
Abstract
Introduction
M
Harvest of MSCs from the patient's own body fat depots can be carried out using less invasive techniques; so a protocol for satisfactory neuronal differentiation of these precursor cells would make this form of autologous stem cell therapy very promising. Specifically, adipose stem cells (ASCs) obtained from subcutaneous tissues or lipoaspirates are resistant to apoptosis and much more likely to undergo differentiation than any other types of adult stem cells.
3
A comparative analysis has found that ASCs accelerate nerve repair and are more efficient in differentiating into neural and vascular endothelial cell types.
4
In fact, the greatest advantages of using ASCs are: (1) lack of social and ethical concerns that apply to embryonic stem cells; (2) ease of isolation and abundance in patients; (3) good capacity for proliferation and differentiation; (4) minimal risk of histo-incompatibility and disease transmission; and (5) ability to pass through the blood–brain barrier to lesion sites in the brain. Given their ability to replenish cell components in vivo, ASCs are now used in clinical trials for various conditions, including stroke (NCT02813512,
In the developing brain, neural growth occurs by the generation of neural “rosettes,” which morphologically represent the neural tube. Neural precursor cells (NPCs) located in these neural “rosettes” in experimental animals can be isolated and expanded to generate neurons and glial cells in cell culture. NPC therefore is a mixed population of neuronal stem cells (NSCs) and neural progenitor cells, which can give rise to neurons and supporting cells. 5 Transdifferentiation is a process by which the multi-potent cells from the original germline are changed to another germ layer in vitro. This is experimentally performed by subjecting ASCs to small molecules or physical agents designed to alter cell physiology that will mimic the developmental cues of the brain during organogenesis. Although several of these methodologies are reviewed and modified time after time, there seems to be no ideal protocol that could meet the demand of these cells in clinics. A better alternative would be using these small molecules along with physical agents such as light or electricity for differentiating ASCs in culture. Photobiomodulation is the efficient and safe application of low-intensity laser irradiation that may contribute to increased yield and viability of ASCs differentiating into specific neuronal lineage cells, while enhancing its differentiation capacity.
Induction of Differentiation into Neuronal Lineages
Growth factors and chemical compounds
Various inducers, influencing cell morphology and functionality, are used for the differentiation of ASCs into other lineages. Most of these inducers are biological or chemical factors that have a proliferative effect as well. Biological inducers include a variety of growth factors that are efficient and safe to use in cell culture conditions. They are potent regulators of cell proliferation and differentiation, and are important in normal development, tissue maintenance, and wound repair.
In fact, growth factors such as brain-derived nerve factor (BDNF), basic fibroblast growth factor (bFGF), nerve growth factor (NGF), and neurotrophic factor-4/5 (NT-4/5) serve to promote the survival and differentiation of NPCs to neurons in CNS (Table 1). The neurotropin, NGF, activates tropomyosin receptor kinase A (trkA), which plays a vital role in neural induction and differentiation. 6 It effectively modulates signaling pathways, especially PI3K/Akt, which increases ASC survival and Rac activity (a member of the Rho family of GTPases) leading to changes in its shape and migration potential. Moreover, polarization of Rho/myosin II components in response to growth factors alter cytoskeleton of differentiating cells to resemble fibroblast processes. 7
BDNF, brain-derived neurotrophic factor; EGF, epidermal growth factor; FGF, fibroblast growth factor; GDNF, glial cell line-derived neurotrophic factor; HGF, hepatocyte growth factor; IGF, insulin-like growth factor; JAK-STAT, janus kinase-signal transducer and activator of transcription; MAPK/ERK, mitogen-activated protein kinases/extracellular signal-regulated kinases; NGF, nerve growth factor; PDGF, platelet-derived growth factor; PI3K/Akt, phosphoinositide 3-Kinase/v-akt murine thymoma viral oncogene homolog 1; RA, retinoic acid.
