
Abstract
Objective:
The aim of this study was to evaluate the effects of 940-nm diode laser irradiation on proinflammatory cytokine secretions [interleukin (IL)-6 and IL-8] by human gingival fibroblasts in vitro.
Background:
Photobiomodulation has been routinely used in many dental procedures; however, the exact biological action mechanism of photobiomodulation and its therapeutic benefits have not been established.
Methods:
Cells derived from systemically healthy individuals were treated with three different laser parameters—6 J for 20 sec [0.84 J/cm2 (0.04 W/cm2)], 10 J for 20 sec [1.4 J/cm2 (0.07 W/cm2)], and 14 J for 20 sec [1.97 J/cm2 (0.09 W/cm2)]—in the presence and absence of 1 μg/mL lipopolysaccharide (LPS) stimulation. Laser irradiations were carried out by a 940-nm diode laser device in continuous pain therapy mode with a deep tissue handpiece. Changes in cell viability, cytokine secretions, and mitogen-activated protein kinase pathway expressions were investigated, and results were compared with negative (medium) and positive control (1 μg/mL LPS) groups. The data obtained were statistically analyzed by the Mann–Whitney U test for pairwise comparisons among groups at the 0.05 level of significance.
Results:
Laser therapy with 0.84–1.4 J/cm2 amplified IL-6 and IL-8 secretions, whereas 1.97 J/cm2 suppressed IL-6 and IL-8 release in LPS-stimulated cells. Cell viability did not show a variation with photobiomodulation.
Conclusions:
These results demonstrate that photobiomodulation can alter IL-6 and IL-8 release, with cytokine suppression potency at a relatively high dose, as demonstrated previously. However, in contrast, we found that a low level of stimulation (6 J) in the presence of inflammation (LPS stimulation) may further enhance IL-6 and IL-8 release. We also found that p38 and ERK1/2 pathways are activated by LPS as well as by photobiomodulation.
Introduction
Periodontal disease is a common and complex inflammatory disease that results in progressive destruction of the periodontium. 1 The pathogenesis of periodontal disease is mediated by inflammatory responses. 2,3 Various immune receptors act during inflammation process. Receptors recognize and detect pathogens and activate host cells to form host responses. 2,4
Probably the most central of these receptors are Toll-like receptors (TLRs). TLRs detect multiple pathogen-associated molecular patterns, including lipopolysaccharide (LPS), which is a glycolipid component of the outer membrane of Gram-negative bacteria. LPS can stimulate the innate immune system, leading to the production of inflammatory responses that activate adaptive immunity. Trace amounts of LPS can activate the innate immune system, leading to the production of an array of proinflammatory mediators, such as tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6). For this reason, LPS is generally acknowledged to play a central role in eliciting inflammatory responses. 4 –6
Raised levels of proinflammatory cytokines have been identified in the gingival crevicular fluid and saliva of patients with chronic periodontitis. 7 Immune responses to infection are regulated by cytokine and chemokine signals. These low-molecular weight proteins are involved in the initiation and further stages of inflammation, in which they regulate the amplitude and the duration of the responses. 2,3,8 IL-6 is a pleiotropic cytokine with a wide range of biological activities in immune regulation, hematopoiesis, inflammation, and oncogenesis. Chemokines are a type of cytokine that direct the migration of white blood cells to infected or damaged tissue. IL-8 is a potent chemoattractant chemokine for neutrophils and belongs to the CXC family. 4,6
Scaling and root planning also known as conventional periodontal therapy is an effective method to remove dental plaque and calculus deposits in periodontal disease. However, the technique may be unable to eliminate pathogenic microbes. Therapies such as photobiomodulation, high-intensity laser therapy, and antimicrobial photodynamic therapy have been proposed as adjuncts to scaling and root planing for the treatment of chronic periodontitis. 1,9,10
Photobiomodulation is the nonsurgical application of laser light to stimulate cell responses to promote tissue healing, reduce inflammation, and induce analgesia. 11 Photobiomodulation has been safely and efficiently applied in various dental procedures. 11 It stimulates many biological mechanisms, such as cell growth, proliferation, and differentiation. Proliferative effects of photobiomodulation have been demonstrated in various cell types in vitro. 12,13 Although it is known that photobiomodulation has encouraging effects on inflamed tissues, the underlying molecular mechanisms have not been fully established. 12 In a recent systematic review, Kellesarian et al. 9 investigated the effect of laser-assisted scaling and root planing on the expression of proliferative cytokines in patients with chronic periodontitis and reported that the role of laser-assisted scaling and root planing on the expression of proinflammatory cytokines remains unclear. Knowledge of how immune mechanisms and inflammatory responses are regulated is critical for understanding the pathogenesis of periodontitis and its treatment.
