
Abstract
Current immunosuppressive treatments for central nervous system demyelinating diseases fail to prevent long-term motor and cognitive decline in patients. Excitingly, glial cell transplantation arises as a promising complementary strategy to challenge oligodendrocytes loss occurring in myelination disorders. A potential source of new oligodendrocytes is the subventricular zone (SVZ) pool of multipotent neural stem cells. However, this approach has been handicapped by the lack of functional methods for identification and pharmacological analysis of differentiating oligodendrocytes, prior to transplantation. In this study, we questioned whether SVZ-derived oligodendrocytes could be functionally discriminated due to intracellular calcium level ([Ca2+]i) variations following KCl, histamine, and thrombin stimulations. Previously, we have shown that SVZ-derived neurons and immature cells can be discriminated on the basis of their selective [Ca2+]i rise upon KCl and histamine stimulation, respectively. Herein, we demonstrate that O4+ and proteolipid protein-positive (PLP+) oligodendrocytes do not respond to these stimuli, but display a robust [Ca2+]i rise following thrombin stimulation, whereas other cell types are thrombin-insensitive. Thrombin-induced Ca2+ increase in oligodendrocytes is mediated by protease-activated receptor-1 (PAR-1) activation and downstream signaling through Gq/11 and phospholipase C (PLC), resulting in Ca2+ recruitment from intracellular compartments. This method allows the analysis of functional properties of oligodendrocytes in living SVZ cultures, which is of major interest for the development of effective grafting strategies in the demyelinated brain. Additionally, it opens new perspectives for the search of new pro-oligodendrogenic factors to be used prior grafting.
Introduction
Recent advances in neural stem cell (NSC) biology have raised considerable prospects for the development of remyelinating therapies based on glial cell replenishment. The multipotency of NSCs has been well characterized, unveiling a renewable source of new oligodendrocytes for use in regenerative medicine concerning myelin pathologies. 2,3 Indeed, subventricular zone (SVZ) stem cells produce oligodendrocyte progenitors that differentiate into mature and functional myelinating oligodendrocytes. 3 –7 Nevertheless, remaining hurdles still limit the grafting success, including a lack of functional pretransplantation studies on reparative cells that identify key pharmacological targets to drive their differentiation in myelinating oligodendrocytes, as well as their survival and integration in the diseased brain. Currently, characterization of oligodendrocytes or their progenitors is performed mainly by immunocytochemical studies, limiting the support of an engrafting approach.
In the present study, we describe a functional method to study oligodendrocytes derived from SVZ cultures. Thus, monitoring the intracellular calcium variations ([Ca2+]i) of single cells upon a defined sequence of stimulations allows the identification of oligodendrocytes among other cells in SVZ cultures and their pharmacological characterization. Our group has previously developed a calcium imaging protocol that rapidly discriminates neurons and immature cells in SVZ cultures. 8 Indeed, KCl-induced depolarization and a consequent rise in [Ca2+]i is typical of excitable neuronal cell lineage, whereas histamine triggers a response in immature cells. This method unveiled the proneurogenic effects of neuropeptide Y (NPY) and tumor necrosis factor-α (TNF-α) in mice SVZ cultures. 9,10 We hypothesized whether thrombin can be used as a specific stimulus for SVZ-derived oligodendrocytes. Wang et al. 11 have shown that the rat OLN-93 oligodendrocyte cell line displays an increase of [Ca2+]i following thrombin stimulation, an effect mediated by protease-activated receptor-1 (PAR-1) activation. Thus, we propose to study the thrombin-induced calcium transients in SVZ-derived oligodendrocytes and develop a reliable protocol based on single-cell calcium imaging to identify new SVZ-derived oligodendrocytes. This will be a useful tool to perform pharmacological studies and cell fate identification in SVZ NSC cultures. Our results show that oligodendrocytes in SVZ cultures can, indeed, be distinguished according to the uniqueness of their response to thrombin.
Materials and Methods
All experiments were performed in accordance with National Institutes of Health (NIH) and European Union (86/609/EEC) guidelines for the care and use of laboratory animals.
Cell cultures
SVZ cells were obtained from P1-3 C57Bl/6 mice as described previously in Agasse et al.
8
Six- to 8-day-old primary neurospheres were deposited on 0.1 mg/mL poly-
