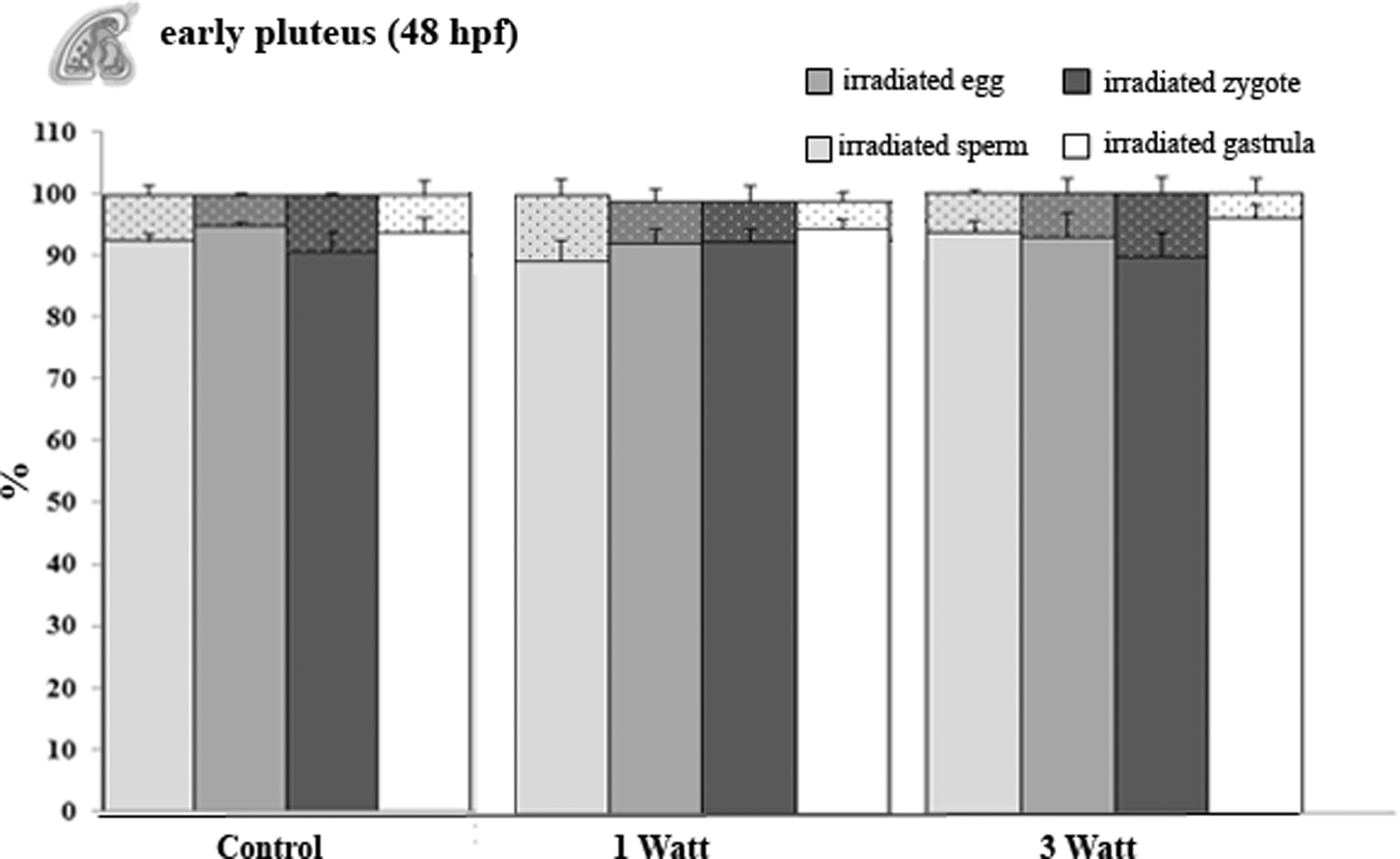
Shows the percentage of developed (full bars) and anomalous development (tiny dotted bars) from the irradiated sperm (light gray bars), the egg (gray bars), the zygote (dark gray bars), and the gastrula (white bars). Data are mean ± standard error (M ± SE) of three replicates.
