
Abstract
Objective:
The purpose of this study was to investigate the possible cytotoxic and genotoxic impact of new-generation 206 nm femtosecond solid-state laser irradiation on murine skin cells in vitro, and to compare the cell and DNA damage caused by different wavelength (206 vs. 257 nm) femtosecond laser pulses.
Background data:
The first attempts to evaluate the possible genotoxic impact of ultrashort laser pulses on the murine bone marrow cells in vitro revealed the unlooked-for DNA-damaging effect. However, the impact of far-ultraviolet (UV) radiation on genetic material of internal and external organs' cells may differ due to differences in size, structure, and biochemical composition of the cells.
Methods:
Mouse skin cells were exposed to different doses of 206 and 257 nm wavelength femtosecond laser, and 254 nm UV lamp irradiation. Comet assay in two versions—the standard alkaline and the enzyme-linked—was used for the evaluation of DNA damage.
Results:
The irradiation determined by different parameters demonstrated intensity-dependent genotoxic impact. The pyrimidine dimers made up the greater part of DNA photodamage, but with rising exposure dose the increase of relative amount of more energy-consuming primary damage—DNA strand breaks—was detected.
Conclusions:
The 206 nm femtosecond laser irradiation was much more cytotoxic but caused less primary DNA damage than the same pulse duration longer wavelength (257 nm) laser irradiation. DNA-damaging effect of 206 nm femtosecond laser pulses with extremely low penetration force may highly depend on the size, structure, and biochemical composition of the cells of organ or tissue targets.
Introduction
The rapid advance in technology of ultrashort pulsed lasers and their properties as a treating and surgical tool that allow to achieve desired result more quickly, precisely, and safer compared with longer pulsed picosecond or nanosecond lasers determined that femtosecond lasers are increasingly applied in such medical disciplines as ophthalmology, dermatology, dentistry, otology, and some others. 1 –5 The tentative testing of ultrashort pulsed lasers demonstrated that new-generation ultraviolet (UV) femtosecond solid-state lasers could be a progressive change in corrective surgery of surface organs requiring very precise ablation procedures. For example, the results of corneal stromal ablation with femtosecond UV pulses in rabbits were comparable with or superior to those obtained using argon fluoride excimer laser. 5 However, a substantial advantage of the new-generation femtosecond laser, PHAROS, is the possibility of generating femtosecond light pulses in both the infrared and UV ranges, implying that a wide range of ophthalmic or dermatological procedures can be performed using a single solid-state laser device. 5 –7
Although employment of UV femtosecond lasers in microsurgery opens new possibilities, the impact of their use on genetic material is not yet fully known and investigated. Our first attempts to evaluate the possible genotoxic impact of the brand-new 205 nm femtosecond laser, PHAROS, on the murine bone marrow cells in vitro revealed the unlooked-for DNA-damaging effect. 8,9 However, the impact of the same parameters' radiation on genetic material of internal and external organs' cells may differ due to differences in structure, biochemical composition, and susceptibility to UV radiation of the cells. 10 –12 So the purpose of the present study was to extend our previous research and evaluate the possible genotoxic effect of the femtosecond laser, PHAROS, on cells of such surface organ as skin. In our previous investigation we detected that the DNA damage induced depended mainly upon wavelength rather than on other parameters, such as pulse duration, repetition rate, or even beam delivery to a target. 9 Therefore, in our current work, we compared the DNA-damaging effect of different wavelength (206 and 257 nm) femtosecond laser pulses. Comet assay in two versions—the standard alkaline and the enzyme-linked—was used for DNA damage measurement. Comet assay in standard alkaline version reveals the presence of single-strand breaks, double-strand breaks, and alkali-labile sites, 13 –15 whereas in the enzyme-linked version specific UV-induced damage—cyclobutane pyrimidine dimers—additionally is detected. 14,16,17 The use of both versions of comet assay allowed us to evaluate more accurately genotoxic impact of ultrashort UV laser pulses on the genetic material of cells.
Materials and Methods
Animals
Skin cells were isolated from BALB/C mice 1-day-old pups. The animals from the Animal Facility Institute of Immunology, Vilnius University (Lithuania) were used in the current work. Ethics approval (No. 0212) was given by the Committee of Lithuanian Animal Care and Use.
Isolation and culture of murine skin cells
The pups of mice were euthanized, a piece of dorsal skin was cut off, and immersed in the transfer medium with antibiotics for a few minutes. The tissue was then mechanically chopped fine in the sterile glass and 5 mL of Iscove's modified Dulbecco's medium (Sigma-Aldrich, St. Louis, MO) enriched with 10% fetal bovine serum (Life Technologies, Carlsbad, CA) and antibiotics was added. The content of glass was transferred into a flask (25 cm 2 ), thoroughly pipetted, and cultured in a 37°C humidified incubator in a 5% CO2 atmosphere. Within 2 weeks a monolayer of cells on the bottom of the flask formed and primary culture of cells was obtained. When the density of cells in the monolayer reached 80–90%, the cells were reseeded: the old culture medium was poured off, the monolayer was washed with 4 mL 0.25% tripsin–EDTA solution (Biological Industries, Beit Haemek, Israel), then 0.5 mL of the same solution was poured on the monolayer, and the flask was incubated 5–7 min at 37°C humidified, containing 5% CO2 atmosphere. When the monolayer dispersed into separate cells, 10 mL of new culture medium was added, cell suspension resuspended, and in equal parts transferred into two new flasks.