Cortical cells were obtained from the dissection of the parietal cortex of early postnatal mice. The cells were cultured similarly as for SVZ cells. Seven- to 8-day primary cultures of hippocampal neurons were prepared from E18-19 C57Bl/6 mice embryos, as described in Silva et al. 12 . Seven-day cortical astrocyte cultures were prepared from P6-7 C57Bl/6 mice as described in Matos et al. 13
Single cell calcium imaging analysis
Following the 7- to 10-day differentiation period, SVZ cultures were loaded for 40 min, at 37°C, with 5 μM Fura-2AM (Molecular Probes, Eugene, OR) in Krebs solution (132 mM NaCl, 4 mM KCl, 1.4 mM MgCl2, 2.5 mM CaCl2, 6 mM glucose, 10 mM HEPES, pH 7.4) plus 0.1% fatty acid free bovine serum albumin (BSA; Sigma-Aldrich) and 0.02% pluronic acid F-127 (Molecular Probes). Afterward, cells were washed in Krebs and subsequently mounted on RC-25 chamber (Warner Instruments, Hamden, CT,
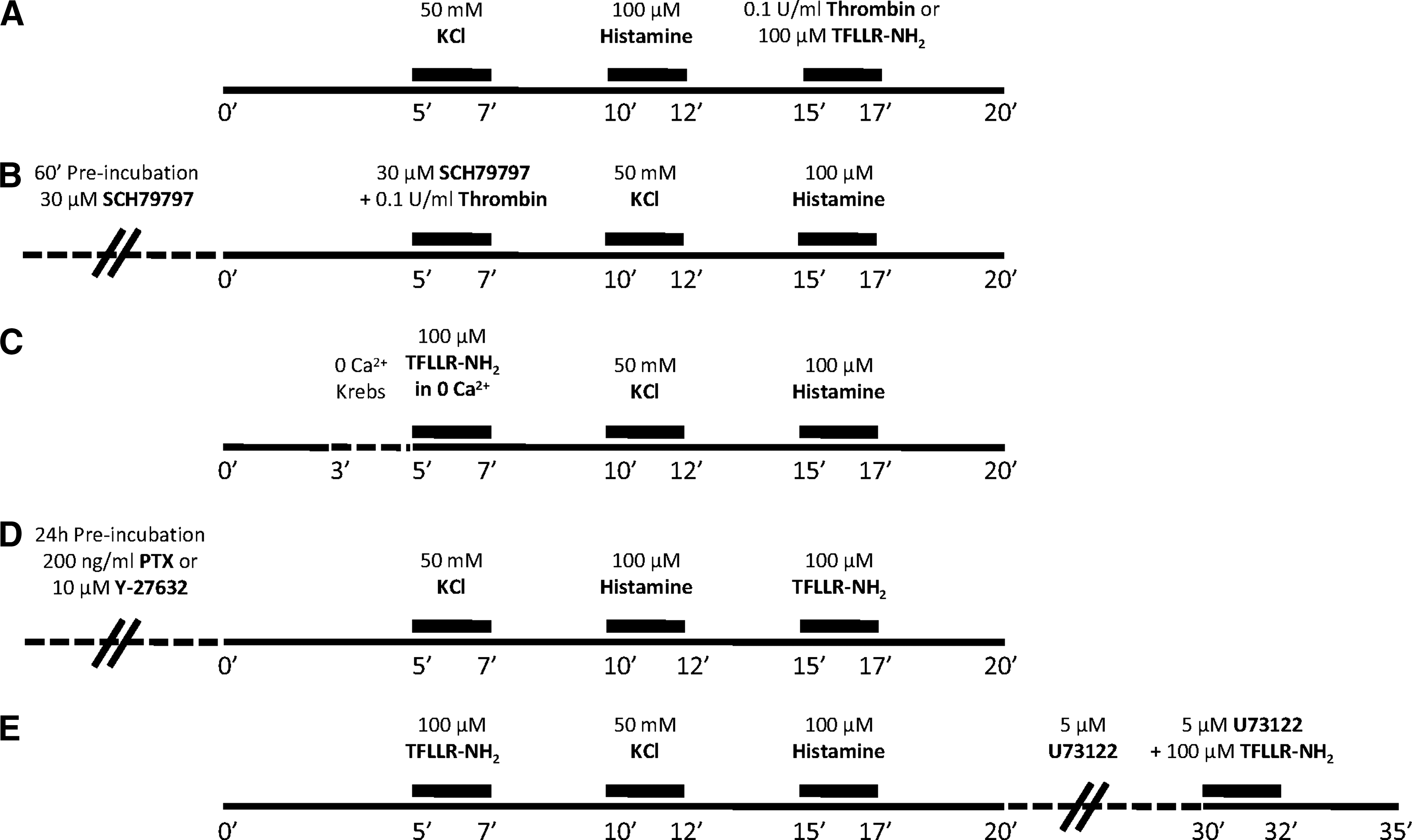
Experimental protocols performed in single cell calcium imaging (SCCI) analysis, aiming at the functional identification of subventricular zone (SVZ)–derived oligodendrocytes by thrombin response (
With this data we could quantify the percentage of astrocytes plus oligodendrocytes, or oligodendrocytes only, respectively, in SVZ cultures. Indeed, as demonstrated in Agasse et al. 2008 8 , a Hist/KCl ratio below 0.8 identifies neurons, whereas a Hist/KCl ratio above 1.1 identifies immature cells. Glial cells display a Hist/KCl ratio between 0.8 to 1.1 as confirmed by using primary astrocyte cultures. According to our hypothesis, Throm/Hist ratio would allow us to discriminate oligodendrocytes from other cells present in SVZ-derived heterogeneous cell populations. Additionally, the same single cell calcium imaging (SCCI) experimental protocol (Fig. 1A) was carried out on hippocampal neurons and cortical astrocytes.
To identify the signaling transduction pathway involved in the thrombin-induced [Ca2+]i rise in SVZ-derived oligodendrocytes, we designed other protocols of stimulation, represented in Fig.1, A–E. To investigate protease-activated receptors (PARs) involved in the response, 14 –16 a PAR-1 agonist TFLLR-NH2 (100 μM; Tocris Cookson, Bristol, UK) and a selective nonpeptide PAR-1 antagonist, SCH79797 (30 μM), 17 were used (Fig.1A,B).
To assess whether the elevation of [Ca2+]i induced by PAR-1 results from Ca2+ release from intracellular stores or extracellular Ca2+ influx, we replaced the regular Krebs solution by a Krebs solution containing low Ca2+ concentration (0 Ca2+ Krebs: regular Krebs with 38 μM instead of 2.5 mM CaCl2 and 50 μM EGTA) 2 min before stimulation with TFLLR-NH2, prepared in 0 Ca2+ Krebs as well (Fig. 1C). To further explore the signaling mechanism triggered by PAR-1 activation, particularly the coupling to heterotrimeric G proteins, a set of SVZ cells was preincubated with 200 ng/mL pertussis toxin (PTX; Sigma-Aldrich), which inhibits Gi/o proteins, or with 10 μM Y-27632 (Calbiochem, Merck Biosciences, Darmstadt, Germany), which inhibits the G12/13 effectors Rho/ROCK, 24 h before SCCI experiments, where cells were stimulated with TFLLR-NH2 (Fig. 1D). To investigate whether the rise in [Ca2+]i resulted from phospholipase C (PLC) activation, cells were first stimulated with TFLLR-NH2, KCl, and histamine, and then incubated for 10 min with 5 μM U73122 (Sigma-Aldrich), a PLC inhibitor, before a last pulse of TFLLR-NH2 plus U73122 (Fig. 1E).
Immunocytochemistry
Cells were fixed in 4% paraformaldehyde (PFA; Sigma-Aldrich) diluted in phosphate-buffered saline (PBS), and classical immunocytochemistry was performed using the following primary antibodies: mouse monoclonal anti-O4 (1:100; Chemicon, Millipore), rabbit polyclonal anti-NG2 (1:500; Chemicon, Millipore), mouse monoclonal anti-proteolipid protein (PLP) (1:600; Serotec, Oxford, UK), mouse monoclonal anti-MAP-2 (1:200; Sigma-Aldrich), rabbit monoclonal anti-glial fibrillary acidic protein (GFAP; 1:100; Sigma-Aldrich), mouse monoclonal anti-nestin (1:200; Chemicon, Millipore), rabbit polyclonal anti-doublecortin (1:200; Cell Signaling Technology, Danvers, MA), goat polyclonal PAR-1 (1:200; Santa Cruz Biotechnology, Santa Cruz, CA). The following secondary antibodies were used: donkey anti-rabbit Alexa Fluor 488 (1:200), rabbit anti-mouse Alexa Fluor 488 (1:200), rabbit anti-goat Alexa Fluor 594 (1:200), and goat anti-mouse Alexa Fluor 594 (1:200; all from Invitrogen, Carlsbad, CA). Nuclei were counterstained with Hoechst 33342 (2 μg/mL, Invitrogen), and coverslips were mounted in Dako fluorescent medium (Dako, Glostrup, Denmark). Fluorescent images were taken in an Axioskop microscope (Carl Zeiss) or with a confocal microscope (LSM 510 Meta; Carl Zeiss, Oberkochen, Germany).