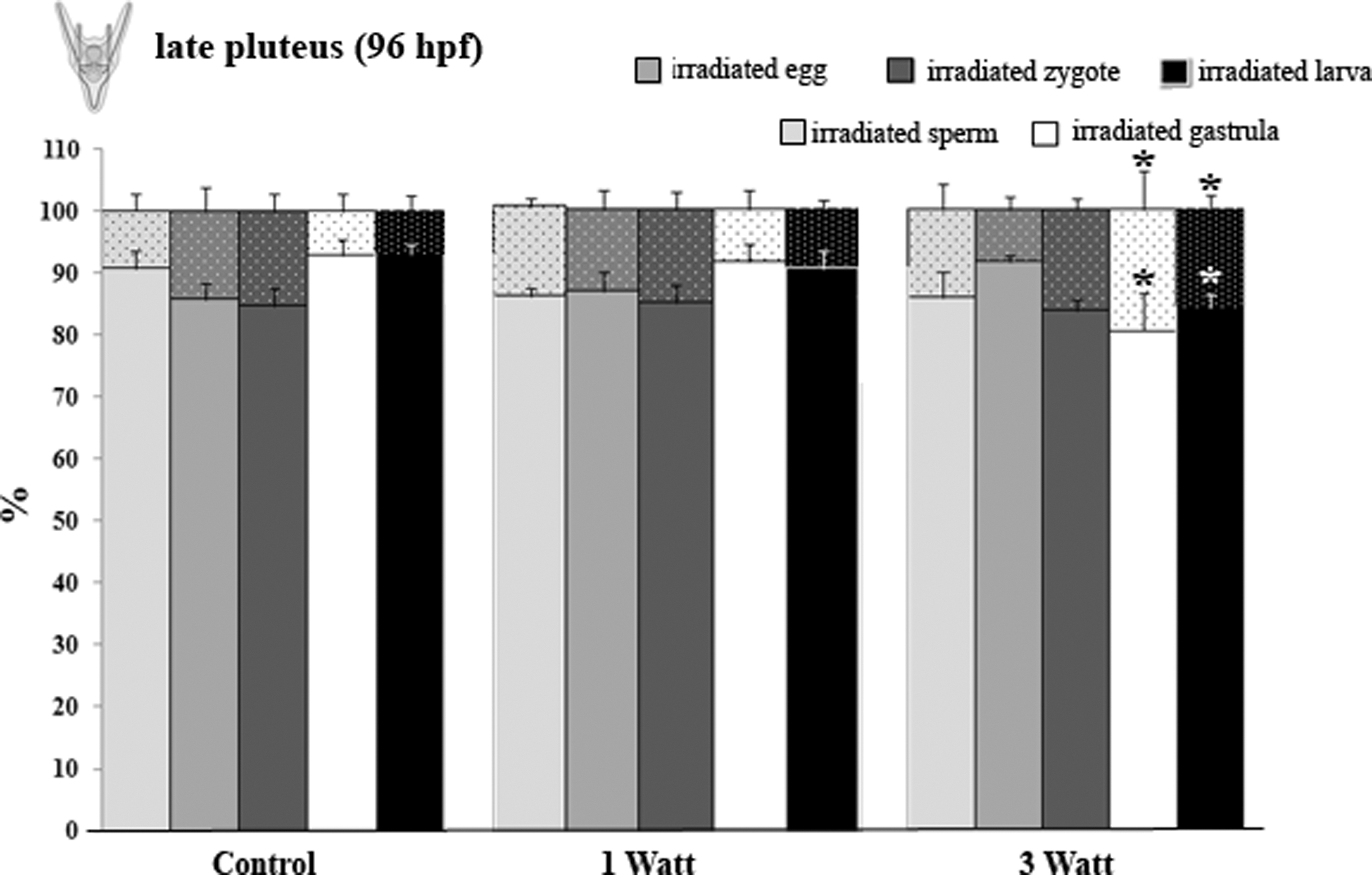
Shows the percentage of the developed (full bars) and the anomalous development (tiny dotted bars) from the irradiated sperm (light gray bars), egg (gray bars), zygote (dark gray bars), gastrula (white bars), and pluteus (black bars). Data are mean ± standard error (M ± SE) of three replicates. *p < 0.05.
In the early pluteus stage (Fig. 4), the percentage of anomalous development in the exposed samples at 1 and 3 W was very low in comparison to the control samples (p > 0.05). Since there were no differences observed between larvae monitored at 72 h and the early/late pluteus (48 and 96 hpf), the data were not recorded as significant.
In the late pluteus stage (Fig. 5), there were no differences in the developmental percentage observed between the control and the treated samples, with the exception of larvae from gastrulae and larvae, which were irradiated at 3 W (p < 0.05). Similarly, the percentage of the anomalous development was significantly different in these samples, when compared to the controls.
Discussion
Recent studies have shown a positive contribution of PBM on in vitro fertilization. 11 However, if the effect of PBM on the gametes is evident, almost nothing is known of the embryogenesis of organism deriving from irradiated gametes, irradiated zygotes, and irradiated embryos. The results of our study showed that PBM does not exert any negative effect on the fertilization rate and the early developmental stages of the sea urchin after 1 W of irradiation.
In this regard, the authors have demonstrated that gamete and zygote irradiation at 1 and 3 W did not exhibit any negative effect on the success of the fertilization and the offspring development. However, the opposite effect was observed in the late plutei (96 hpf) at 3 W irradiated embryos and larvae. This effect was detected by observing and quantifying the high percentage of stress morphological markers in comparison to the control samples.
In this study, it is worth mentioning that the 3 W power setting of irradiation does not exert any effects on gametes and zygotes and subsequently on the fertilization rate. However, this power setting showed an effect on the embryo and larval stages. This trend was not observed in gastrulae and plutei, which were irradiated at 1 W power setting. These observations by the authors suggested that higher power may have an effect on the embryo and larval stages, which can lead to alterations in signal transduction and cell-to-cell communication, causing these anomalies in the offspring. Falugi 45,46 studies showed that the ability of the ACh to cause an effect on the gastrulating sea urchin embryos was related to the receptors of the neurotransmitters that were expressed at the time of gastrulation when cells interacted not only with each other but also with the extracellular environment. 47 Conversely, before this time, the embryo has a limited ionic change and is surrounded by a fertilization envelope.