Similarly, hepatocyte growth factor (HGF) is another neurotrophic factor, which is associated with the growth and survival of sensory and parasympathetic neurons. It has multiple functions such as axonal chemoattractant to spinal motor neuron, growth promotion to neocortical explants, as well as survival factor of motor neurons in the PNS. 8,9 Basic FGF and NGF along with retinoic acid (RA) induce neuroglial differentiation of bone marrow MSCs. 10 Human umbilical cord-derived stem cells induced by nor-epinephrine, a neurotransmitter, can generate oligodendrocytes in spheroid in vitro cultures. 11 Together, ciliary neurotrophic factor (CNTF), interleukin-6, and epidermal growth factor (EGF) are capable of differentiating NPCs to astrocyte lineages. 12
Chemical compounds are also used as neuroglial inducers, (Table 2). Agents like β-mercaptoethanol (a thiol compound) promote neuronal survival in culture by increasing cysteine uptake and intracellular glutathione. 13 Similarly, butylated hydroxyanisole, potassium chloride, valproic acid, forskolin, hydrocortisone, and insulin are agents that can act individually, or in combination, for neural or glial differentiation. 14 A combination of chemical inducers viz. β-mercaptoethanol, butylated hydroxyanisole, and dimethylsulfoxide promotes neuronal differentiation of MSCs. 15 Various chemical inducers such as ethanol, potassium chloride, valproic acid, as well as selected hormones, including hydrocortisone, and insulin can also achieve neuronal differentiation of ASCs. 16 Forskolin activates adenylate cyclase for increasing intracellular cAMP (cyclic adenosine monophosphate) levels, whereas 3-isobutylmethylxanthine (IBMX) acts as an inhibitor of phosphodiesterase and phosphorylation by protein kinase A. 17 Chemical inducers have a limited use due to their toxicity when used alone or in combination with their counterparts; however, individual effects of these chemicals on stem cell differentiation are fairly understood, and their combined effects still need to be investigated.
cAMP, cyclic adenosine monophosphate; GABA, γ-amino butyric acid; IBMX, 3-isobutyl-1-methylxanthine; Shh, sonic hedgehog.
ASC differentiation into neuronal cells has also been demonstrated successfully using a combination of growth factors and chemical compounds. Neuronal differentiation induced by growth factors viz. bFGF, forskolin, and NGF leads to the expression of surface markers of mature neurons within 1 week. 18 Ashjian et al. demonstrated the combined effect of IBMX, indomethacin, and insulin in the differentiation of processed human lipoaspirate cells into neuron-like cells. 19 However, these differentiated neuron-like cells failed to express mature neuronal or glial markers and demonstrated no electric conductivity (Table 3). Differentiation of stem cells to specific neurons releasing neuro-transmitters is also feasible by the combined use of biological and chemical inducers. Cultured bone marrow MSCs were seeded in serum-free media containing 1 mM β-mercaptoethanol for 1 day followed by incubation with 100 ng/mL NGF for 6 days as a cholinergic neuron inducer. 20 MSCs can also be platelet-derived growth factor (PDGF) cultured in media containing 0.5 mM IBMX for 2 days and transferred into an induction medium containing glia-derived nerve factor (GDNF) (10 ng/mL) and IL-1β (100 pg/mL) to obtain dopaminergic neurons. 21
FBS, fetal bovine serum; GFAP, glial fibrillary acidic protein; GAP-43, growth-associated protein-43; MAP2, microtubule-associated protein2; NCAM, neural cell adhesion molecule; NeuN, neuronal nucleus; NSE, neuron-specific enolase; Trk-A, tropomyosin receptor kinase-A.
Additional studies showed the combined effect of RA, bFGF, and NGF in directing bone marrow MSCs toward neuroglial differentiation, 10 and a combination of β-mercaptoethanol and RA induced the differentiation of MSCs into Schwann cells. 22 This method has been extended to ASCs as well for the in vitro generation of Schwann cells. 23 Similarly, a cocktail of hormonal and chemical factors, such as PDGF, bFGF, heregulin, and forskolin, increases myelination and differentiation toward Schwann-like cells. 24,25 Also, Schwann cells secrete survival factors such as insulin-like growth factor (IGF), NT3, PDGF-BB, leukemia inhibitory factor (LIF), and lysophosphatidic acid (LPA) in an autocrine manner. 26,27 Together, these supplemented chemicals as well as secreted survival factors are capable of bringing differentiation of ASCs to a specific glial lineage in vitro. Generation of oligodendrocytes, NPCs, is plated onto poly-D-lysine-coated dishes and grown in customized media containing DMEM/F12 high glucose, N2 supplement, GlutaMAX, bFGF, EGF, and PDGF-AA (34).