Preparation of cell suspension for irradiation
Cells of third–sixth passages were used for UV laser and lamp irradiation. The dispersion of monolayer was performed in the same way as it was done for the cells' reseeding. The separated cells were vacuumed and mixed with 5–7 mL phosphate-buffered saline solution (PBS; Biochrom AG, Berlin, Germany). After centrifugation at 1500 rpm for 5 min, the resulting cell pellet was resuspended and PBS gradually added until the number of cells per milliliter of suspension reached 50,000. The absorbance of irradiation light by such density cells' suspension is quite similar to that of PBS (Fig. 1) and the number of cells is sufficient for comet assay performance.
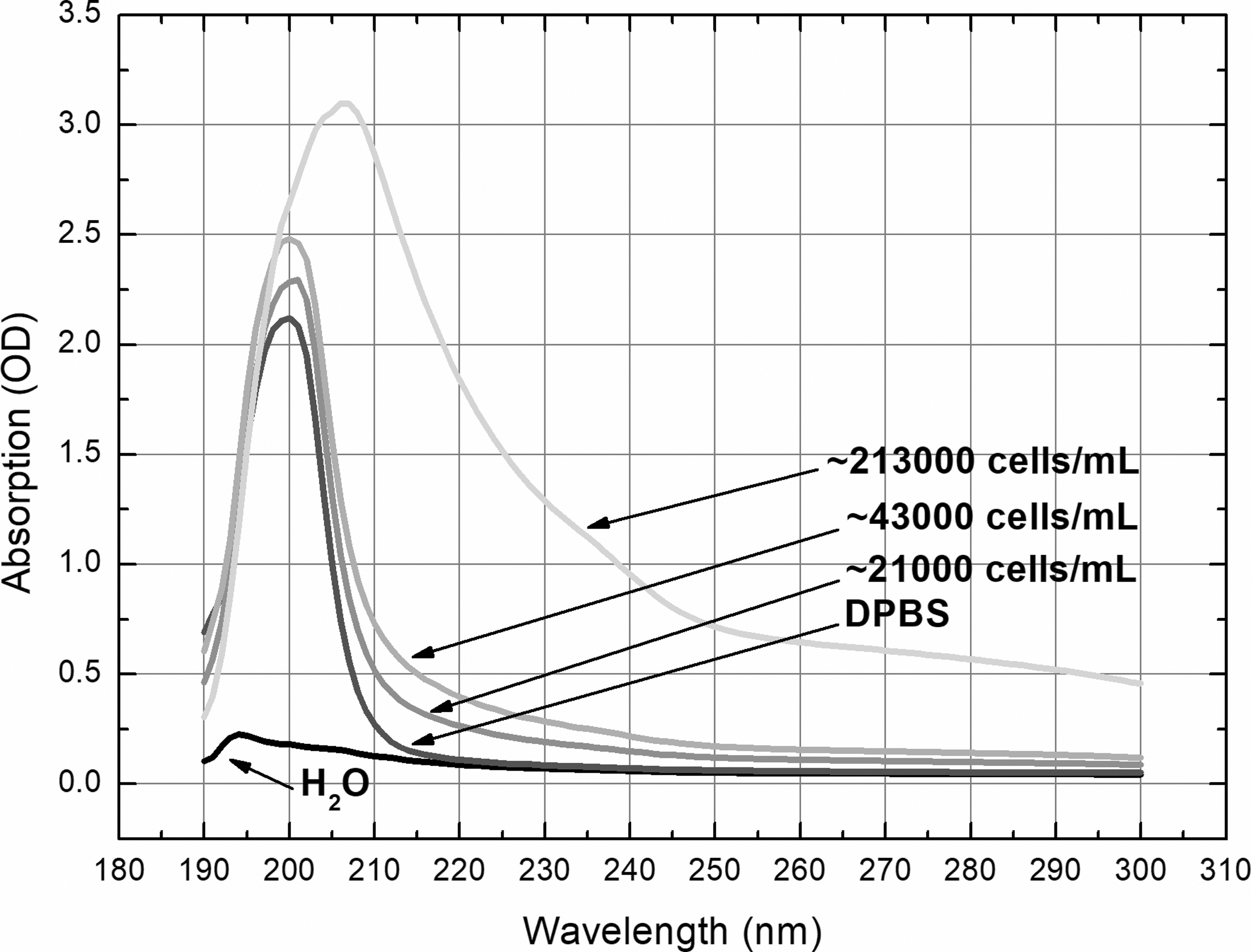
Absorption spectra of H2O, DPBS, and suspension with different number of skin cells in 1 mL DPBS. The absorbance of irradiation light depended on the density of cell suspension and was much stronger in deep-UV range. DPBS, Dulbecco's phosphate-buffered saline solution; UV, ultraviolet.
Laser irradiation
The fifth harmonic of the solid-state Yb:KGW laser, PHAROS (Light Conversion, Ltd., Vilnius, Lithuania), was used for generating two different wavelength (206 and 257 nm) femtosecond radiations. For positive control, we used the 254 nm line of mercury discharge lamp (Carl Roth, GmbH & Co. KG, Karlsruhe, Germany) because this type of radiation is known for having a strong DNA-damaging impact. 17 –19 Parameters (beam diameter and pulse energy) of the laser were adjusted to achieve a constant energy density at different irradiation doses: 9.5 and 0.03 μJ/cm 2 for 206 and 257 nm, respectively. The energy parameters of irradiation are summarized in Table 1. The exposure time for radiation sources was calculated by dividing the dose by the average density of irradiation power.