The aim of this study was to evaluate the effects of photobiomodulation on secretion of IL-6 and IL-8 in LPS-stimulated human gingival fibroblast cells. The null hypothesis was that photobiomodulation would have no influence on cytokine secretions on unstimulated or even LPS-stimulated gingival fibroblasts.
Materials and Methods
Cell isolation
Gingival fibroblast cells were isolated from human gingival connective tissue of systemically healthy young individuals (n = 5). All patients gave informed consent before providing the samples. The study protocol was approved by the Akdeniz University Clinical Research Ethics Committee (214/05.04.2017—Antalya, Turkey). The gingival samples were fragmented into small pieces with a scalpel and transferred to culture dishes containing Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich, Inc., St. Louis, MO) supplemented with 10% fetal calf serum and 100 μg/mL streptomycin (Sigma-Aldrich, Inc.). The tissue samples were transported to the Medical Pharmacology Laboratory. Cells were incubated in a humidified atmosphere of 5% CO2 at 37°C. The medium was changed the next day and twice in a week. After reaching ∼70% confluence, the cells were subcultured with 5 mL of 0.25% trypsin–0.53 mM ethylenediaminetetraacetic acid solution (EDTA; Gibco BRL, Gaithersburg, MD).
In all groups, gingival fibroblasts were used between the second and sixth passages. The plates were divided randomly into eight treatment groups. A set of untreated cells was used as control. To stimulate inflammatory conditions, 1 μg/mL Porphyromonas gingivalis LPS (InvivoGen, San Diego, CA) was used. Our preliminary tests with 1 μg/mL P. gingivalis strongly stimulated expressions of IL-6 and IL-8 without cytotoxicity. To avoid from a toxic dose, we used 1 μg/mL P. gingivalis LPS and we applied once a time. Details of the study design and groups are presented in Table 1.
Study Design
LPS, lipopolysaccharide; PBM, photobiomodulation.
Photobiomodulation
All laser irradiations were carried out by means of a diode laser device (Epic, Biolase, indium gallium arsenide phosphate, 940 nm—semiconductor diode, 940 nm) in continuous pain therapy mode with a deep tissue handpiece (Table 2). For photobiomodulation, six-well plates were used, and cells were plated as 30,000 cells/well. To prevent cross-irradiation, only three wells of the plates were used, and wells were irradiated at a single point. Irradiation was done in partially dark conditions without the influence of other light sources. During experiments, to avoid possible reflections, cover plates were removed, and the laser was irradiated from the base of the plates for 20 sec in contact mode. All procedures were performed under “clean bench” conditions to prevent bacterial contamination. Experiments reported here were repeated three times and representative results were demonstrated. Each experiment was performed using cells from different individuals. The control and nonirradiated LPS groups were maintained in identical conditions except that the laser device was switched off. Table 1 gives the parameters of photobiomodulation applications.
Parameters of Laser Irradiation
Cytokine detection
To analyze cytokine levels, we collected culture supernatants after 24, 48, and 72 h. Levels of IL-6, IL-8, and cytokines were detected by means of ELISA kits [R&D (IL-6) and BD Pharmingen, Franklin Lakes, NJ (IL-8)] according to the manufacturer's instructions with a 450-nm microplate reader (BioTek Instruments, Inc., Winooski, VT). Cytokine analysis initially was performed using 24-well plates in which 10,000 cells per well were seeded. Experiments were repeated using 96-well plates (3000 cells/well) for two more times. Representative results are given.
Cell viability assay
Cell viability was measured after 72 h by means of a colorimetric assay for 96-well plates with the WST-1 test (Roche Diagnostics, Mannheim, Germany). A 10% cell proliferation reagent was added to the culture medium of each well. After storage in an incubator at 37°C and 5% CO2 for 3–4 h, the absorbance was measured by means of a 450-nm microplate reader (BioTek Instruments, Inc.). Cells were plated as 3000 cells per well in a 96-well plate. Treatments were performed 24 h after plating. A standard curve was prepared to determine the number of cells for each OD measure.