An improved strategy for differentiating ASCs, while ensuring increased yield and viability, would be to adopt methods shown in Fig. 1. Freshly harvested ASCs incubated at 37°C under 5% CO2 can be induced to differentiate into neurons by growth factors. 28 ASCs are used to generate specific neurotransmitter release neurons by a combination of biological and chemical inducers. Similar to using MSCs in generating cholinergic neurons, ASCs may also be treated with β-mercaptoethanol as a pre-inducer and NGF as an inducer in a culture medium. 20 In a similar way, ASCs can be induced to dopaminergic neurons after incubation in IBMX for 2 days followed by a medium containing several other inducing agents for 15 days. 21 Darbinyan et al. have provided a detailed protocol for differentiating NPCs to oligodendrocytes in culture. In this study, neural stem cell medium and the glial medium enriched with several growth factors are used in equal amounts to make up the oligodendrocyte differentiating medium. 29 Although labor intensive, this method is quite robust in producing oligodendrocytes and can be employed to induce differentiation of ASCs as well. For Schwann cell differentiation, subconfluent ASCs are incubated with β-mercaptoethanol for a day and then shifted to a stem cell growth medium with RA. 23
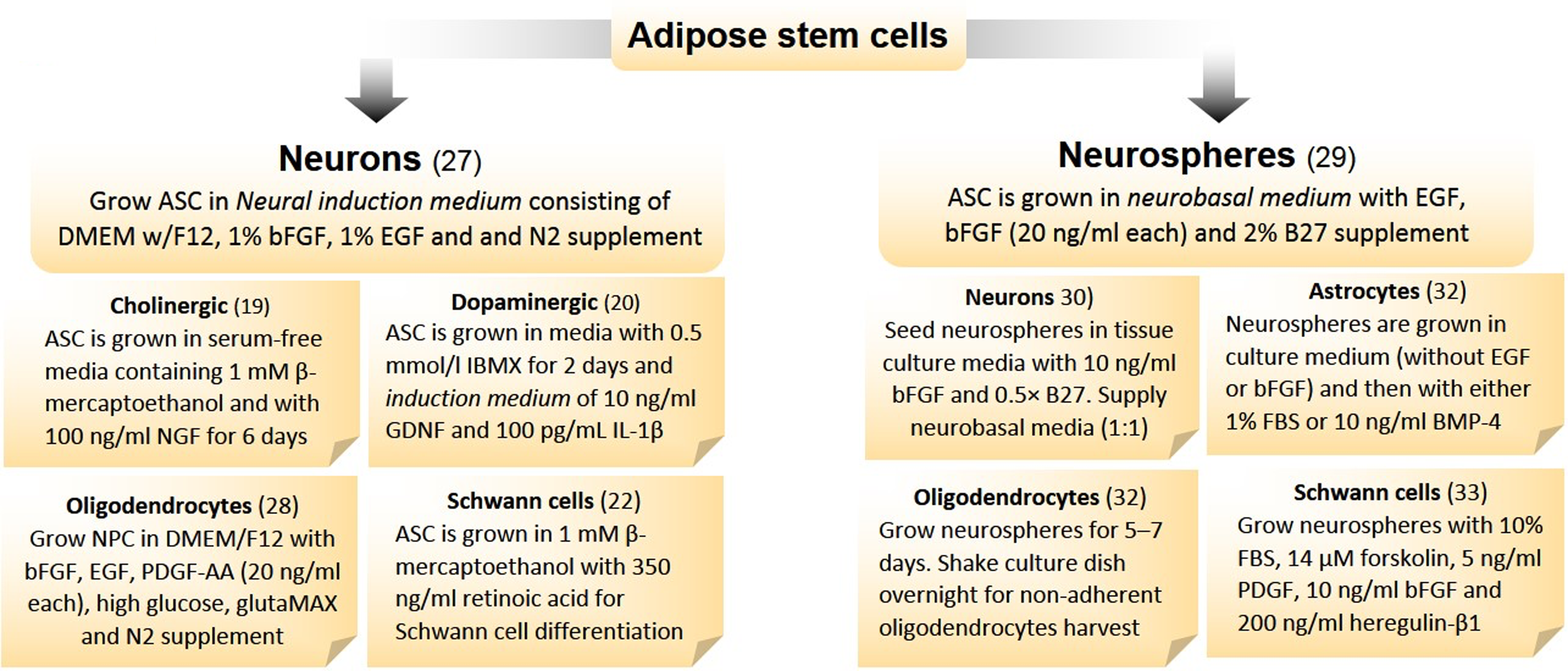
Transdifferentiation protocols for neuroglial cells. Isolated adipose stem cells can be differentiated into neurons in adherent and neurospheres in suspension cultures. For detailed protocols, researchers requested to refer respective publications. ASC, adipose stem cells; BMSC, bone-marrow stem cells; BMP4, bone morphogenetic protein-4; DMEM, Dulbecco modified Eagle's medium; EGF, epidermal growth factor; bFGF, fibroblast growth factor (basic); GDNF, glia-derived nerve factor; IBMX, 3-isobutylmethylxanthine; IL-1β, interleukin-1β; MSC, mesenchymal stem cells; NGF, nerve growth factor; NPC, neuro-progenitor cells; PDGF, platelet-derived growth factor.