Parameters of Ultraviolet Light Sources Used for Irradiation of Skin Cells
UV, ultraviolet.
In dose–response experiments, it is important to ensure uniform irradiation of the sample containing skin cells. For the femtosecond pulses of 206 nm, the desired energy density per pulse was achieved by tuning the beam diameter to 2.2 mm. To ensure uniform sample irradiation, the laser beam was delivered over the sample using galvanometric scanners HurrySCAN 10 (SCANLAB, Puchheim/Munich, Germany). In this way, the high repetition rate of 206 nm femtosecond pulses were evenly spread out over the entire sample area by scanning a circle structure. For the femtosecond pulses of 257 nm, the laser beam was expanded with a single spherical lens. In front of the lens the stainless steel aperture was used. By adjusting the diameter of the aperture the desired energy density was achieved.
The average power of both pulsed and continuous UV light sources in the central part of the irradiated area was measured using power meter Nova II with thermal power sensor 3A (Ophir Optronics, Inc., Jerusalem, Israel). The active area of the power sensor was ∼0.7 cm 2 (smaller than the irradiated area). The irradiation power density was determined by dividing measured power value by the active area of the power sensor. Spectrophotometer UV-3101PC (Shimadzu, Tokyo, Japan) was used to measure the UV absorption spectra.
Treatment and preparation protocol
Fifty-four microplates were divided into 3 groups of 18 microplates and each of these groups was further subdivided into 3 subgroups of 6 microplates. Then a 45 μL aliquot of prepared skin cell suspension was transferred onto each of the microplates and spread over an area of 1 × 1 cm to irradiate by two different wavelengths (206 and 257 nm) femtosecond laser and 254 nm continuous lamp light. To evaluate the possible effect of medium on the extent of DNA damage, the absorbance of irradiation light by distilled water, PBS, and cell suspension was measured (Fig. 1). Skin cells on separate plates of each group were exposed to irradiation of different parameters at six doses—0.000175, 0.00035, 0.00175, 0.0035, 0.0175, and 0.035 J/cm 2 . Immediately after irradiation, the suspension from each microplate was picked up and transferred into separate Eppendorf microtubes, which were kept for several minutes (until further procedures were started) in the dark at 4°C. Simultaneously, a 40 μL aliquot of unexposed suspension (control) was dropped into five additional microtubes, which were kept under the same conditions as those containing affected samples. The samples of one of subgroups were used to assess cell viability by Trypan Blue exclusion assay and dual Acridine Orange/Ethidium Bromide (AO/EB) staining test, whereas samples of the two other subgroups—to measure the amount of DNA damage by standard alkaline and alkaline enzyme-linked comet assays.
Trypan blue exclusion assay
Ten microliters of each sample suspension was transferred from microtubes onto a microscope slide and mixed with 10 μL of 0.4% Trypan Blue (Sigma-Aldrich). After 1 min a cover glass was placed on top and microscopic analysis was performed. At least 100 cells on each slide per treatment were scored to determine the percentage of viable and dead cells.
Dual AO/EB staining test
Twenty microliters of each sample suspension was transferred from microtubes onto a microscope slide, mixed with 4 μL of AO/EB staining solution (Sigma-Aldrich), and then covered with a coverslip. At least 100 cells on each slide per treatment were scored to determine the percentage of living undamaged, early stage apoptotic, late-stage apoptotic, and necrotic cells.
Standard alkaline comet assay
A 40 μL aliquot of 1% low melting point agarose (Carl Roth GmbH & Co. KG) in PBS, warmed up to 50°C, was dropped in turns into each of the 18 microtubes containing irradiated samples of suspension and into a microtube with unexposed (control) sample suspension. The content of each microtube was quickly mixed and transferred onto a microscope slide precoated with 1% standard agarose (Thermo Fisher Scientific, Vilnius, Lithuania) in PBS, and a cover glass was placed on top. The further procedures were performed according to the protocol described by Singh et al. 13 and Collins, 14 but with some modification. 8,9 The analysis of the slides was performed the next day and 100 cells on each slide per treatment were scored. The computerized image analysis system, Lucia (Laboratory Imaging Ltd., Praha, Czech Republic), was used to determine the percentage of DNA (%DNA) in the comet tail. Each experiment was performed five times.
Enzyme-linked alkaline comet assay
The insertion of cells into agarose and theirs transfer from the microtubes onto the slides was performed in the same way as described above, but for enzyme-linked comet assay, three slides with unexposed cells for control were made. The only and essential difference in procedures was the treatment of DNA of cells embedded in agarose with T4 endonuclease V (T4 PDG V, 10,000 U/mL; New England Biolabs, Ipswich, MA). For that all slides with affected suspension samples and two slides with nonirradiated suspension samples (control) after lysis procedure were washed with cold enzymatic buffer (100 mM NaCl, 40 mM Hepes, 1 mM Na2EDTA, 0.2 mg/mL bovine serum albumin) for 5 min, three times; a slide with the third control sample was washed the same time with cold PBS. Then, all slides with affected suspension samples and one of the first two control slides (T4 PDG V control) were treated with 50 μL of T4 PDG V solution (1 μg/mL in enzymatic buffer) for 30 min at 37°C in a humidified chamber. Simultaneously, on the remaining control slides, one of which was previously washed with enzymatic buffer, and the other one with PBS, 50 μL of enzymatic buffer (ENB control) and PBS (PBS control) were dropped, respectively. Both slides were then placed together with other slides for 30 min at 37°C in a humidified chamber. After incubation, all slides were transferred into alkaline electrophoresis solution for DNA uncoiling and the same procedures as described for standard comet assay were then performed.