Morphology of gingival fibroblasts
Cell morphology was examined microscopically. Images of cells were taken after 72 h by microscopy (Olympus Life Sciences, Japan).
Western blot analysis
The effects of LPS and photobiomodulation on phosphorylation of ERK1/2 and p38, Akt, were also studied (all antibodies were from Cell Signaling, Danvers, MA). Primary fibroblasts underwent lysis at the end of the experiments (after third stimulations), and phosphorylated forms of p38 and ERK1/2 were determined as markers for the activation of p38 and ERK1/2. For Western blot analysis, a six-well plate was used and cells were plated as 30,000 cells/well. Cells were collected using 300 μL of lysis buffer containing protease inhibitor for each well. A 50-μL quantity was taken from each group and mixed with 25 μL loading dye. The samples were then centrifuged at 7826 × g for 30 sec and incubated for 4 min. After centrifugation, a 10% polyacrylamide gel was prepared, and the samples were separated by electrophoresis at 120–150 V. At 40 V, gelatin polyvinylidene difluoride membrane proteins were transferred. Membranes were kept for 1 min in methanol and stained with Ponceau S for control. This was followed by the blocking step, whereby samples were incubated at room temperature for 2 h with 1 × Tbs-Tween20 (wash buffer) containing 5% milk powder. Membranes were incubated with primer antibodies overnight. After being washed with 1 × Tbs-Tween20 (3 × 10 min), membranes were incubated with a secondary antibody at room temperature for 2 min, and the washing step was repeated to complete the marking step. For imaging, membranes were incubated with the chemoluminescence agent for 4 min at room temperature and were inserted into the cassette. In the dark, the radiation on the membrane was transferred onto the film, and the washing tape became visible on the film.
Antibodies used were phospho-ERK1/2 (4370L; Cell Signaling), phospho-p38 (4511L; Cell Signaling), GAPDH (Meridian H86504), and horseradish peroxidase-conjugated secondary antibodies (1:3000; Santa Cruz).
Statistical analysis
All experiments were carried out in triplicate. Cell viability data were statistically analyzed by ANOVA. Interleukin results did not show a normal distribution and were analyzed by the Kruskal–Wallis test complemented by the Mann–Whitney test for pairwise comparisons at a 5% level of significance with SPSS for Windows version 16.0 (SPSS).
Results