Neuronal differentiation of stem cells can be achieved either by differentiating them to neurons or neurospheres in culture, and the latter subsequently be transformed into neurons (Table 4). In this study, a combination of biological and chemical inducers acts jointly on stem cells to differentiate it into various neurotransmitter neurons or glial cells in vitro. Neurospheres can be prepared by growing ASCs in a neurobasal medium without serum, but with EGF, bFGF, and B27 supplement. 30 These free-floating spheres are mechanically or enzymatically dissociated into a single cell suspension when plated on substrates such as poly-L-ornithine, laminin, or fibronectin under low serum conditions. Neurospheres transition to neurons by growing them in the culture medium with bFGF and B27 supplement (EGF withdrawal). 31 Neurospheres generated from MSCs can be differentiated into dopaminergic neurons using forskolin, IBMX, and dibutyrl-cAMP through activation of cAMP pathway. 32 They can also be differentiated to astrocytes and oligodendrocytes by biological and/or mechanical means. 33 Moreover, Schwannn cells are differentiated from these cultured neurospheres by specific lineage inducers. 34 This schema of differentiation may hold well in terms of time lapse and outcome and can easily be adopted in laboratories with minimal arrangements.
HRG, heregulin; microtubule-associated protein2; S100β, S100 family protein; TTX, tetrodotoxin.
Co-culture systems
Co-culture systems are the dynamic association of two or more cell types that allows the growth and differentiation of one cell type under the influence of the other. In this study, the growth stimulatory effect, as well as the fate of the cell with respect to differentiation, is brought about by changes in signaling pathways and secretion of growth factors. Co-culture has the benefit of growing different cell lines in common media, defining cell growth conditions for differentiation and measurement of growth characteristics. However, growing cells in a co-culture is limited by the difficulty in identifying the signaling process as well as the quantification of extrinsic factors during the differentiation process. Often, the results of these co-culture methods or chemicals combined with a set of growth factors are inconclusive. Occasionally, co-cultures may also be intermingled with fibroblasts, creating difficulties in identification and segregation. Thus, use of a transwell plate with two or more chambers is a better choice that allows the propagation of a differentiating cell line in constant communication with its inducing cell line. This has the benefit of not mixing up two cultures that may cause one culture to take over the other or deliver confounding results.
ASCs differentiate into smooth muscle cells in a co-culture using SKUT-1 cells with an increase in the expression of SMC markers and decrease in the expression of stem cell markers. 35 Both, CD29 and Thy-1, stem cell marker expressions decreased significantly in co-cultures that had growth factors, TGF-β1 and RA. 36 ASCs could be co-cultured with glial cells for initiation of differentiation and long-term maintenance into a glial lineage. Co-culture of ASCs with Schwann cells has shown to induce differentiation of ASCs into a Schwann lineage. Direct contact between Schwann cells and ASCs may boost phenotypic and functional changes. A more efficient approach may be to use transwell dishes with a polycarbonate membrane separating Schwann cells from ASCs, but allowing growth factor exchange. Adult Schwann cells in this co-culture secrete BDNF and NGF for induction of neuronal progenitors. 37 Additional studies have also indicated the use of rat sciatic nerve leachate, which has an admixture of glial cell differentiating factors as an alternative for ASC differentiation into Schwann-like cells. 27
Transcriptional inducers
Somatic cell reprogramming seems the most promising approach in creating neuronal lineage cells. Transcription factors may influence differentiation of ASCs. Assembly and function of transcription factors are dictated by the cell milieu, which is influenced by extrinsic factors in the cell growth medium. Reprogramming of adult somatic cells to neurons using appropriate transcription factors is a convenient option for replacing lost or damaged CNS cells. Recent studies have shown the direct conversion of mouse and human skin or liver cells into neurons, termed “induced neurons.” Human fibroblasts from a skin biopsy were converted into neurons by a combination of transcription factors ASCL1, BRN2, and either MYT1 L or ZIC1. 38 Induced neurons were also generated from human dermal fibroblast cells using the transcription factors MIR124, BRN2, and MYT1 L. Induction of neurite outgrowth by a combination of FGF receptor 1 treatment and peripheral nerve grafts successfully restored hind limb function in animal models. 39,40
