
Abstract
Background:
The lipid metabolism is essential for maintaining the body's energy responses. Laser photobiomodulation triggers many important cellular effects, but these effects on lipid metabolism are not well described. In this study, we analyzed the laser photobiomodulation in the hormone-sensitive lipase (HSL) activity, a key enzyme in the triglycerides (TAG) hydrolysis in adipose tissue 3T3-L1.
Methods:
Cells were submitted to the differentiation protocol in adipose cells, irradiated with 1, 2, and 3J with laser (904 nm—60 mw—laser diode) and incubated for 4 h after irradiation.
Results:
The response of laser photobiomodulation was able to trigger an inhibition of HSL activity (control = 0.057 ± 0.0008; 1J = 0.050 ± 0.0003; 2J = 0.0477 ± 0.002; 3J = 0.051 ± 0.002; p = 0.0003 against the control), but no modulation was observed in TAG levels into the medium (control = 26.5856 ± 0.52; 1J = 26.5856 ± 0.52; 2J = 27.2372 ± 1.41; 3J = 25.9991 ± 0.1303; p = 0.18).
Conclusions:
This is the first study of HSL activity modulation with laser radiation, suggesting that photobiomodulation can influence adipose tissue metabolism and open a new field of study.
Introduction
Mammals are able to store excess of consumed calories to supply their metabolic activities when needed. 1 Because of their hydrophobic characteristics, lipids can be anhydrous stored in large amounts on adipose tissue. 2 The adipose tissue is the main energetic reservoir, being controlled by endocrine signals. 3 In mammals, there are two varieties of adipose tissue: white adipose tissue, which stores excess energy in the triglycerides (TAG) form, and brown adipose tissue, which is specialized in dissipating energy through the thermogenesis. 4
The white adipose tissue is large, with mean diameters of 100–1250 μm, 5 and shows a spherical form. 6 In addition, they can markedly change their size according to the amount of TAG. 5 In energy restriction conditions, a lipolytic process, defined as TAG hydrolysis, generates fatty acids and glycerol, under the influence of catecholamine, which are released into the bloodstream for other organs use as an aerobic energy source. 7 The increase of intracellular cyclic adenosine monophosphate levels triggers the activation of protein kinase A (PKA), which phosphorylates the enzyme hormone-sensitive lipase (HSL), as well as pyrilipins. Phosphorylation of pyrilipins, through PKA, allows the phosphorylate HSL activity on the fat droplet. 8 Apfelberg 9 was the first to describe lipolytic responses in adipose tissue by low-level laser stimulation. In 1994, they performed a study on the application of laser in liposuction procedures to reduce inflammatory responses, however, without promoting fat reduction. 10 The effects of photobiomodulation on fat reduction are still controversial. 11,12 There are few reports regarding the functions of the laser in the metabolic processes of adipose tissue, especially in the action of catabolic pathways. 13 Therefore, we propose to verify the effects of laser photobiomodulation on catabolic responses in 3T3-L1 adipocyte differentiated cells.
Methods
Cell line and adipocyte differentiation protocol
The 3T3-L1 cell line was maintained in Dulbecco's modified Eagle's medium (DMEM) medium (2% sodium bicarbonate/10% fetal bovine serum (FBS), conditioned in 75 cm2 cell culture flask at 37°C/5% CO2). Upon 80% confluency, the cells are trypsinized with 1 mL 0.25% trypsin/ethylenediaminetetraacetic acid and transferred to new plates. A total of 1 × 10 3 3T3-L1 cells were transferred to 12-well plates and incorporated a differentiation medium (BMII medium, 0.5 mM IBMX, 1 μg/mL recombinant insulin, 0.25 μM dexamethasone, and 2 μM of rosiglitazone) for 48 h and maintenance with 1 μg/mL of insulin for 72 h for total differentiation.
Irradiation protocol
After differentiation, cells were washed with sterile phosphate buffered saline (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, and 1.8 mM KH2PO4) for irradiation. The cells were irradiated (904 nm—60 mw laser—1, 2, and 3J of total energy; 17, 33, and 50 s; 0.07 cm2 spot area; 14.2, 28.5, and 42.8 J/cm2 energy density; and 0.8 W/cm2 power density). The cells were then incubated (DMEM 10% FBS for 4 h at 37°C, CO2 5%).