Data analysis
In SCCI experiments, fluorescence measurements were performed in cells located at the outside border of the seeded neurospheres. Upon deposition of the spheres, these cells migrated from the periphery of the sphere forming a pseudo-monolayer where single cells are distinguishable and easily delimited, allowing single cell analysis. Each experimental condition was assessed at least in three different wells (about 100 cells per coverslip). The experiments were replicated at least in three different cultures except where otherwise specified. Data are expressed as means ± standard error of the mean (SEM). Statistical significance was determined by using one-way analysis of variance (ANOVA), followed by Bonferroni posttest for multiple comparisons, with *p < 0.05, **p < 0.01, ***p < 0.001.
Results
Thrombin triggers the increase in [Ca2+]i in SVZ-derived oligodendrocytes
We have previously demonstrated that neurons and immature cells in SVZ cultures can be rapidly distinguished on the basis of their unique responses to KCl or histamine, respectively. 8 Accordingly, the membrane depolarization evoked by KCl on neurons results in Ca2+ entry via voltage-sensitive Ca2+ channels, whereas histamine activates the H1 receptor expressed in immature SVZ cells, inducing the mobilization of Ca2+ from the intracellular stores. Moreover, neither KCl nor histamine trigger a significant [Ca2+]i rise in astrocytes. Herein, we characterize the intracellular Ca2+ profile of oligodendrocytes upon perfusion with KCl and histamine and used thrombin as a putative [Ca2+]i rise inducer in SVZ-derived oligodendrocytes. 11
For this purpose, primary mouse SVZ neurospheres were seeded on poly-
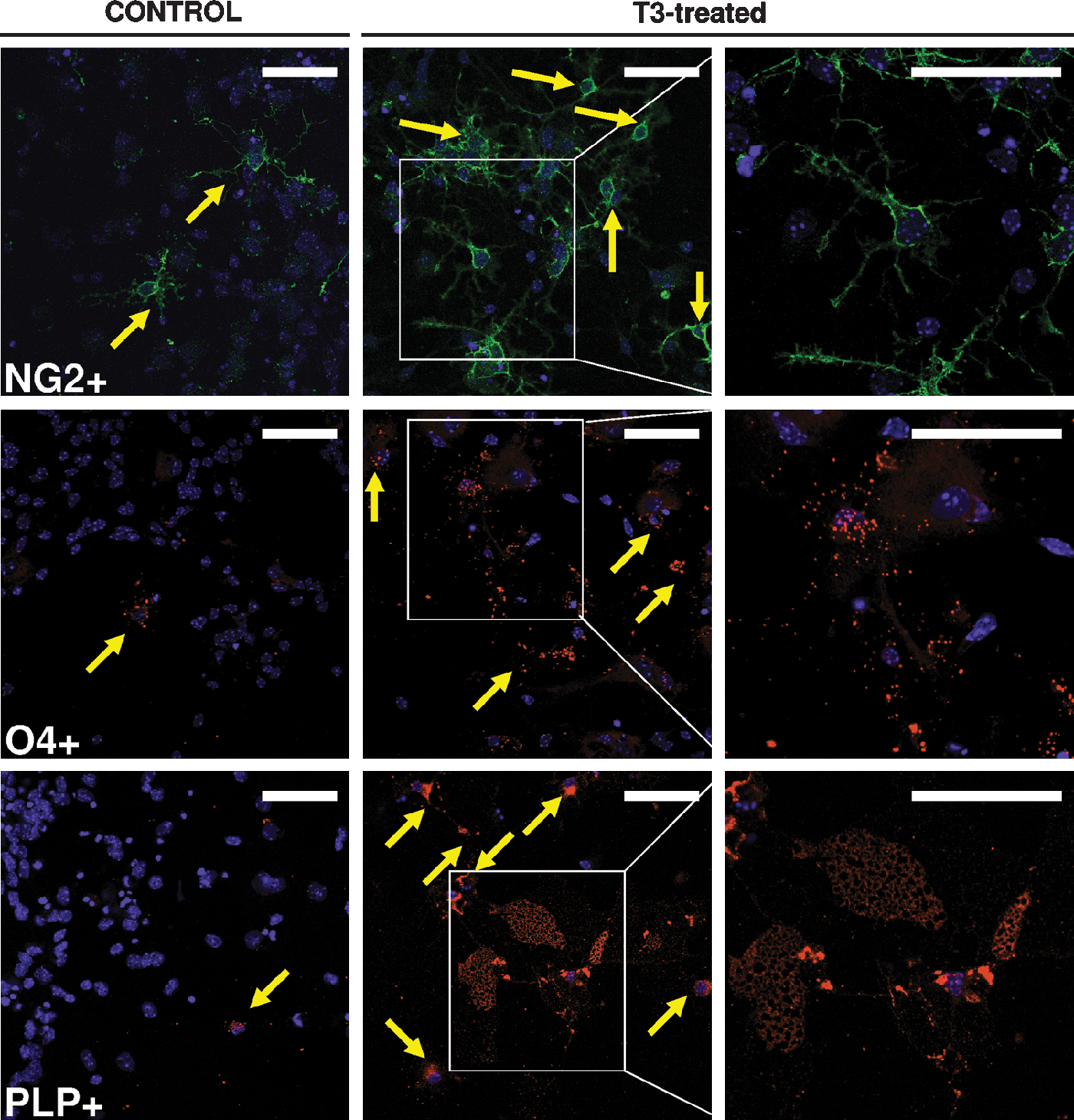
T3 increases the production of oligodendrocytes in subventricular zone (SVZ) cultures. Representative fluorescence images of NG2+ cells (green; top), O4+ oligodendrocytes (red; middle), and PLP+ oligodendrocytes (red; bottom), counterstained with Hoechst 33342 (blue nuclei) in SVZ cells treated or not with 30 nM T3 for 7–10 days after spheres deposition. Yellow arrows indicate NG2+, O4+, and proteolipid protein-positive (PLP+) cells in control and 3,3′-5-triiodo-
To correlate profiles of [Ca2+]i oscillations with the respective cell phenotype, we performed SCCI analysis in SVZ neurospheres seeded on microgrid coverslips and subsequent immunostaining for the different cell type markers. Cells were preloaded with Fura-2AM Ca2+ probe and then continuously perfused with Krebs solution. They were stimulated subsequently with 2-min pulses of 50 mM KCl, 100 μM histamine, and 0.1 U/mL thrombin (Fig. 1A). Single cell [Ca2+]i oscillations were given by the normalized ratios of Fura-2AM fluorescence at 340/380. We demonstrated that O4+ oligodendrocytes do not respond to KCl or histamine, whereas thrombin clearly induces an increase in [Ca2+]i in these cells (Fig. 3A). O4 sulfatide expression begins at the stage of preoligodendrocytes and remains throughout maturation into mature myelinating oligodendrocytes, when myelin proteins are expressed (for review see Baumann et al. 2001 21 ). To further detail the response of oligodendrocytes to thrombin in different stages of maturation, profiles of [Ca2+]i variations were investigated in NG2+ and PLP+ cells. NG2 is a proteoglycan expressed in oligodendrocyte progenitors whereas the myelin proteolipid protein PLP is the major constituent of CNS myelin, being expressed by mature oligodendrocytes. PLP+ oligodendrocytes respond similarly to the stimuli as do the O4+ cells (Fig. 3B). Concerning NG2+ cells, we observed a significant variability in the profile of response (data not shown). This included cells that responded exclusively to thrombin, but also cells that responded to histamine alone, or to both thrombin and histamine. This variability in the response may be correlated with the variability of progenitors that express NG2. Indeed, NG2+ cells from the early postnatal SVZ are C-type cells, i.e., multipotent progenitors able to give rise to interneurons. 22 Most of the NG2+ cells responded to histamine at immature stages. It is noteworthy that no NG2+ cell responding to both KCl and thrombin was observed (data not shown).