Effects of LPS and/or photobiomodulation on IL-6 levels
Stimulation of gingival fibroblasts with LPS or laser irradiation alone significantly increased the levels of IL-6. Costimulation with LPS and laser somewhat counteracted their mutual effects. Specifically, LPS-induced increases in IL-6 levels were prevented by 10 and 14 J stimulation (Fig. 1A). Desensitization to the effects of 14 J stimulation on IL-6 release was also observed after the second and third stimulations. Costimulation with LPS and laser counteracted their mutual effects after the second and third stimulations, and was more prominent with 14 J stimulations (Fig. 1B, C).
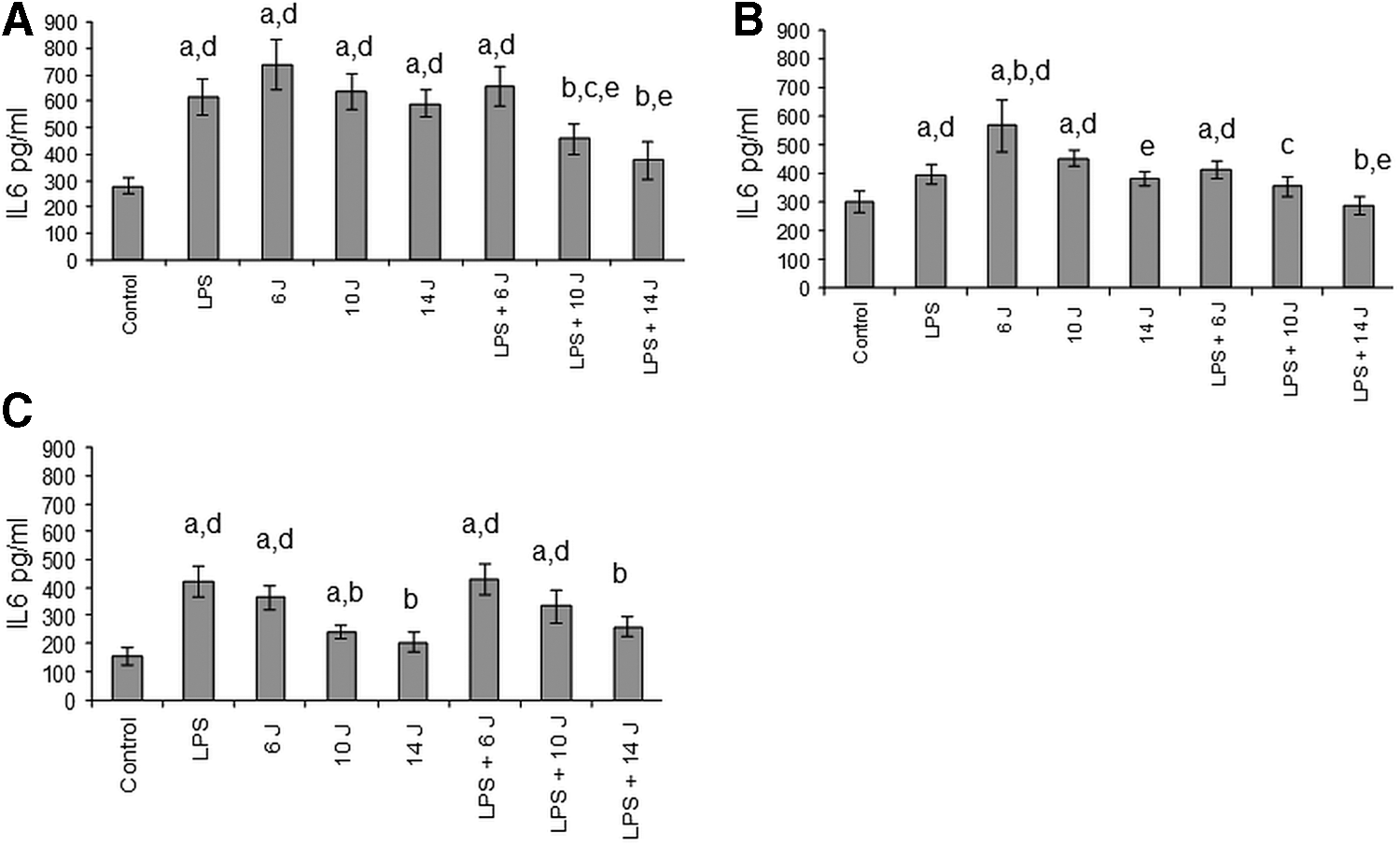
Changes in IL-6 secretion.
Effects of LPS and/or photobiomodulation on IL-8 levels
Stimulation of gingival fibroblasts with LPS or laser significantly increased IL-8 levels. Costimulation with LPS and laser largely counteracted their mutual effects such that IL-8 levels were like control levels in groups stimulated with both LPS and 14 J. Similarly, IL-8 levels were lower in the LPS +10 J stimulation group than in the only 10 J stimulation group within 24 h (Fig. 2A).

Changes in IL-8 secretion.
The second stimulation with laser induced effects similar to those observed after the initial stimulations except 14 J. Repeated stimulation with 14 J did not alter the levels of IL-8, suggesting desensitization to 14 J stimulation. Stimulation with 14 J, however, was able to counteract the effects of LPS on IL-8 release. Interestingly, repeated stimulation with 6 J increased IL-8 secretion by approximately three times, an effect more prominent than after the first stimulation (Fig. 2B). After the third stimulation, increased levels of IL-8 release were detected after LPS and LPS +6 J stimulation.
Effects of LPS and/or photobiomodulation on phosphorylated forms of p38 and ERK1/2
In accordance with the cytokine results, LPS or laser stimulation increased phosphorylated forms of p38 and ERK1/2, demonstrating the activation of intracellular pathways involving p38 and ERK1/2 compared with the untreated (control) group (Fig. 3). In contrast to the cytokine results, co-treatment with LPS and laser did neutralize their effects. In other words, p38 and ERK1/2 pathways are activated by LPS as well as by photobiomodulation. However, desensitization observed toward the effects of 14 J stimulation on cytokine release was not detected here (Fig. 3).