Differentiation of ASCs can be achieved within the mesoderm or can be switched to ectoderm and endoderm by overexpression of a lineage-specific transcription factor. 41 ASCs are reprogrammed using polycistronic plasmid with inserts for OCT3/4, SOX2, KLF4, and c-MYC and then differentiated into neuronal lineage cells using well-defined induction media. 42 This method eliminates the need for embryoid body formation required during normal somatic cell reprogramming. It also excludes the stringent growth conditions, while reducing the risk of differentiation into undesired cell types. Transfection of ASCs using a retrovirus with a sole transcription factor gene, SOX2, promotes NSC formation. The transduced ASCs demonstrate NSC phenotype and are capable of differentiating into neuron-like cells. 43 Similarly, astroglia can be reprogrammed to functional neurons by expression of Pax6 or the proneural transcription factor neurogenin-2. 44 Transplantation of fetal dopaminergic neurons to an adult suffering from Parkinson's disease has shown promising results. 45 Possibly, induced neurons and glial cells can be used for such replacement therapies in the future.
Physical factors used for differentiation
Electrical stimulation
Electricity provides physical stimulation of ASCs to differentiate into other lineages, especially neurons. Direct current causes ASCs to orient themselves perpendicularly to the field vector and exhibit a dose-dependent migration toward the cathode. This reorientation is accompanied by reorganization of their cytoskeleton and organization of a well-formed lamellipodium simulating dendrite arborization. Further, there is a transient increase in cytosolic calcium causing activation of growth factors and membrane signaling components. 46 Even when cultured in the absence of biochemical growth factors, MSCs respond to stimulation by a pulsed electric field by switching to a neuron-specific lineage. Electrical stimulation results in differentiation of cells by creating changes in gap junctions, altering the density and distribution of ion channel receptors, production of free radicals, and modification of various cellular signaling pathways. Together, these changes affect cellular morphology and variations in membrane potential, enabling hyperpolarization and depolarization.
Direct current electric fields can guide neurite growth and encourage migration of neurons and glial cells. 47 Studies investigating the effects of electrical stimulation on MSC differentiation into neurons have documented that an extremely low-frequency pulsed electromagnetic field resulted in the expression of osteocyte and osteogenic genetic markers. 48,49 Similarly, a brief exposure to low-frequency electrical stimulation increased axonal myelination as well as oligodendrocyte survival in co-cultures. 50,51 Signaling by cAMP activated the ATP-LIF pathway in astrocytes to promote myelination along with various other cell components. 52 Neurotrophins enhance myelination of Schwann cells by electrical stimulation, indicating that biological and physical inducers may act together to improve the functionality of differentiated cells. 53
Transdifferentiation of MSCs can also be achieved by the interaction of cells with bioactive matrices possessing electrical conductance and microfluidic systems for three-dimensional monitoring of neural differentiation. 54,55 Such novel technologies are useful in creating microenvironments suitable for neuronal differentiation and axon elongation of ASCs by fine tuning its signaling and redox status. 56,57 These matrices are also capable of altering the electroresponsive properties of stem cells, thereby modulating cellular adhesion and differentiation to neural or muscle cells. 58 Interestingly, a very low voltage (1–11 mV) electrical stimulation of ASCs seeded on a gold microelectrode array showed alteration in its growth function and energy metabolism, indicative of a neural phenotype. 59 A combination of electrical stimulus with chemicals or growth factors and the use of nanomaterials may result in the differentiation of MSCs into neural lineage cells.
Electrical stimulation of MSCs is also facilitated by surfaces of different conductivities (such as polyaniline) resulting in higher levels of expression of neuronal markers. 60 In this study, the differentiation of MSCs into neuron-like cells was observed when grown on 0.1 M HCl-doped polyaniline substrates, with neuronal differentiation occurring in a conductivity dependent manner. Even without any doping application, a steady potential of 1.5 Volts for an hour resulted in differentiation of NSCs into mature neuron-like cells on the polyaniline substrate. 61 In another study, MSCs demonstrated significant filopodial elongation as well as a decrease in nestin expression simultaneously, with an increased expression of βIII tubulin. 62 Electrical stimuli and substrate conductivity can result in a controlled release of free radicals in living systems. This results in changes to cellular redox status capable of modifying signaling pathways necessary for transforming stem cells to other cell types.