TAG quantification test
In 96-well plates, 10 μL of the culture medium of irradiated cells and 1 μL of the reagent solution were incubated at 37°C for 10 min under constant stirring and was read at a 500 nm wavelength reader. All reactions were normalized with the blank and standard reagent.
Protein extraction protocol
The cells were diluted in 1 mL assay buffer (50 mM, a2PO4-pH 7.2) and sonicated for 2 min. The lysate was centrifuged (10,000g/10 min at 4°C) and the supernatant was discarded. The pellet was conditioned at −80°C until its use. For the protein quantification or enzyme assay, the pellet was resuspended in the lipase activity assay kit buffer solution (Lipase Activity Assay; Sigma Aldrich, USA).
Protein quantification
The proteins were quantified by the 660 nm Pierce Protein Assay Kit (Thermo Scientific, USA) colorimetric assay method. All reactions were performed in triplicate.
HSL activity test
The lipase activity assay protocol (Sigma Aldrich) was used for the assay of the enzymatic activity of HSL. All the procedures performed will be according to the manufacturer's recommendation.
Statistical analysis
The data were presented as mean ± standard error of the mean. For analysis of more than two samples, analysis of variance (one-way analysis of variance) followed by Tukey's post hoc verification was applied. Statistically significant differences were considered where the p value was ≤0.05.
Results
The 3T3-L1 cells initially presented a flattened characteristic when adhered to the plaque (Fig. 1A). It is possible to verify the phenotype change after the differentiation (Fig. 1B), similar to the adipocytes structure in culture medium. 14

3T3-L1 cells cultured in Dulbecco's modified Eagle's medium and 10% fetal bovine serum.