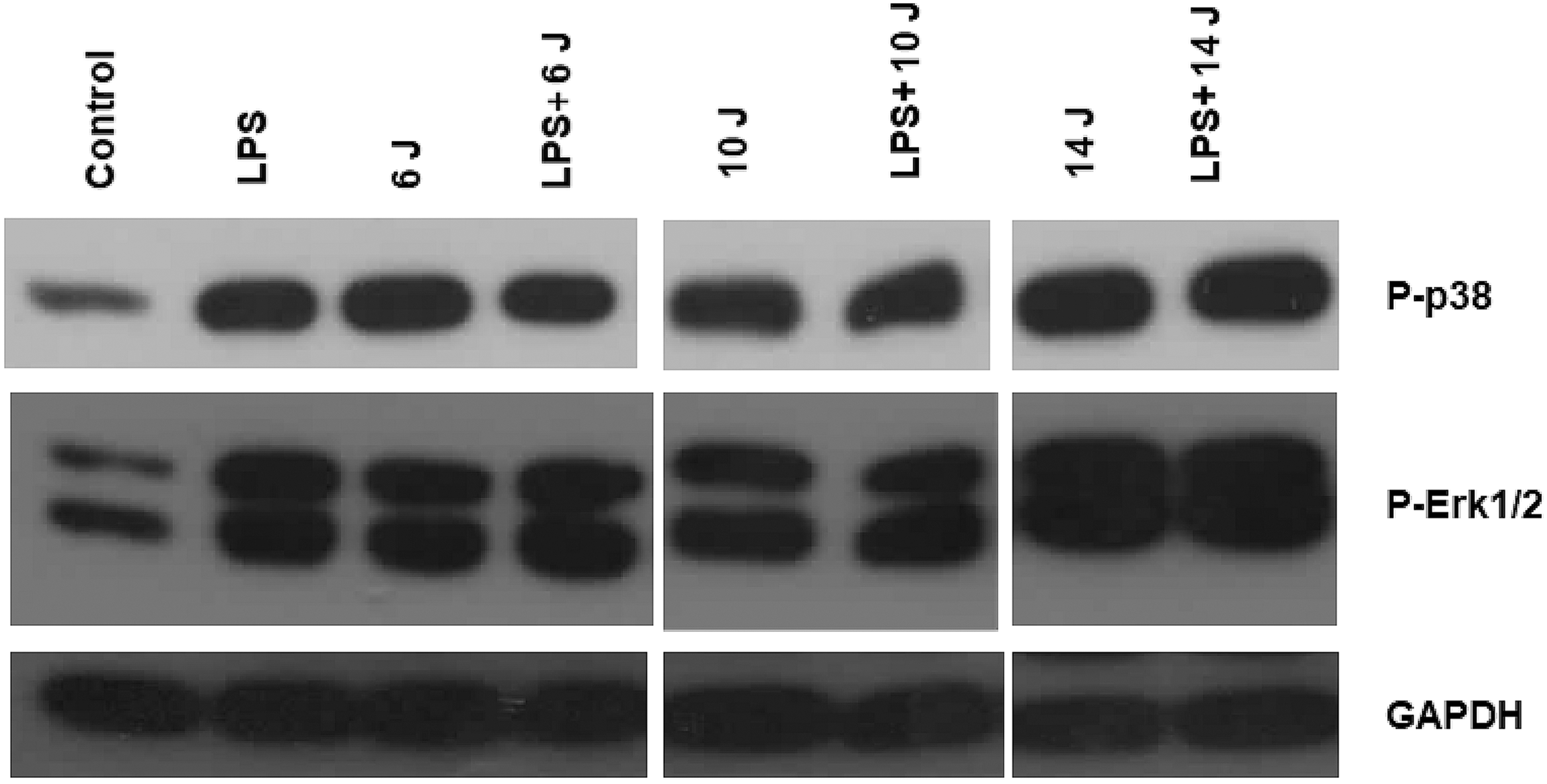
Effects of laser stimulation and LPS on p38 and ERK1/2 phosphorylation. Primary fibroblasts were lysed at the end of the experiments (after third stimulation), and phosphorylated form of p38 and ERK1/2 was determined using specific antibodies.