Statistical analysis
Data were processed using the InStat V2.02 (GraphPad Software, CA) statistical package. Statistical analysis was performed using unpaired Student's t-test. The mean and standard error of the mean of the average of the DNA damage score of 5 gels per experimental condition were calculated. Pearson's determination coefficient (R 2 ) was used to evaluate the dose–effect relationship. p < 0.05 was chosen as the level of significance.
Results
The effect of different wavelength (206 and 257 nm) femtosecond lasers and 254 nm continuous lamp irradiation on the viability of skin cells assessed by the Trypan Blue exclusion is presented in Table 2 and Fig. 2. The data show that 206 nm femtosecond irradiation was most cytotoxic and significantly decreased the viability of cells at all doses used, except the lowest one. This type of irradiation at two greatest doses (0.0175 and 0.035 J/cm 2 ) decreased the number of viable cells to about 23% and 42%, respectively, whereas the same-intensity irradiation of both longer wavelengths—to only about 10%. The shortest wavelength irradiation showed the strongest dose–response dependence as well (R 2 = 0.98, R 2 = 0.60, and R 2 = 0.71 for 206, 257, and 254 nm irradiation, respectively).
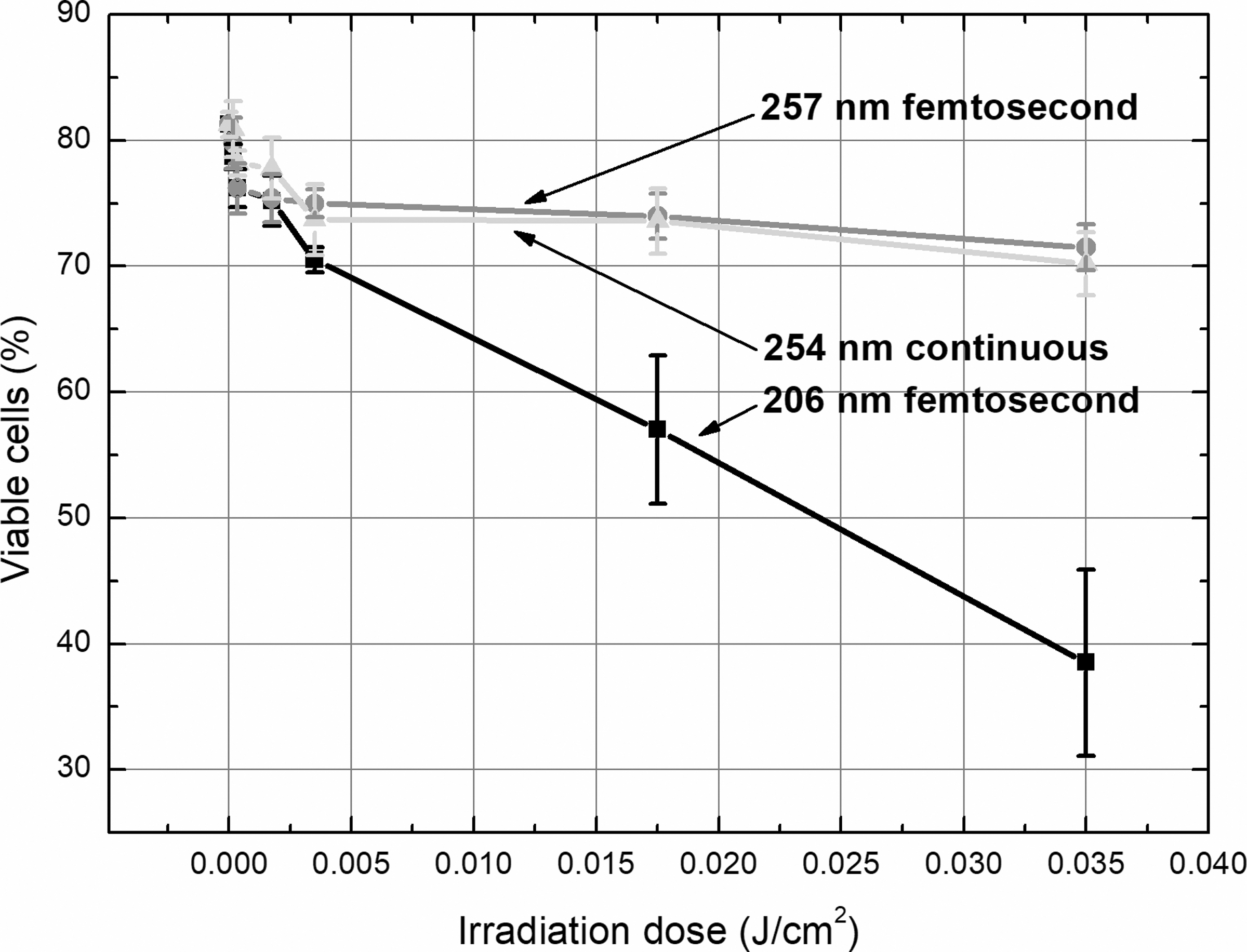
Viability of skin cells exposed to 206 and 257 nm femtosecond laser and 254 nm continuous lamp irradiation assessed by the Trypan Blue exclusion. Two hundred six nanometer femtosecond pulses were most cytotoxic as compared with longer wavelength both pulsed and continuous UV irradiation. UV, ultraviolet.