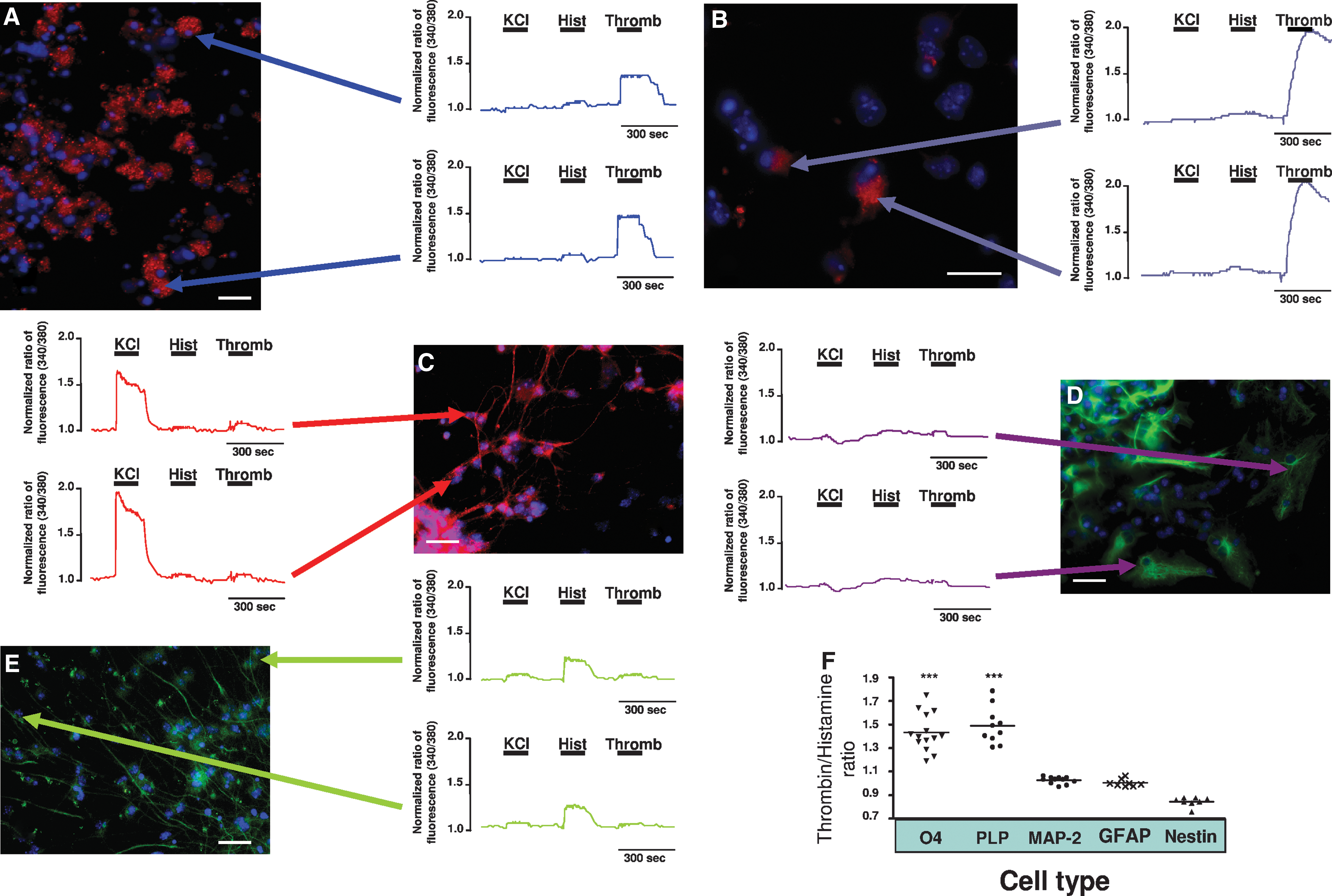
[Ca2+]i variations upon KCl, histamine (Hist), and thrombin (Thromb) in subventricular zone (SVZ)–derived cells. O4+ oligodendrocytes (
Increase of [Ca2+]i upon thrombin stimulation was not seen in MAP-2+ (microtubule-associated protein 2–positive) neurons (Fig. 3C), or in DCX+ neuroblasts (data not shown), Nestin+ immature cells (Fig. 3D), or GFAP+ astrocytes (Fig. 3E). In agreement with the previous work, cells from the neuronal lineage presented a [Ca2+]i rise upon KCl stimulation, and none [MAP-2+] neurons or a weak (DCX+ neuroblasts) response to histamine, whereas immature cells expressing Nestin responded solely to the histamine pulse.
SVZ-derived oligodendrocytes can be identified on the basis of the selective response to thrombin
Normalized data were used to calculate the ratio of responses to thrombin and histamine (Throm/Hist) (Fig. 3E). The high Throm/Hist values for O4+ and PLP+ oligodendrocytes (O4, 1.43 ± 0.04, 14 cells analyzed; PLP, 1.49 ± 0.05, 10 cells analyzed) were due to their marked response to thrombin and no response to histamine, whereas the low values of Throm/Hist for Nestin+ cells (0.84 ± 0.01, 8 cells analyzed) reflect their null response to thrombin but high response to histamine. Other cell types had a Throm/Hist ratio of approximately 1 because they did not respond to any of these two stimuli (11 MAP-2+ neurons [1.03 ± 0.01], 10 DCX+ neuroblasts [1.02 ± 0.01], and 8 GFAP+ astrocytes [1.00 ± 0.01] were analyzed). According to these findings, SVZ-derived oligodendrocytes can be functionally discriminated among other cells in SVZ cultures. We concluded that, in our cultures, cells with a Throm/Hist ratio above 1.3 are oligodendrocytes.