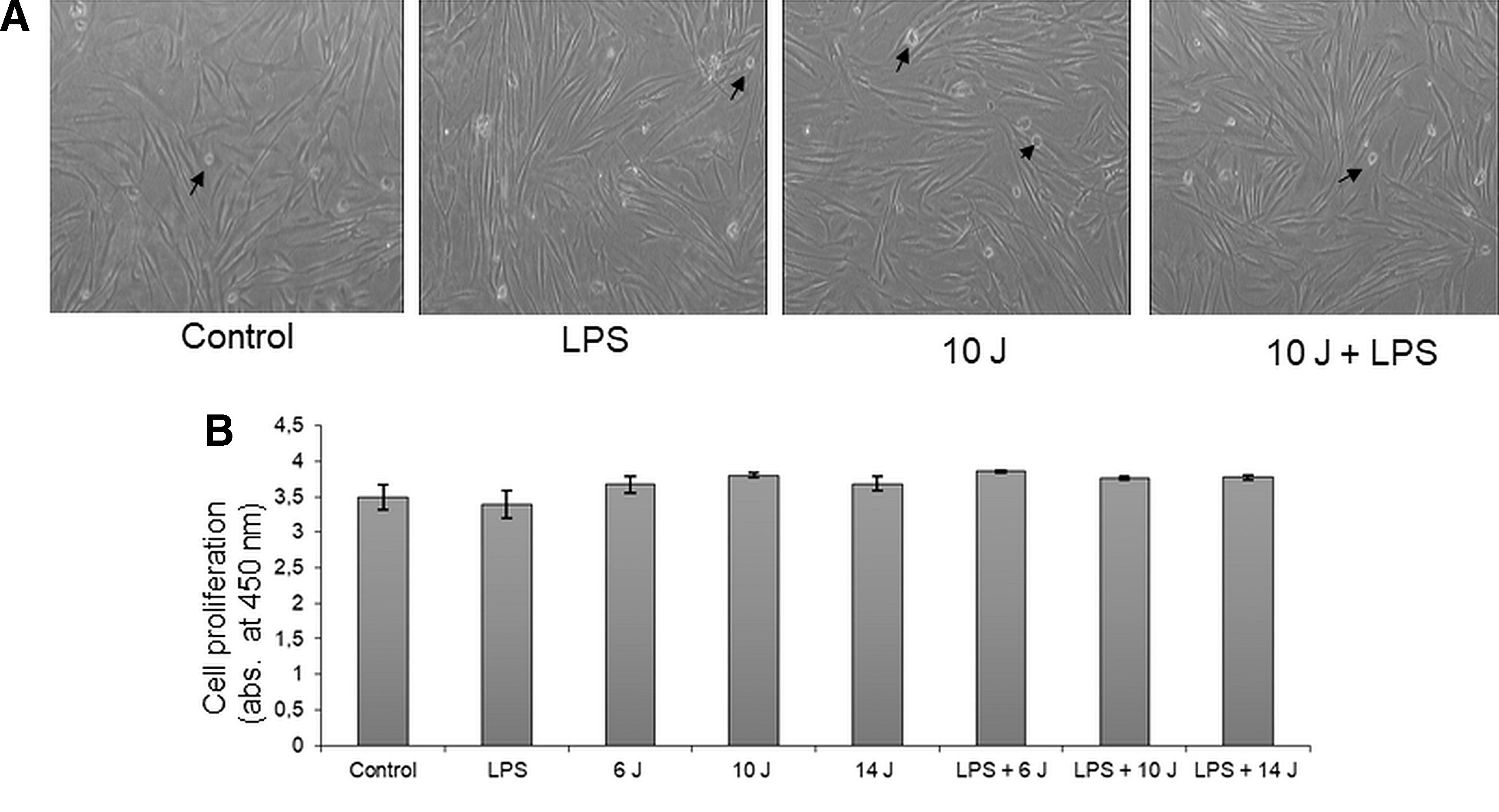
Effects of LPS and/or photobiomodulation on morphology of gingival fibroblasts
Primary fibroblasts were grown to confluence (Fig. 4A). In all groups, only a few apoptotic cells were visible (arrows), and there was no morphological variance between treatments.
Changes in cell viability.
Effects of LPS and/or photobiomodulation on cell viability
Changes in cell viability were determined by means of a WST-1 cell proliferation kit. Absorbance at 450 nm reflects cell viability. Neither LPS nor laser treatment, alone or in combination, affected the cell viability. Cell viability values were also similar in control and treatment groups, demonstrating that LPS and laser stimulation do not alter the growth rate of gingival fibroblasts.