Percentage Amount (Mean Value ± Standard Error of the Mean) of Viable Skin Cells Exposed to 206 and 257 nm Femtosecond Laser, and 254 nm Continuous Lamp Irradiation at Different Doses Assessed by the Trypan Blue Exclusion
p < 0.0001, b p < 0.001, c p < 0.005, d p < 0.01, e p < 0.05 when compared with unirradiated control.
UV, ultraviolet.
Dual AO/EB staining test showed very similar impact of irradiation on viability of cells as Trypan Blue exclusion test when early stage apoptotic cells were classified as living cells. The proportion of cells of four distinguished groups in different irradiated cell suspension identified by AO/EB staining test is presented in Fig. 3. The lower doses of irradiation caused more often apoptotic death of cells, whereas the part of necrotic cells rose as dose of irradiation increased, and this trend was most pronounced in cell suspension exposed to 206 nm pulsed irradiation.
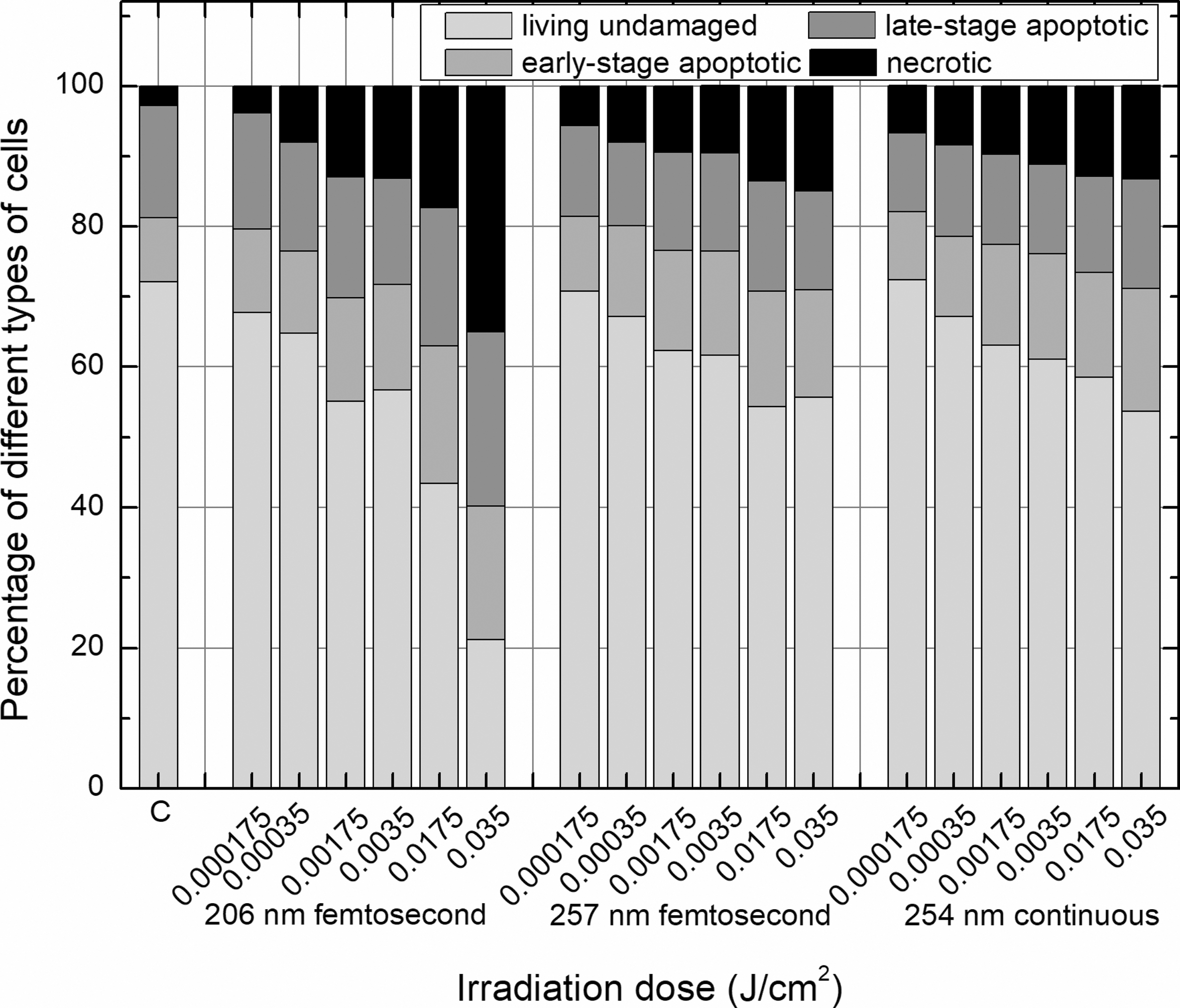
The percentage amount of living undamaged, different-stage apoptotic and necrotic cells, assessed by dual AO/EB staining test in skin cell suspension exposed to 206 and 257 nm femtosecond laser and 254 nm continuous lamp irradiation. The percentage of necrotic cells increased after exposure to higher irradiation doses, and this trend was most pronounced in cell suspension exposed to 206 nm pulsed irradiation. AO/EB, Acridine Orange/Ethidium Bromide.