Thrombin-induced [Ca2+]i increase in SVZ-derived oligodendrocytes is mediated by PAR-1 activation
Many of the effects of thrombin are mediated by PAR-1, being the most extensively studied subtype and the prototype of the PARs family. 14,23 PAR-4 is a low-affinity receptor requiring higher concentrations of thrombin for activation in comparison with the other thrombin receptors. 15 PAR-3 does not signal by itself having a possible co-factor role in the cleavage and activation of PAR-4. 16 On the basis of this knowledge, we focused on PAR-1 expression and function in SVZ cultures. Our hypothesis was also supported by the finding that PAR-1 is the only thrombin receptor functionally expressed in OLN-93, where it was associated with an increase of [Ca2+]i evoked by thrombin. 11 In addition, the same authors showed that primary cultured rat oligodendrocytes express high levels of PAR-1 but almost undetectable levels of PAR-3 and no PAR-4.
Therefore, we ran a perfusion protocol consisting on the subsequent application of KCl, histamine, and TFLLR-NH2, a PAR-1 agonist (Fig. 1A), in T3-treated SVZ cultures. We focused on the glial cell population, i.e., responding with a Hist/KCl ratio between 0.9 and 1.1. Cells were highly responsive to TFLLR-NH2 (Fig. 4A, top), showing a similar profile to the one obtained upon thrombin stimulation (see Fig. 6B). Additionally we used SCH79797, a selective PAR-1 antagonist. After a 60-min preincubation of T3-treated SVZ cultures with SCH79797 (Fig. 1B), we verified that thrombin-induced rise of [Ca2+]i was abolished (Fig. 4, bottom). We measured the peak of response due to thrombin, or TFLLR-NH2, or thrombin plus SCH79797. A similar magnitude of response to TFLLR-NH2 (1.42 ± 0.01, 854 cells analyzed) as compared to thrombin (1.41 ± 0.01, 431 cells analyzed) was observed, as well as a complete inhibition of thrombin peak by SCH79797 (1.06 ± 0.01, 381 cells analyzed) suggesting that PAR-1 is the receptor involved in the thrombin-induced [Ca2+]i increase in SVZ-derived oligodendrocytes (Fig. 4B). In agreement, we observed a dense and clustered PAR-1 punctuated immunoreactivity in every O4+ and PLP+ cell, whereas MAP-2+ neurons do not express PAR-1. Nevertheless some GFAP+ and Nestin+ cells express PAR-1 (Fig. 4C), albeit not responding to thrombin stimulus.
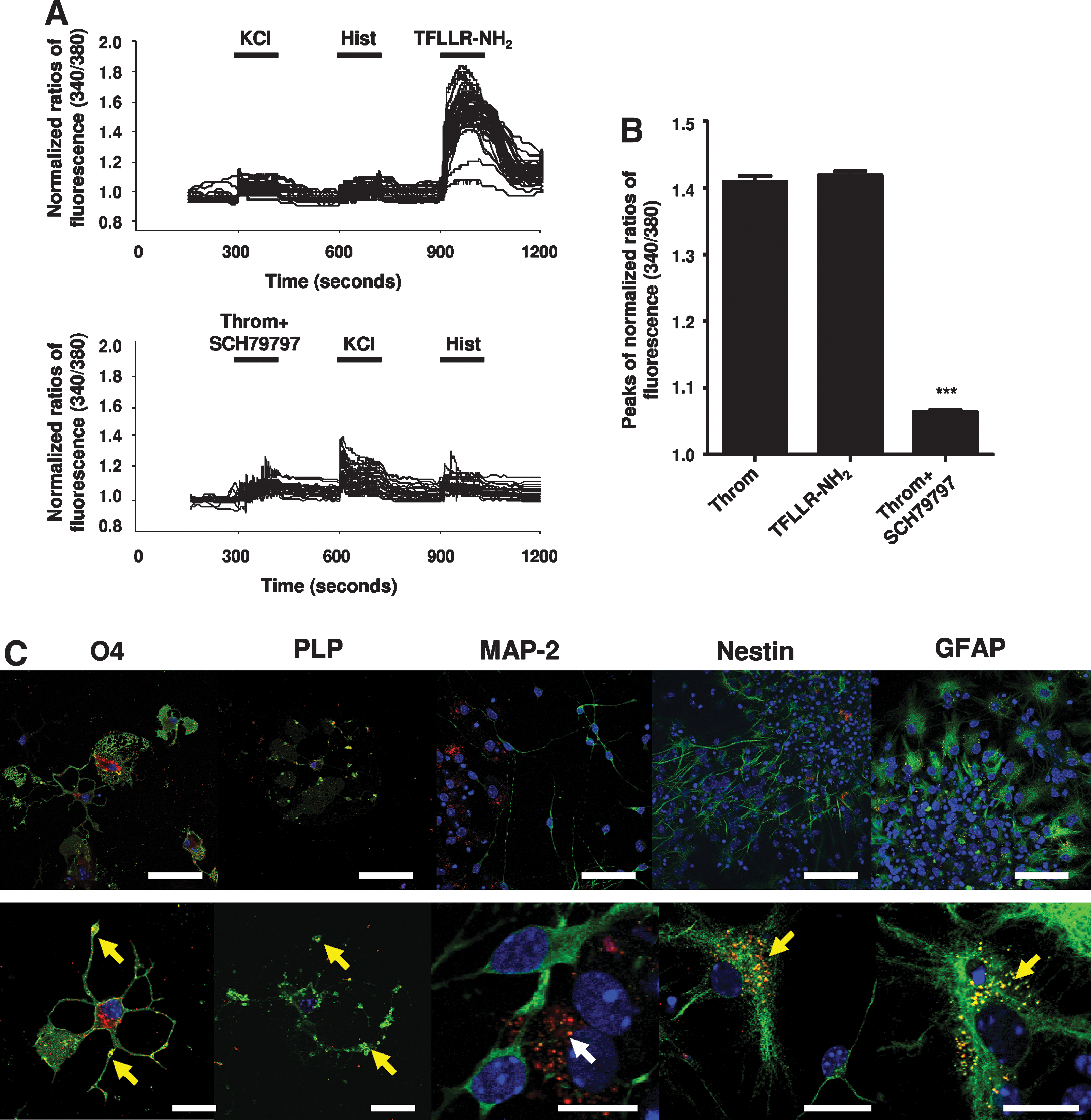
Thrombin-induced [Ca2+]i increase in subventricular zone– (SVZ) derived oligodendrocytes is mediated by protease-activated receptor-1 (PAR-1) activation. (
Moreover, the signaling mechanism of PAR-1-induced [Ca2+]i rise in SVZ cultures was investigated. As Fig. 1C shows, we replaced the normal Krebs by a 0 Ca2+ Krebs solution 2 min before TFLLR-NH2 stimulation and during this pulse. Under these conditions, the increase of [Ca2+]i was as high (TFLLR-NH2 in 0 Ca2+, 1.38 ± 0.01; 127 cells analyzed) as in normal Krebs, indicating that Ca2+ comes from intracellular stores and not from the extracellular solution (Fig. 5).