Discussion
For pathological periodontal pockets, scaling and root planing are the traditional treatment methods. Current studies suggest that supporting these mechanical processes with laser stimulation results in successful outcomes. 13 In this study, we show that photobiomodulation, especially at relatively high doses, counteracts the release of inflammatory cytokines such as IL-6 and IL-8, as shown previously. However, in this study we show for the first time that low levels of photobiomodulation may potentiate LPS-induced inflammation. We also demonstrate that photobiomodulation and LPS activated similar intracellular pathways.
Gingival fibroblasts are not a member of immune system cells. However, they are active in defense against periodontal diseases and involved in regeneration during wound healing. 12 This study examined the effect of photobiomodulation on cellular pathways by cytokine levels, and p38 and ERK1/2 phosphorylation levels. We observed that photobiomodulation could influence IL-6 and IL-8 cytokine secretions in stimulated and nonstimulated gingival fibroblasts. We rejected the null hypothesis.
Interactions between inflammatory cells and fibroblasts regulate some local inflammatory responses. Cytokines and chemokines are dynamic messengers for the initiation and maintenance of immune responses and act as important roles in immunopathological problems. 14 As a proinflammatory cytokine, IL-6 directs inflammation responses and stimulates the production of acute phase proteins. IL-8 is also a key mediator involved in inflammation, with a longer half-life. IL-8 also acts as a chemoattractant and an angiogenic factor. 4,8,13,14 P. gingivalis is a significant periodontal pathogen found in dental plaque. 5 The host response to periodontitis begins with the effect of LPS, which interacts through the “Toll-like receptors” on the bacterial wall and initiates the inflammatory process. P. gingivalis is a Gram-negative anaerobic bacterium recognized as an important etiological agent of periodontitis. 15 P. gingivalis LPS is a crucial virulence factor strongly associated with periodontitis. Previous studies stated that the scale of the responses of gingival fibroblasts to P. gingivalis varies strongly among individuals. It is possible that heterogeneity is caused by the host genetic background of the cells. 15 –17 In our study, preliminary tests with 1 μg/mL P. gingivalis strongly stimulated the expressions of IL-6 and IL-8 without cytotoxicity. To avoid from a toxic dose, we used 1 μg/mL P. gingivalis and we applied once a time.
Laser applications significantly increased the IL-6 and IL-8 secretions like LPS stimulation after 24 h. With repetitive laser applications (after the second and third), this stimulatory effect had mostly disappeared, especially in the groups wherein the 14 J laser was applied. IL-6 and IL-8 levels were not statistically significant with those in control groups. Recurrent laser applications to immunologically stimulated gingival fibroblasts caused lower cytokine secretions. Moreover, IL-6 and IL-8 levels were comparable with those of the control groups, even after 24 h in the 14 J group. Costimulation of LPS and laser counteracted the stimulation effects. In LPS-stimulated 14 J laser-applied groups, after 48 and 72 h, IL-6 and IL-8 levels did not differ from those of control. At relatively high laser energy densities, inhibitory effects of diode lasers on selected cytokines were observed.
In this study, we tested three laser parameters (6, 10, and 14 J). Laser applications were performed with a special biostimulation tip with 7.1 cm2 area. For each square centimeter, 0.84 J–1.4 J–1.97 J energy was irradiated, respectively. In the literature, 1.5–16 J/cm2 laser irradiation is reported to alter inflammation in human gingival fibroblasts. 18 Nomura et al. 19 reported that low-energy laser irradiation (830 nm, 3.95–7.90 J/cm2) significantly inhibited IL-1 production stimulated by LPS in human gingival fibroblasts (HGF) cells. Basso et al. 20 evaluated the effects of photobiomodulation on the levels of inflammatory cytokines associated with oral mucositis. The authors demonstrated that photobiomodulation provides a beneficial biomodulatory effect on the levels of inflammatory cytokines related to oral mucositis. Photobiomodulation caused significant decreases in the levels of TNF-α, IL-6, and IL-8 at 1.5 and 3 J/cm2. In this study, however, we found that a low level of stimulation (6 J) in the presence of inflammation (LPS stimulation) may further enhance inflammatory responses.