The specific antagonists of the neurotransmitter receptors (i.e., ACh) act as inhibitors of morphogenetic cell movements during specific phases of gastrulation. 48 In this regard and within the limitations of our results, we hypothesize that 3 W irradiation may prevent the ligand (ACh) from binding to its receptor since it has been demonstrated that low-intensity near-infrared laser radiation (810 nm) produces changes in the acetylcholinesterase activity (the enzyme responsible for ACh degradation) of human erythrocytes. 49
Since the effect of PBM on the movement of the sea urchin sperm flagellum is not part of our investigation, we have not measured it. However, our data show that our parameters photobiomodulate sea urchin sperm, since we observed an early development if compared to fertilized eggs from nonirradiated sperm. PBM has been demonstrated to have both primary and secondary stimulatory effects on different cell types, including sperm. 50 In this regard, 632 nm laser irradiation of human sperm as well as PBM with light-emitting diodes seems to improve the motility and speed. 14 –17 Likewise, we found a positive effect of 808 nm diode laser on sea urchin sperm, according to previous reports on the sea urchin Strongylocentrotus purpuratus. 13
PBM was only found in zygotes from irradiated sperm and not in zygotes/offspring from irradiated eggs, zygotes, or early developmental stages.
The stimulatory effect was observed in sea urchin sperm that does not exert any negative effect on the fertilization rate and does not induce abnormality in sea urchin early developmental stage after it was irradiated with 1 W. It can be assumed that our irradiation of sperm may have accelerated the physiological sequences of biochemical and biophysical developmental events, as this has already been hypothesized for electromagnetic waves on sea urchin early development. 51 Laser irradiation applied for a short time with different output powers (1 and 3 W) has accelerated the cleavages of sea urchin zygotes, which allowed normal development.
Previously published works have shown that the potential of low-power laser irradiation of sea urchin spermatozoa increased their motility. 8,11 Laser irradiation may have an effect on the Ca2+ binding sites of sea urchin sperm structure, stimulating its release during the fertilization process and first cleavage. When the membranes are fused, the sperm nucleus, mitochondria, centriole, and flagellum can enter the sea urchin egg and variations in Ca2+ flux rates can be observed as well as cleavage stimulation in sea urchin embryos. 46
Many authors have suggested that modifications of the Ca2+ flux rates are as a consequence of laser irradiation on other species. Shahar et al. 18 reported that human sperm hyperactivated motility irradiated with visible light (400–800 nm) has increased significantly, while the concentration of intercellular Ca2+ accompanied with a reduction in the hyperactivated motility rapidly increased when the voltage-dependent Ca2+ channel was blocked. Wenbin et al. 52 have found that laser irradiation increased Ca2+ absorption in Saanen bucks sperm as well as its motility and survival time. Likewise, laser irradiation may be involved in Ca2+ flux rates in sea urchin sperm as well as increasing its motility. 8
Paul et al. 53 showed that when the sea urchin Arbacia punctulata eggs were irradiated with low-level laser energy before fertilization, no microscopic changes occurred in the eggs after irradiation. However, many developmental abnormalities appeared after fertilization. Also, a pronounced delay in postfertilization DNA synthesis was observed. Karu 54 detected that higher fluences caused destruction of photoreceptors, which was accompanied by growth inhibition and cell lethality. Further, researchers have demonstrated that irradiation with fluences higher than 10 J/cm2 damages DNA synthesis. 55
The discrepancy between results in our study and the literature could be due to the use of a different hand piece in the experiments (Gaussian vs. flat-top) that highlighted the AB2799 features, thus allowing higher fluence irradiation without thermal effects; additionally, flat-top delivery provides uniform PBM dose values of parameters used, as shown in our previous articles. 2,3,7,22
However, the application of a higher dose of 192 J/cm2 on the stage of gastrula and larva resulted in pluteus anomalies, as the adverse result with the higher fluence PBM in in vitro fertilization can induce damage. In consequence, the higher fluence has to be carefully utilized and analyzed. Indeed, Amaroli et al. 31 showed that every stage of the early developmental process of the sea urchin has to occur at a precise moment. Otherwise, redundancy and disorder in the formation of structures and systems appear, because of the effect on a peculiar physiological process 31 that could be the target of PBM.
Conclusions
This study has demonstrated that both the fluence (64 J/cm2–1 W) and higher fluence (192 J/cm2–3 W) do not induce damage on the irradiated sea urchin gametes, whose zygotes generate normal embryos and larvae. Also, the overstimulation of some sperm leading to an accelerated cleavage of sea urchin zygotes is not deleterious to a correct embryogenesis of P. lividus. In consequence, our data support the assumption to use the higher fluence values in preliminary studies on in vitro fertilization.
Footnotes
Author Disclosure Statement
All authors disclose any commercial associations that might create a conflict of interest in connection with the submitted article.