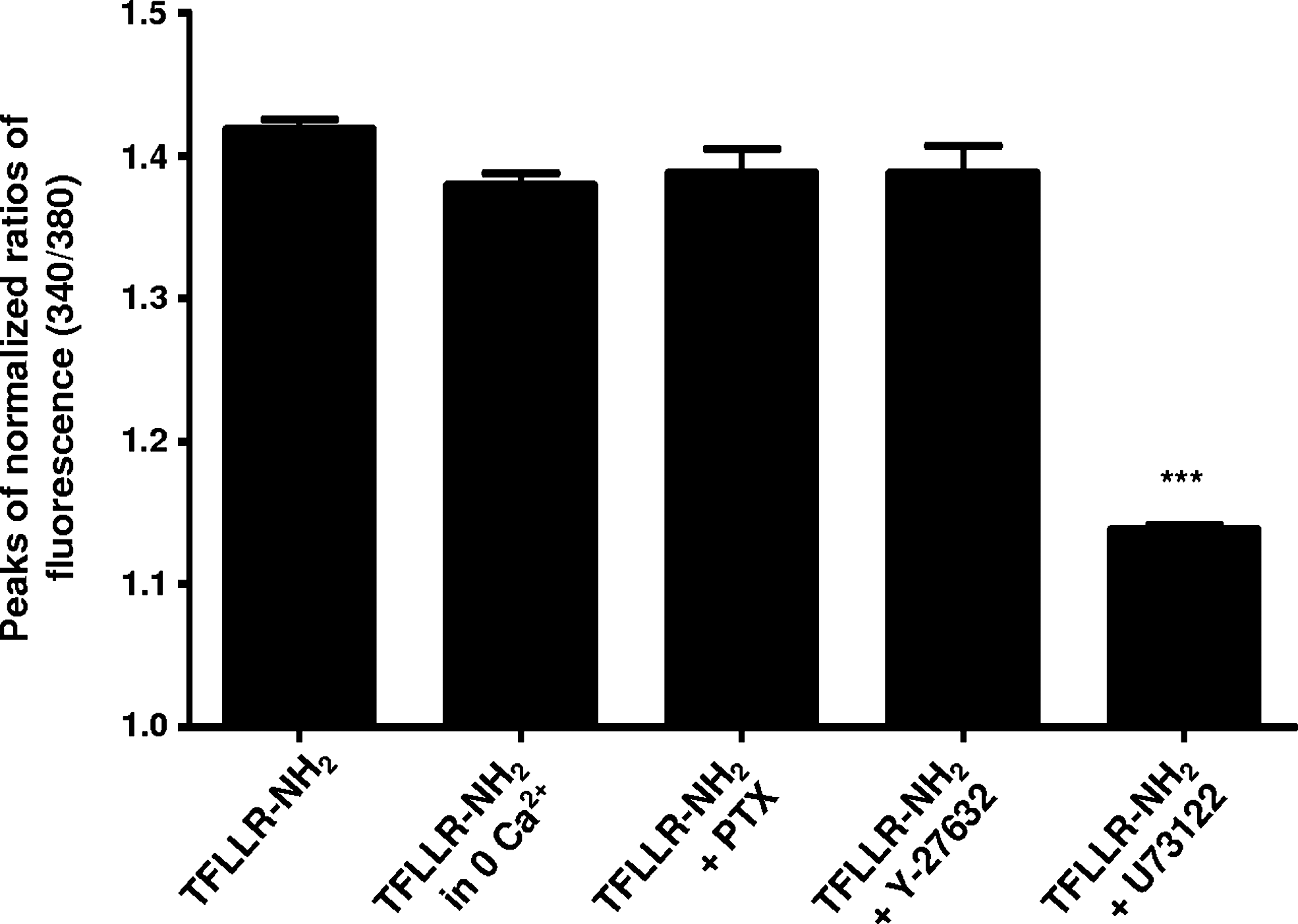
PAR-1 signaling in subventricular zone– (SVZ)-derived oligodendrocytes involves Gq/11 and phospholipase C (PLC) activity, with calcium recruitment from intracellular stores. Means of peaks of normalized ratios of fluorescence at 340/380 displayed by SVZ-derived oligodendrocytes following perfusion protocols, as in Fig.1A (TFLLR-NH2 as third pulse) and C–E. Data are expressed as means ± standard error of the mean (SEM) of at least four independent experiments. (***) p < 0.001 using one-way ANOVA and Bonferroni post-test, as compared to TFLLR-NH2.
PAR-1 is a transmembrane metabotropic G protein–coupled receptor coupled to members of the Gi/o, G12/13, and Gq/11 families of large heterotrimeric G proteins. 23 To disclose the role of G proteins in the thrombin-induced [Ca2+]i rise, we preincubated cells with 200 ng/mL PTX, which inactivates Gi/o proteins 11 (Fig. 1D). PTX incubation did not alter the Ca2+ mobilization in SVZ-derived oligodendrocytes, indicating that Gi/o are not involved in the PAR-1–induced [Ca2+]i increase (1.39 ± 0.02, 149 cells analyzed) (Fig. 5). Therefore, the signaling cascade triggered by TFLLR-NH2 includes the PTX-insensitive proteins Gq/11 and/or G12/13. The pathway upon PAR-1-coupled Gq/11 activation is quite well-described. Gq/11 activates phospholipase C isoform β (PLCβ), which catalyzes phosphoinositide hydrolysis, leading to Ca2+ recruitment from the intracellular compartments, and protein kinase C (PKC) activation. On the other hand, G12/13 binds guanine-nucleotide exchange factors (RhoGEFs) initiating a Rho-associated protein kinase– (ROCK) dependent pathway that leads to the activation of PLC isoform ε (PLCε), thus resulting in Ca2+ recruitment and PKC activation. 24 –26 To search for the involvement of Gq/11 and/or G12/13 in PAR-1–induced [Ca2+]i oscillations in SVZ-derived oligodendrocytes, we preincubated cells with 10 μM Y-27632, a ROCK inhibitor, for 24 h, and then ran the SCCI protocol, as depicted in Fig. 1D. When ROCK was inhibited, the TFLLR-NH2-induced [Ca2+]i increase was still present and at the same magnitude as compared to the peak evoked by TFLLR-NH2 alone (1.39 ± 0.02, 284 cells analyzed). Furthermore, preincubation with 5 μM U73122, a PLC inhibitor (Fig. 1E), abolished the response to TFLLR-NH2, indicating that PLC activation is part of the triggered pathway (1.14 ± 0.01, 398 cells analyzed).