The effect of different parameters of irradiation on DNA damage in skin cells assessed by standard alkaline comet assay is presented in Table 3 and Fig. 4. As can be seen, the 206 nm pulsed irradiation was the least genotoxic: four of the six doses used caused a significant increase in the amount of DNA in the comet tail, whereas five doses of the 254 nm continuous and six doses of the 257 nm pulsed irradiation had such effect. It should be noted that substantial increase in DNA damage after exposure at the greatest dose of 206 nm pulsed irradiation was due in the larger part to cytotoxic impact, because the viability of cells irradiated at this dose decreased by more than 40%. The strong or very strong dose–effect dependence was ascertained for all types of irradiation: R 2 values for 206 nm (excluding the highest dose of exposure), 257, and 254 nm irradiation were 0.98, 0.88, and 0.96, respectively.

The DNA-damaging effect of 206 and 257 nm femtosecond laser and 254 nm continuous lamp irradiation assessed by standard alkaline comet assay in skin cells. Two hundred six nanometer femtosecond pulses caused less DNA strand breaks as compared with longer wavelength both pulsed and continuous UV irradiation. UV, ultraviolet.
Percentage Amount (Mean Value ± Standard Error of the Mean) of DNA in the Comet Tail of Skin Cells Exposed to 206 and 257 nm Femtosecond Laser, and 254 nm Continuous Lamp Irradiation at Different Doses Assessed by Standard Alkaline Comet Assay
p < 0.0001, b p < 0.0005, c p < 0.001 when compared with unirradiated control.
UV, ultraviolet.
The enzyme-linked comet assay, which extended the range of detected DNA damage with pyrimidine dimers, revealed a much stronger genotoxic impact of irradiation than standard alkaline comet assay (Table 4 and Fig. 5). All five doses of different parameters' UV irradiation (the greatest 0.035 J/cm 2 dose was not used for DNA damage measurement by enzyme-linked comet assay) caused a significant increase in the amount of DNA in the comet tail. The 254 nm continuous irradiation showed the strongest damaging effect on DNA, 206 nm femtosecond pulses—the weakest. The dose–effect dependence for all types of irradiation was obvious but weaker than estimated using standard alkaline comet assay: R 2 values for 206, 257, and 254 nm irradiation were 0.86, 0.66, and 0.61, respectively.
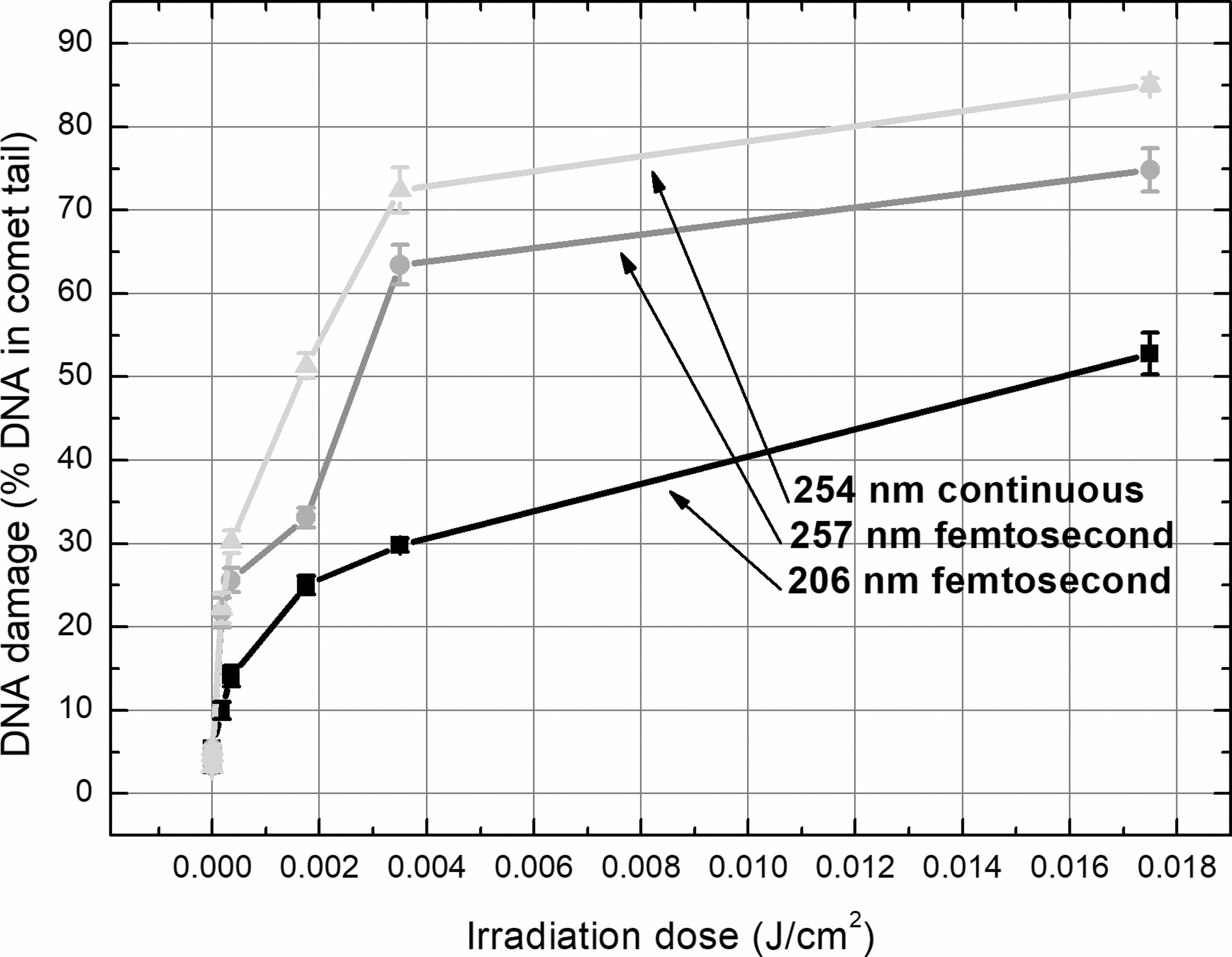
The DNA-damaging effect of 206 and 257 nm femtosecond laser and 254 nm continuous lamp irradiation assessed by enzyme-linked alkaline comet assay in skin cells. The amount of photodamage (strand breaks plus cyclobutane pyrimidine dimers) in skin cells was lower after exposure of cell suspension to 206 nm femtosecond pulses than caused by longer wavelength both pulsed and continuous UV irradiation. UV, ultraviolet.