Three doses of total energy of an infrared laser (904 nm) were applied to the differentiated cells (1, 2, and 3J). After 4 h of incubation period, TAG levels were measured in the culture medium and HSL activity. As can be seen in Fig. 2, there were no differences in TAG release to the medium.
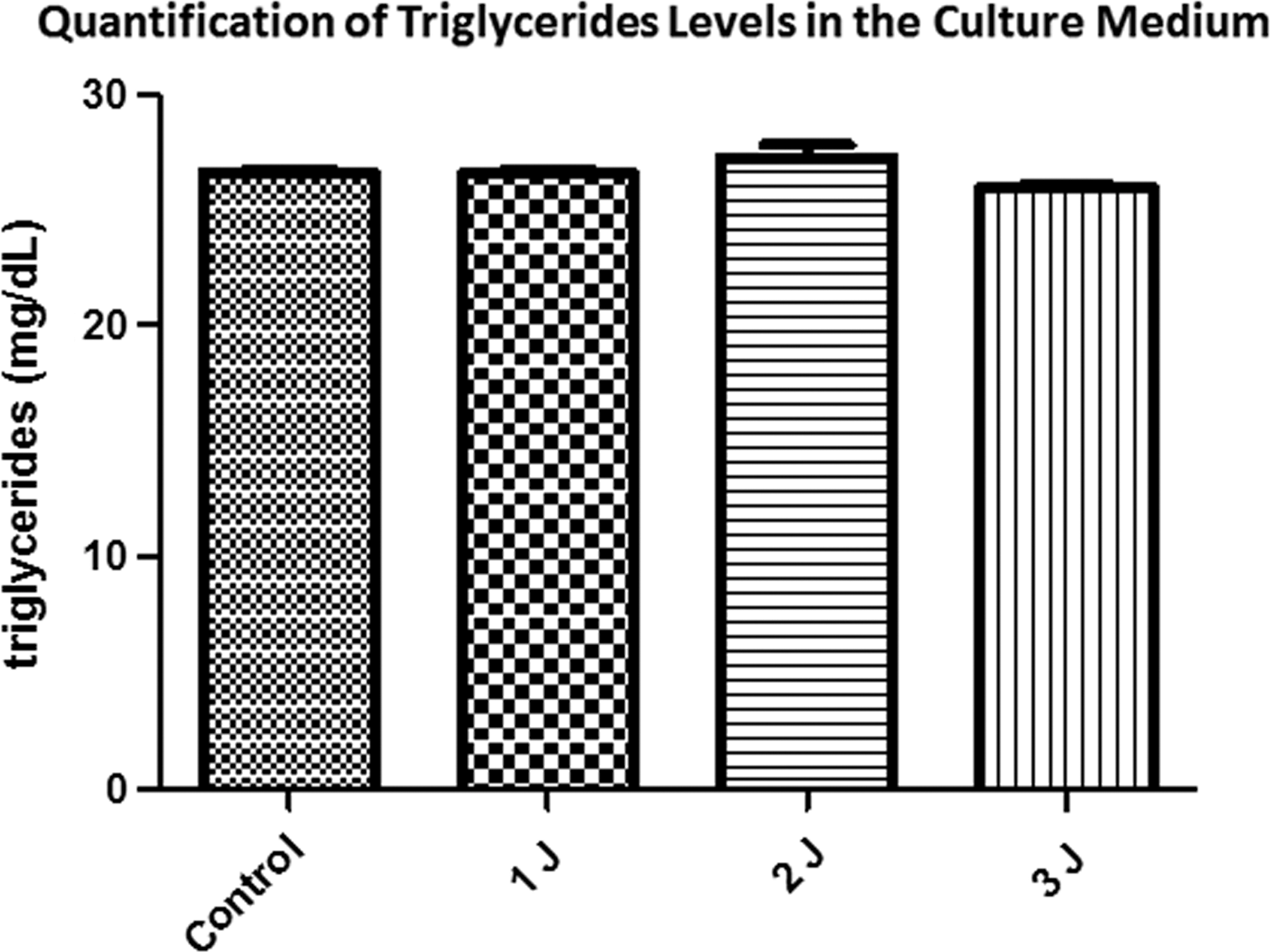
Triglycerides quantification from the 3T3-L1 adipocyte differentiated cells culture medium after treatment with different laser doses. Control = 26.5856 ± 0.52; 1J = 26.5856 ± 0.52; 2J = 27.2372 ± 1.41; 3J = 25.9991 ± 0.1303; p = 0.18.
For HSL activity, the calibration curve was used to determine the molarity of the glycerol levels (Fig. 3A), and it was observed that the laser decreased the activity of HSL in all doses applied (Fig. 3B).
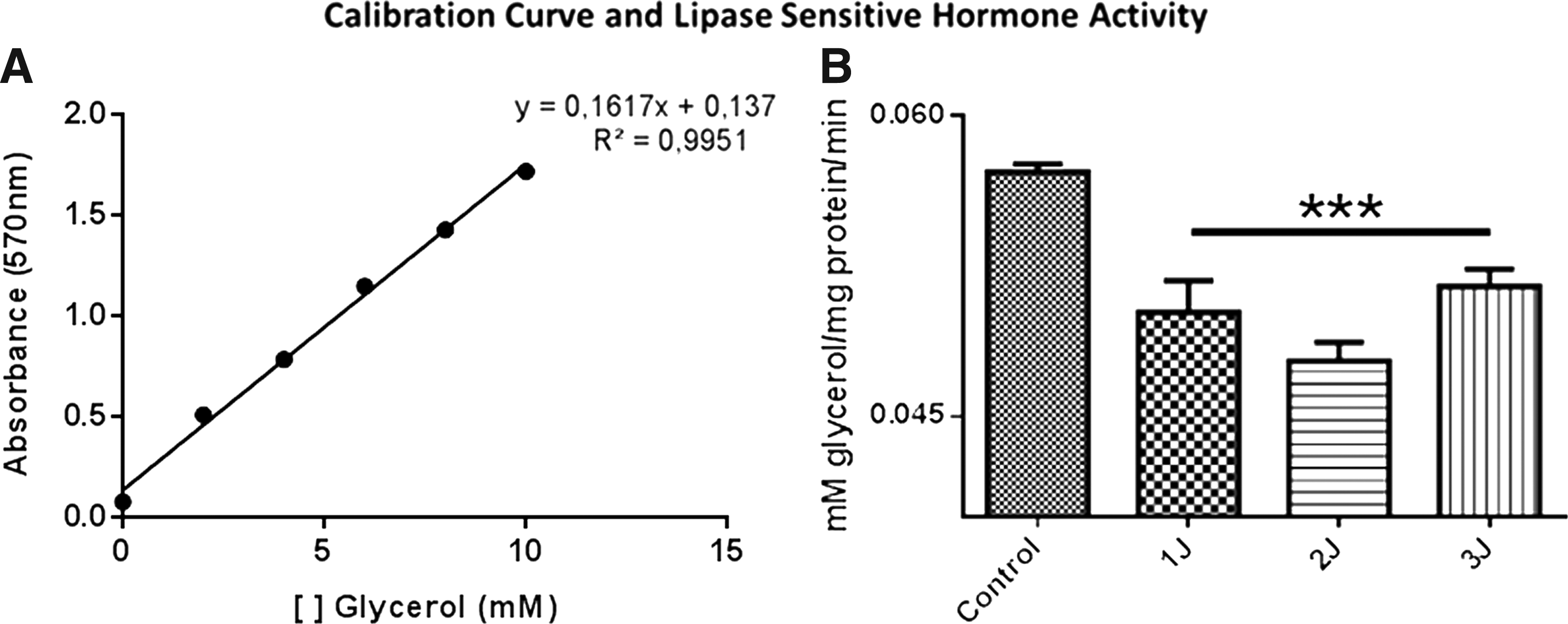
HSL assay of the 3T3-L1 adipocyte differentiated cells submitted to different laser doses.
Discussion
The lipid metabolism regulation is fundamental for energetic maintenance state of the organism, both for energy stock, as well as to impart highly energetic carbons to energetic oxidative metabolism. 15 The energy supply in adipose tissue follows an extensive network of anabolic cell signaling, such as insulin response, inducing the production of sterol regulatory element-binding protein (SREBP-1c) 16 with concomitant activation of acetyl CoA carboxylase 1. 17 Regarding catabolic responses, beta adrenergic signaling in adipose tissue triggers the activation of lipolytic components, including the HSL. 18
We demonstrated for the first time that laser photobiomodulation was able to significantly inhibit the HSL activity in 3T3-L1 cells. As HSL is the key enzyme for the release of fatty acids present in the adipose tissue for circulation, 19 we believe that the response of laser photobiomodulation may play a significant role in the catabolic processes of adipose tissue. Despite a decrease in HSL activity, we did not verify changes in the culture medium fatty acids release in all energy doses. Probably, the 4-h time course may be too small to observe the variation in the culture medium fatty acids release.