Oligodendroglial differentiation can be assessed by measuring the variations of [Ca2+]i upon stimulation with thrombin and histamine
To validate the method described in the present work, we functionally compared nontreated SVZ cultures with T3-treated cultures. Upon SCCI assay consisting of KCl, histamine, and thrombin pulses (Fig. 1A), normalized peaks of fluorescence of all the individualized cells were measured and the Throm/Hist ratio was calculated. Indeed, control cultures presented ∼3% of cells responding with a Throm/Hist ratio above 1.3 (3.24 ± 1.06, 21 coverslips, 1790 cells analyzed) consistent with the normal oligodendrocyte differentiation in SVZ cultures. 27,28 Upon T3 treatment, cultures contained ∼35% of these cells (35.07 ± 8.22, 10 coverslips, 626 cells analyzed) (Fig. 6A), which is in accordance with the well-described effect of T3 hormone increasing the OPCs number and promoting the differentiation in oligodendrocytes. 29 –32 Furthermore, SVZ cultures pretreated with the astrogliogenic factor CNTF 27,33 or the neurogenic factor SCF, 34 during the differentiation period, displayed a similar percentage of cells responding as oligodendrocytes as found in nontreated cultures (CNTF, 2.92 ± 1.19, 8 coverslips, 776 cells analyzed; SCF, 4.74 ± 1.91, 13 coverslips, 1304 cells analyzed). Representative profiles of [Ca2+]i variations along the SCCI protocol are depicted in Fig. 6B. As shown, oligodendrocyte-like responding cells were hardly found in control cultures. On the contrary, when treated with T3, most cells were insensitive to KCl or histamine, responding markedly to thrombin, thus confirming the shift to an oligodendrocyte phenotype. On the other hand, in SCF-treated cultures displaying a neuronal tendency with many KCl-responding cells, and in CNTF-treated cultures exhibiting a typical astrocytic tendency with many nonresponsive cells, the oligodendrocyte population was unaffected and similar to levels in control cultures. Additionally, SCCI experiments were performed in defined cultures of hippocampal neurons as well as in cultures of cortical astrocytes (data not shown). Both cultures presented no oligodendrocyte-like responding cells with the established ratio Throm/Hist >1.3 (hippocampal neurons, 0.00 ± 0.00, 8 coverslips, 243 cells analyzed; cortical astrocytes, 0.00 ± 0.00, 7 coverslips, 305 cells analyzed).
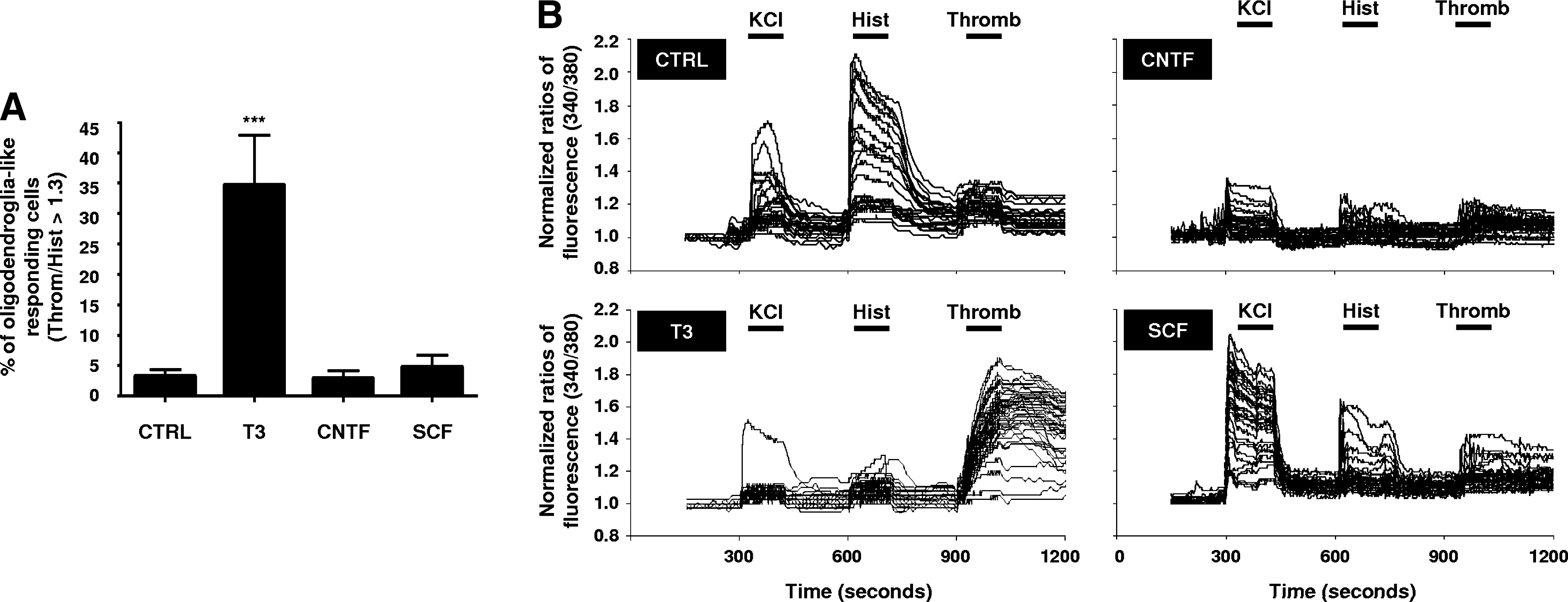
Functional evaluation of oligodendrocyte differentiation in subventricular zone (SVZ) cultures. (
We also investigated if the response to thrombin is exclusive to oligodendrocytes derived from the neurogenic SVZ. For that, we prepared neurospheres from early postnatal cortex, because it has been reported that at this stage the cortex contains astrocytes with stem cell potential. 35 The neurospheres were seeded on microgrid coverslips, incubated in the absence or the presence of T3, and evaluated under SCCI and immunocytochemistry procedures. Cortical and SVZ-derived PLP+ oligodendrocytes responded similarly to thrombin (data not shown). However, we also observed some cortex-derived thrombin-sensitive cells that responded to histamine. These data raise interesting questions about the influence of chemical/cellular environment in cell fate determination and functional responses of differentiating oligodendrocytes.