Percentage Amount (Mean Value ± Standard Error of the Mean) of DNA in the Comet Tail of Skin Cells Exposed to 206 and 257 nm Femtosecond Laser, and 254 nm Continuous Lamp Irradiation at Different Doses Assessed by Enzyme-Linked Alkaline Comet assay
p < 0.0001, b p < 0.005 when compared with unirradiated T4 PDG V control.
EB, enzymatic buffer; PBS, phosphate-buffered saline solution; UV, ultraviolet.
Discussion
In the current study, we investigated the cytotoxic and genotoxic effect on skin cells of low- and very-low-intensity ultrashort UV pulses. The greatest and the lowest irradiation doses we used were ∼7 and 1300 times lower, respectively, than analogous clinical parameters. 5,20,21 Choosing a spectrum of low irradiation doses in which the fringe exposure doses differed from each other by 200 times, we simulated a situation that could not be completely avoided when a laser as a treating or surgical tool is used. Although during the laser procedure the energy of irradiation is directed to the cells of target area, a small part of the photons can be scattered and absorbed by the cells surrounding the target. The DNA photodamage could lead to genetic disorders, if cells containing damaged genetic material survived and continued the cell cycle. For this reason, risk of mutation and, therefore, malignancy is the lowest in cases in which laser irradiation is more cytotoxic and induces less primary damage to genetic material.
Our study demonstrated that cytotoxic effect of radiation was mainly dependent on wavelength: radiation of the shortest investigated wavelength was able to cause more irreversible lesions in membrane and maybe in other superficial structures of cells, because of the greater energy of photons in the shorter wavelength range. The 206 nm pulses caused death of cells due to necrosis more often as compared with radiation of both longer wavelengths and this trend was most pronounced after exposure of cells by the greatest irradiation doses. The shift of apoptotic and necrotic cells' ratio toward growing of part of necrotic cells was indicated with increasing of irradiation's intensity for longer wavelength UVB irradiation as well. 22,23
The effect on genetic material of 206 nm femtosecond irradiation assessed by both—standard alkaline and enzyme-linked—comet assays was weaker than DNA-damaging impact of 257 nm pulsed and 254 nm continuous irradiation. However, although the genotoxic effect of radiation was mostly dependent on the wavelength, the influence of such parameter as beam delivery to a target on the ratio of different types of DNA damage was revealed: pulsed irradiation of both wavelengths (206 and 257 nm) induced more DNA strand breaks and less pyrimidine dimers as compared with continuous radiation (Fig. 6). It should be noted that cyclobutane pyrimidine dimers made up the larger part of primary damage of genetic material in cells exposed to irradiation of all parameters used, although the proportion of this type of lesions decreased as intensity of irradiation rose. For example, the percentage of pyrimidine dimers in cells exposed to 206 and 257 nm femtosecond and 254 nm continuous irradiation at the lowest dose and the fifth largest dose made up 95.8%, 94.5%, 94.7% and 73.6%, 59.4%, 79.1% of all lesions, respectively. This relative change in the amount of different DNA damage showed growing potency of irradiation with increasing exposure doses to cause more energy-consuming primary lesions of genetic material, such as single- and double-strand breaks. 24 –27