Photobiomodulation
Photobiomodulation refers to the effects of low-intensity laser irradiation or light-emitting diodes on biological tissue, which alters cellular function. The process of photobiomodulation occurs by changes within a living cell after absorption of photons by chromophores or photoacceptors. Although the actual mechanism is still elusive, this process alters cell signaling cascades and downstream effector molecules leading to changes in cell functionalities. 63,64 Application of specific wavelengths, power densities and fluences, of monochromatic light to ASCs may result in differentiation consequential to its biostimulatory effects. Light can alter mitochondrial respiratory rate generating reactive oxygen species (ROS), which changes cellular signaling cascades and the rate of cellular metabolism, leading to cellular proliferation, migration, and differentiation. Any changes in the cell milieu by light cause mitochondria to generate ROS, which changes the whole process of cell signaling. Further, a surge of calcium in the cytoplasm and an increase in mitochondrial energy production are observed (Fig. 2). Photons have the ability to oligomerize and activate receptors for growth factors on cell surfaces leading to autophosphorylation of cytoplasmic kinase domains and onset of cell signaling events. 65
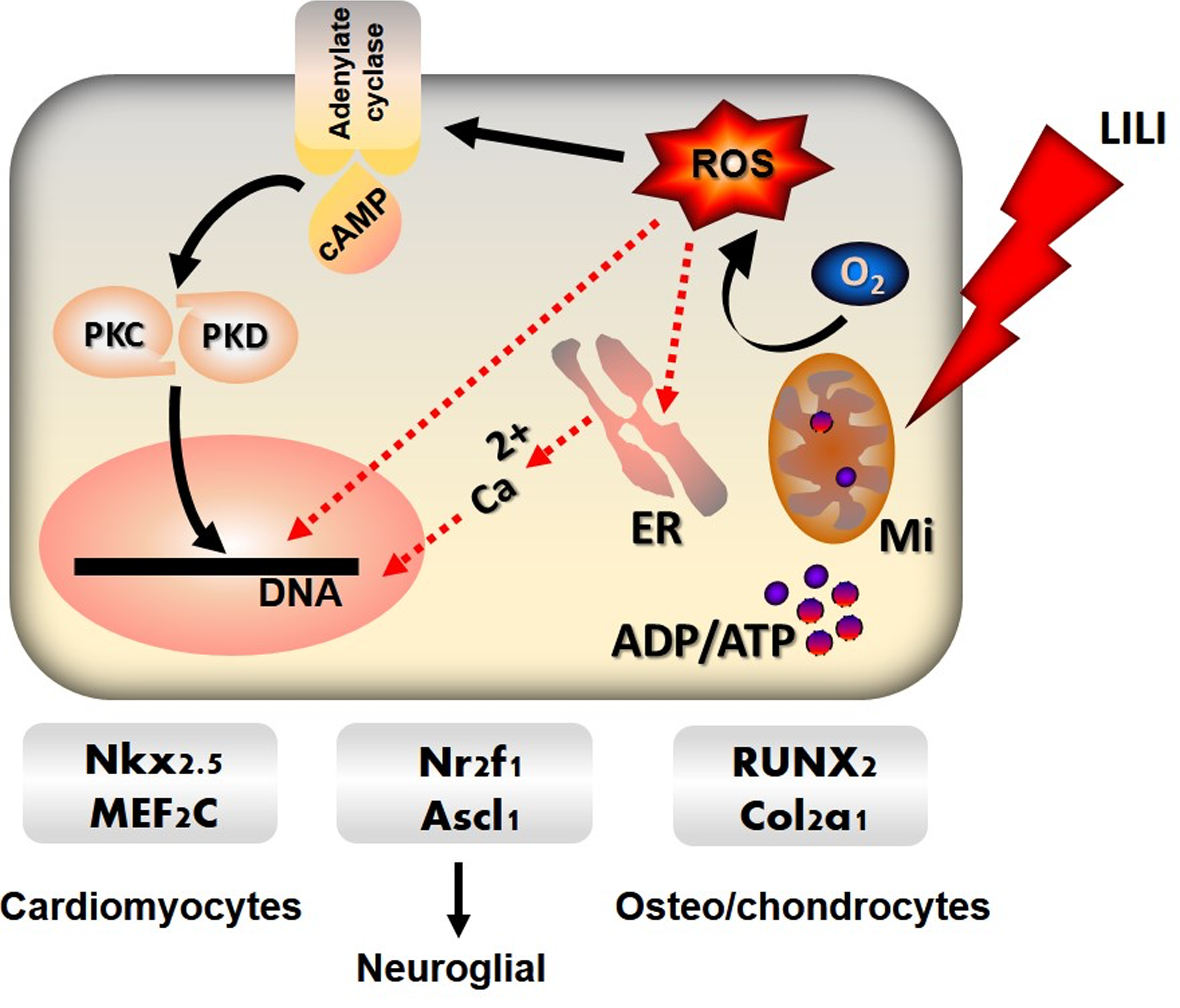
Effect of Photobiomodulation and specific inducers on ASC differentiation. LILI absorbed by chromophores in the mitochondria (Mi) generate ROS from oxygen (O2), which cause the cell membrane enzyme adenylate cyclase to release cAMP and activate protein kinase receptors (PKC, PKD). Photobiomodulation promotes energy metabolism and release of calcium (Ca2+) from (ER) that enters the nucleus to change the dynamics of stem cell growth. This process, in conjunction with biological or chemical inducers, increases the expression of specific transcription factors for lineage-specific differentiation of stem cells. Solid lines are cell signaling pathways and dotted red lines are ROS-mediated interactions. ER, endoplasmic reticulum; LILI, low-intensity laser irradiation; ROS, reactive oxygen species.
Treatment using light leads to proliferation and/or osteogenic differentiation of mouse bone marrow MSCs under the effect of IGF1 and BMP2. 66 The crosstalk between these interventions, for proliferation and differentiation, on the same target cells, remains obscure, although the p65 subunit of NF-kB is speculated to bind to the Smad1/4 complex and interfere with BMP2 signaling for osteogenesis. 67 A low-energy Gallium-aluminum-arsenide (GaAlAs) laser delivering 805 nm and 5.9 J/cm2 induced expression of BMP2 and phosphorylation of Smad/5/8, but not expression of BMP4, IGF-1, or inhibitory Smads. 