According to our results, the laser photobiomodulation promoted an inhibitory effect on HSL activity in 3T3-L1 cells. In some cases, laser photobiomodulation may or may not be able to trigger biological effects at different wavelengths. 20 –22 The standardization of the total energy amounts and the wavelength of the laser used allow a real understanding of the effects of photobiomodulation on the tissues.
The HSL negative modulation directly impacts on lipid metabolism. 19 According to our results, photobiomodulation can trigger anticatabolic responses in 3T3-L1 cells due to downregulation the HSL activity. However, these anticatabolic responses can be derived from the activation of the anabolic processes of lipid synthesis and fat mobilization into adipose tissue. This metabolic condition can imply many physiological effects for certain diseases. In anorexia nervosa, the metabolic inability of adipose tissue to perform its anabolic maintenance can trigger many organic problems. 23 Therefore, according to our results, it created a new field for the performance of laser photobiomodulation in lipodystrophic diseases, for example.
As a conclusion, the laser photobiomodulation responses trigger significant responses in HSL inhibition. These preliminary data open an extensive field of analysis to verify the possible effects of laser photobiomodulation on metabolic responses in adipose cells. In future studies, we will carry out tests in animal models to verify the occurrence of this phenotype.
Footnotes
Acknowledgments
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP N° 2015/06502-3).
Author Disclosure Statement
The authors declare no conflict of interests.