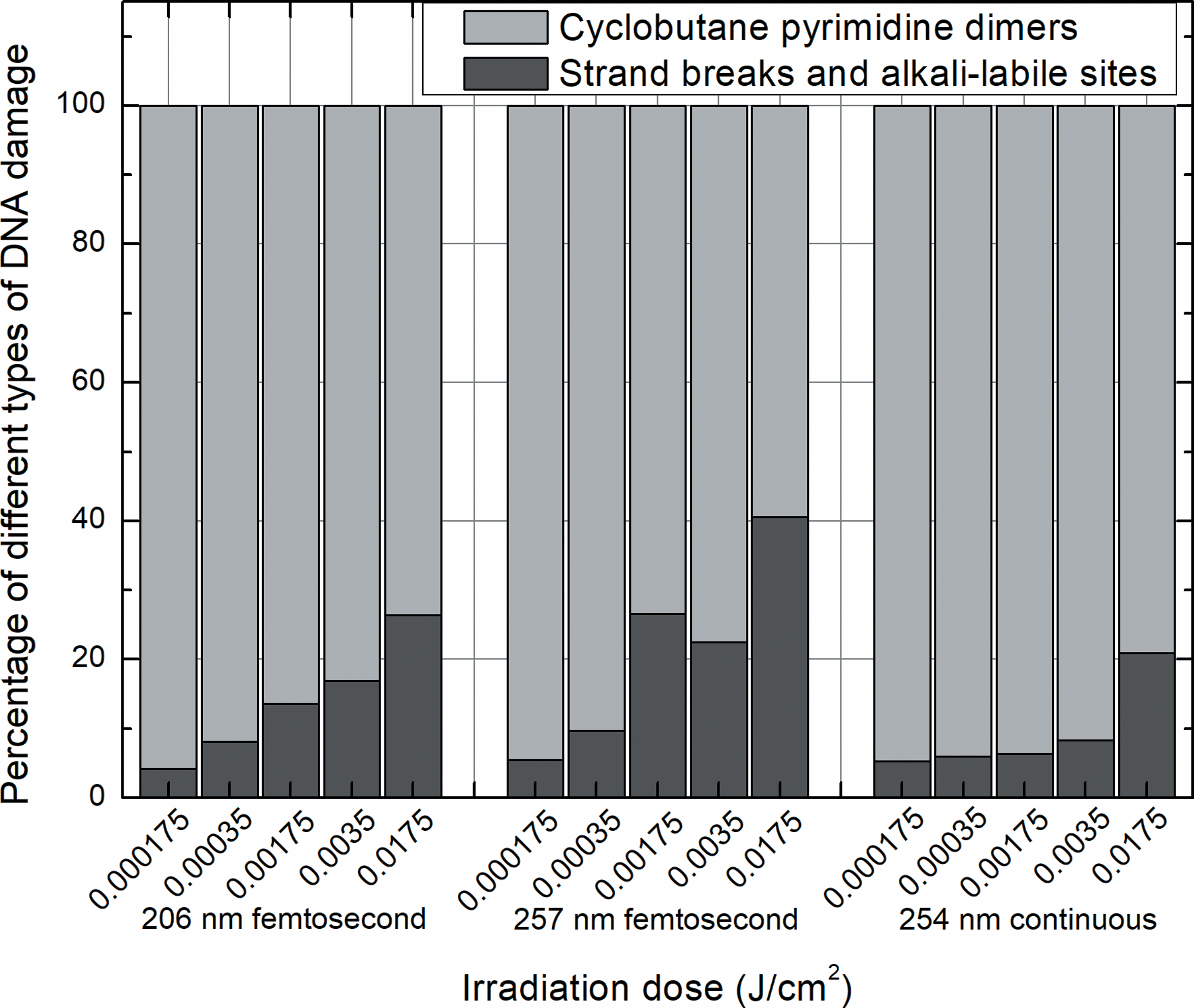
The percentage amount of different types of DNA photodamage in skin cells exposed to 206 and 257 nm femtosecond laser and 254 nm continuous lamp irradiation at different doses. The percentage of pyrimidine dimers was the greatest in cells exposed to UV irradiation of different parameters, but with increasing intensity of radiation the percentage of strand breaks and alkali-labile sites rose. UV, ultraviolet.
Therefore, in our study, 206 nm femtosecond pulses showed the strongest cytotoxic and the weakest genotoxic impact as compared with longer wavelength pulsed (257 nm) and continuous (254 nm) radiation. These features could be considered as the advantages of 206 nm femtosecond laser pulses when applying UV radiation for medical use because of reducing the risk of the formation of mutations and tumors in irradiated tissues or organs. However, it is necessary to note that DNA-damaging effect of far-UV radiation, thus of 206 nm wavelength radiation as well, may highly depend on the size, structure, and biochemical composition of the cells, because of the low penetration power and high energy of rays in this UV range at the same time. 3,10,28 –31 The radiation of deep-UV range will hardly reach the nucleus surrounded by a comparatively thick layer of cytoplasm in a cell with a few dozen micrometers in diameter, therefore DNA-damaging effect will be limited. The filtering effect of thin cytoplasmic layer of a few times smaller cell may be not sufficient to completely prevent far-UV radiation from reaching the nuclear DNA. In that case, the damage of genetic material may be of larger extent than caused by longer wavelength UV rays, because of the greater energy of photons in the shorter wavelength range. In our previous studio, 9 shorter wavelength irradiation (205 and 213 nm vs. 254 and 266 nm) showed a 2–3.5 times stronger genotoxic impact to bone marrow cells, the size of which on the average reached only 7.6 μm, whereas the average size of skin cells investigated in the current work was 35.9 μm. Therefore, along with parameters of laser irradiation, the size of cells of organ- or tissue targets is no less an important circumstance when considering the risk of damage of genetic material due to radiation of deep-UV range.
Conclusions
Our investigation showed that the harmful impact of different wavelength (206 and 257 nm) femtosecond laser pulses on murine skin cells in vitro was different. The 206 nm radiation was much more cytotoxic but caused less primary DNA damage than 257 nm pulsed and 254 nm continuous radiation as well. Cyclobutane pyrimidine dimers were the prevailing form of DNA photodamage in irradiated cells regardless of the parameters of radiation, but the proportion of this form of damage declined and simultaneously the number of DNA strand breaks increased as intensity of irradiation rose. DNA-damaging effect of 206 nm femtosecond laser pulses with extremely low penetration force may highly depend on the size, structure, and biochemical composition of the cells of organ or tissue targets.
Footnotes
Acknowledgments
The authors sincerely thank associate professor Dr. Virginija Bukelskiene for kind consulting, valuable advice, and technical assistance in this work.
Author Disclosure Statement
Ms. Gabryte Butkiene, Mr. Danielius, and Mr. Sobutas are paid employees of Light Conversion Ltd. Mr. Danielius is a shareholder of Light Conversion Ltd. None of the other authors has a financial or proprietary interest in any material or method mentioned.