68 GaAlAs lasers of 660 nm at 2 and 4 J/cm2 induced proliferation and differentiation in human periodontal ligament cells to osteoblasts by cAMP regulation. 69 Similarly, 810 nm GaAlAs lasers at 3 J/cm2 generated ROS in a dose-dependent manner with activation of latent TGF-β1 signaling capable of differentiating host stem cells to odontoblasts in vitro and in vivo. 70 Low-intensity laser irradiation (LILI) induces ROS production and promotes osteoblast differentiation and the activation of redox factor-1 (Ref-1)-dependent AP-1, indicating that the cellular redox status may also be a key player for inducing differentiation. 71
LILI is a technique where cell lines are exposed to various wavelengths and energy levels of laser to bring benevolent effects on biological systems. It has many unique properties viz. coherence, monochromacity, compressibility, and collimation that are essential for its activity in living systems. 62 LILI at wavelengths ranging from 600 to 900 nm and energy densities less than 10 J/cm2 can alter the intracellular levels of ROS in a controlled manner to enhance cell growth, proliferation, and differentiation of biological systems. LILI can alter the rate of cellular metabolism, which may empower mitochondria leading to cellular proliferation, migration, and differentiation. Further, lasers stimulate DNA synthesis as well as increase protein expression and viability and migration of stem cells in vitro. 72,73 Most likely, LILI in combination with various concentrations of growth factors may play a role in the regulation of cellular process and differentiation of human ASCs.
Studies have revealed that ASCs exposed to 5 J/cm2 LILI using a 636 nm diode laser increased its' proliferation and viability at 24 and 48 h with no changes on its cell surface morphology. 74 Low-intensity laser irradiation is also found to promote proliferation and differentiation of human osteoblast cells in vitro at 632 nm with a power output of 10 mW. 75 The beneficial effect of photobiomodulation is manifested in the treatment of brain injuries and neurological disorders. 76 –78 Photobiomodulation can induce conformational changes to various microtubular proteins in response to oxidative stress, resulting in changes to the cytoskeleton. This is manifested as neural membrane stretch, often resulting in the alteration of conductivity of membrane ionic channels. These sequences of events are often a prelude for the morphological changes in ASCs responsible for neuronal differentiation and impulse transmission. 79 However, it was unsuccessful in differentiating ASCs to smooth muscle cells at a fluence and wavelength of 5 J/cm2 and 636 nm, even in the presence of retinoic acid. 80 Perhaps, these conflicting data motivate us in testing a wide range of laser parameters on ASCs for differentiation to body cell types in vitro.