The mitogen-activated protein kinases (MAPKs) ERK1/2, p38, and JNK are crucial components of the signaling network that coordinate a variety of cellular activities such as cell proliferation, induction of apoptosis, and the release of cytokines. Protein kinases are mainly activated by environmental stresses including oxidative stress, but also by the bacterial LPS. 21 –23 MAPKs act as focal points for some extracellular stimuli and regulate a variety of cell functions. 22
In this study, LPS or laser stimulation increased the phosphorylated forms of p38 and ERK1/2 compared with those in the control group and activated intracellular pathways containing p38 and ERK1/2. However, unlike cytokine secretions, the combination of LPS stimulation with laser treatment did not reduce the mutual effects of p38 and ERK1/2 phosphorylation. Similarly, observed desensitization in cytokine release after 14 J laser application was not detected in p38 and ERK1/2 phosphorylation levels. These results indicate that in stimulated cells, cytokine secretion inhibition because of laser use and the desensitizing effect of the 14 J laser application in nontreated cells do not originate from the same intracellular pathways.
Biological responses of cells to photobiomodulation occur through chemical and physical changes in the components of respiratory chains and light-receiving molecules. 24 Principally, the light activates receptors on the cell membrane or mitochondria and translates this energy into chemical energy. It is well known that photobiomodulation has a proliferative effect on fibroblast cells. 18 Illescas-Montes et al. 12 reported that a 940-nm laser has a proliferative effect on HGF cells, depending on dose: 1–4 J/cm2 of laser treatment increased proliferative activity, and this effect was lost >0.5 W. In this study, the WST-1 cell viability test demonstrated no significant change in cell populations after either laser or LPS treatments (p >0.05). Hakkı and Bozkurt 25 have found a similar result. They stated that diode laser parameters they tested (0.3, 1.5, and 2 W) were not efficient in cell proliferation. In this study, cell viability was tested only at 72 h. We should state that different time point results may vary. Also, it is critical to state that the WST-1 cell proliferation test used in this study is based on the cleavage of tetrazolium salts by mitochondria while photobiomodulation also affects mitochondrial metabolism. This might also be due to high confluence of cells before treatment that masks the LPS-induced stress and the possible protective effects of photobiomodulation. One should interpret the results with these shortcomings.
Diode laser applications could yield desirable results since they manage to penetrate deeper into periodontal tissues and facilitate periodontal inflammation control and tissue repair. 26 Karu 24 reported that the effects of lasers on cell cultures are dependent on the wavelength, dose, severity, and cell culture conditions of the blend. The optimal dosage or therapeutic window for photobiomodulation is a topic for discussion in the literature, and various factors such as power density, application method, and exposure interval may affect the efficiency of treatment. Also, dosages could change the overall outcomes. 14
For clinical applications, the findings of this study do not accurately reflect the in vivo situation, because wound healing is a dynamic process, and various cells, cytokines, and mediators would join this progression. The photobiomodulatory response to cells in a tissue or culture may not be identical to that of another cell type, and different cells may be affected differently in similar parameters. Also, periodontal tissues can absorb considerable quantity of laser energy in vivo. In contrast, diode lasers are economical and very convenient. As shown in this study, they have significant potential to be used as an alternative to oral analgesics and nonsteroidal anti-inflammatory drugs, with limited side effects. 26 Further studies are required with different promising low-intensity lasers (for instance, 660 nm) 27 to provide the most efficient protocols.
Conclusions
This in vitro study showed that in LPS-stimulated gingival fibroblasts, photobiomodulation can inhibit IL-6 and IL-8 secretions while not reducing the mutual effects of p38 and ERK1/2 phosphorylation. The inhibitory effects on cytokine secretions were dose dependent, and the time frame of irradiation influenced the results. Low levels of photobiomodulation, in contrast, may potentiate LPS-induced inflammation, which might be beneficial for the bactericidal effects of acute inflammation but may also lead to increased cytokine secretion. All these results depend on doses, and future clinical studies are required.
Footnotes
Acknowledgments
This study was supported by The Scientific Research Projects Coordination Unit of Akdeniz University, Project Number 2014.01.0151.003.
Ethical Approval
This study was approved by the Research Ethics Committee of Akdeniz University (protocol no: 214/2017). All procedures performed were in accordance with the Declaration of Helsinki.
Author Disclosure Statement
No competing financial interests exist. The authors declare that they have no conflicts of interest.