Challenges and Future Perspectives
Problems associated with the isolation, quantity, viability, and proliferative ability of ASCs ex-vivo for stem cell therapy-based treatment modalities have created the need for appropriate research methodologies. During neuronal differentiation, ASCs constitutively express native immature neural proteins, nestin and β-tubulin III, whereas more mature neuronal and glial proteins viz. TH, MAP2, and GFAP are expressed at higher passage numbers. 81 Often, these undifferentiated ASCs express markers characteristic of neural cells such as NSE, MAP1b, NeuN, nestin, and vimentin. 19,82 All these inconsistencies indicate the heterogeneity in the expression of neural markers that might have confounding effects on the identification protocols for differentiated neural cells. Morphology of ASCs in culture shows the onset of cell senescence pathways beyond passages 10, which may have implications for its use in differentiation protocols. 83 Stem cells have a fibroblastic morphology, and, when exposed to stress, undergo changes in cytoskeleton with alterations in their levels of expression of genes, resulting in the variable levels of markers on cell surface. 84,85 This is reflected as changes in the levels of certain antigens (markers) per unit cell area, mimicking differentiation to neuronal lineage, and may not be markers of a true neuronal cell. All these inconsistencies need to be addressed carefully while working on human samples.
Neuronal induction and the expression of specific markers during transdifferentiation processes are often incomplete and reversible. Neuronal lineage cells obtained by the cultivation of ASCs in a neuro-differentiative medium are often a mix of cells expressing one or more neuro-specific markers. 86 Perhaps, differentiated neurons and glia need to be sorted before identification, speciation, and use of these newly differentiated neuronal lineage cells in clinical trials. After differentiation, variability in the expression levels of neural transcription factors in MSCs such as Engrailed1 and Nurr1, related mRNAs such as DRD2, NSE, NFL, MBP and STX1A, has been observed among tissue donors. 82 Similarly, neuronal induction by chemical protocols results in disruption of the cytoskeleton, causing an increase in overall protein concentrations. 85,87 Further, a coaxed differentiation of cells from one lineage to another lineage from an unnatural germ layer could result in abnormal phenotypes, which may end up in carcinogenesis upon engrafting into human patients. 88 Hence, the differentiation protocols should be well optimized not only in terms of yield but also the functional (electrophysiological) status of neurons. Even well-defined, directed differentiation protocols toward neuroglial lineages may be insufficient for successful use due to the genetic background of the patient.
Adult stem cells isolated from the adipose tissue of abdomen are multi-potent and capable of differentiating to various lineages. Neuronal differentiation of these cells seems to be well regulated by signaling pathways, which may even crosstalk by unknown circuitry. Likewise, neurogenic to gliogenic switch is dependent solely upon the intrinsic timing, signaling pathways, and environmental cues. Use of LILI combined with specific inducers will be able to address various challenges in this process. A very good reason is the differentiation of ASCs to neural cells by combining Nurr-1 gene transduction with electrical stimulation. There is not only a higher rate of differentiation but also an increased functionality manifested by the extension of neurites and neurotransmitter release. 89 Production of oxide- and nitrite-free radicals by electricity and light is tightly regulated and their interactions with membrane receptors and signaling pathways control the assembly of transcriptional factors for stem cell proliferation and/or neuronal lineage differentiation.
Thus, the use of various intrinsic and extrinsic factors is an absolute necessity for better outcome. Moreover, photobiomodulation is a therapeutic modality that is also proven effective for proliferating and differentiating stem cells to odontoblasts, osteo/chondrocytes, and cardio/myoblasts. Intense research is required to explore if LILI alone or in combination with inducers may provide an improved option for neural lineage differentiation for use in stem cell therapy.
Footnotes
Acknowledgments
We express our gratitude to Dr. Natasha Hodgkinson for suggestions and formatting this article according to the journal guidelines. This review article is supported by the South African Research Chairs Initiative of the Department of Science and Technology and National Research Foundation of South Africa (Grant No 98337). The authors sincerely thank the University of Johannesburg, Gauteng, South Africa, for financial support.
Author Disclosure Statement
No competing financial interests exist.