Discussion
We have described here a novel method to functionally evaluate oligodendroglia differentiation in SVZ cultures. The method consists in measuring the intracellular Ca2+ currents evoked by KCl, histamine, and thrombin on the different cell types present in a SVZ culture, and is based on the observation that each cell type displays a distinguishable profile of [Ca2+]i oscillations during the stimulation protocol. The main objective of the present work was to disclose whether thrombin is a Ca2+-mobilizing stimulus for oligodendrocytes in SVZ cultures. Because oligodendrocytes are rare cells in SVZ cultures, we treated the SVZ spheres with T3 after deposition in poly-
Using SVZ cultures differentiated on microgrid coverslips, we correlated the Ca2+ profiles with subsequent phenotypic immunodetection. We were enable to show that: (1) Similarly to astrocytes, O4+ and PLP+ oligodendrocytes do not respond to KCl or histamine; (2) [Ca2+]i is not altered by thrombin in MAP-2+ mature and DCX+ immature neurons, as well as in GFAP+ astrocytes; however, (3) O4+ and PLP+ oligodendrocytes display an increase of [Ca2+]i following thrombin perfusion. Cells presenting a Hist/KCl ratio between 0.9 and 1.1 are glial cells, as shown in our previous work, astrocytes, or oligodendrocytes, the latest being identifiable by their specific rise of [Ca2+]i under thrombin perfusion and related Throm/Hist ratio above 1.3. Interestingly, the Throm/Hist ratio characteristic of O4+ oligodendrocytes is variable, and this can be explained by the wide range of oligodendrocyte differentiation stages where O4 sulfatide is present. Moreover, we verified that NG2+ cells have either no response or a variable response to thrombin, which may suggest that PAR-1 expression begins gradually in the stage of progenitors of preoligodendrocytes. At a latter differentiation stage, a clear response to thrombin was observed in O4+ and especially in PLP+ oligodendrocytes. According to these findings, thrombin stimulation was identified as a reliable strategy to discriminate oligodendrocytes among other cells in SVZ cultures.
Using PAR-1–specific ligands, we concluded that a thrombin-triggered [Ca2+]i increase in SVZ-derived oligodendrocytes is mediated by PAR-1 activation. Indeed, PAR-1 immunoreactivity strongly co-localizes with O4 staining, labeling intensively in the oligodendrocyte cell body, and also appearing in clusters on cell processes. A recent study has detected functional PAR-1 receptors in Schwann cell myelin microvilli at the nodes of Ranvier. The authors concluded that PAR-1 activation in Schwann cells leads to a conduction block in the axon, presumably by an influx of calcium and subsequent release of substances that affect the propagation of the action potential. 39 Nevertheless, PAR-1 expression is not restricted to oligodendrocytes. Some astrocytes, and occasionally some Nestin+ immature cells, also express PAR-1. The scarce Nestin+/PAR-1+ cells presented a typical astrocytic morphology, suggesting that expression of PAR-1 in astrocytes begins early in their development when they still express the Nestin marker of immaturity, but are already committed to become astrocytes. In contrast with the robust PAR-1 labeling in the oligodendrocytes cell body, and their high peak of response to thrombin stimulus, these occasional Nestin+ and GFAP+ cells expressing the receptor presented only a dispersed membranas, rather than clustered, punctuated PAR-1 staining, indicating minor expression. No GFAP+ or Nestin+ cells responding to thrombin were found in SCCI analysis, therefore suggesting that low and dispersed expression of PAR-1 makes it insufficient to signal a detectable response by SCCI, or, alternatively, that PAR-1 is not coupled to an increase of [Ca2+]i in these cell types. On the contrary, mature neurons stained with MAP-2 did not express PAR-1. Previous studies have shown the presence of PAR-1 in motor neurons, olfactory neurons, and postnatal hippocampal neurons. 40 –42 Moreover, PAR-1–induced low Ca2+ transients were observed in rat hippocampal neurons. 42,43 However, in our experiments, we observed neither PAR-1 expression in SVZ-derived neurons, nor [Ca2+]i changes in these cells, or in embryonic hippocampal neurons (data not shown). It is worthwhile to mention that the observed Ca2+ transients in hippocampal cultures reported by Gorbacheva et al. 42 are about 200 nM, a very modest response. Furthermore, Yang et al. 43 demonstrate the requirement of a 24 h pretreatment of hippocampal neurons with thrombin to induce calcium response. Also, some authors report the expression of PAR-1 and Ca2+-associated response upon PAR-1 stimulation of newborn rat or mice cortical astrocytes. 44,45 We focused on cortical astrocytes from P7 mice and SVZ-derived astrocytes from newborn mice, the last occasionally having PAR-1 expression, but both being insensitive to thrombin stimulation (data not shown). Differences in culture conditions or age of the animals may be responsible for the different results in PAR-1 expression and thrombin response regarding cortical astrocytes.
Concerning the mechanism of PAR-1 downstream signaling toward cytosolic Ca2+ rise, our data demonstrate that PAR-1 binding leads to PLCβ activation through Gq/11 heteromeric protein activity. PLCβ hydrolyzes phosphoinositides generating inositol 1,4,5-triphosphate (IP3) and diacylglycerol (DAG), with consequent IP3-dependent Ca2+ release from the endoplasmic reticulum and PKC activation by DAG. This is consistent with previous studies examining the PAR-1 downstream signaling pathway in oligodendrocytes, 11 although not so far characterized in SVZ-derived oligodendrocytes.
Finally, we validated the proposed method by comparing the percentage of cells with Throm/Hist ratio above 1.3 (oligodendrocytes) in nontreated SVZ cultures, T3-treated cultures, as well as in CNTF- and SCF-treated cultures. As compared to nontreated cultures, Ca2+ profiles in treated cultures completely shifted differently, reflecting the acquisition of different phenotypes. As expected, T3 induced an increase in the proportion of oligodendrocyte population, whereas treatment with CNTF or SCF conferred no changes in the proportion of oligodendrocytes, further confirming the efficiency of the method.
The unique development of this novel strategy to functionally identify oligodendrocyte differentiation resides on the specific stimulation of this cell type with thrombin, acting through PAR-1 receptors. Moreover, we describe that PAR-1 receptor activation triggers the oligodendroglial increase of the [Ca2+]i, involving PLC-induced recruitment of calcium from internal stores. As a proof of concept, we treated SVZ cultures with T3, SCF, or CNTF to enrich the cultures in oligodendrocytes, neurons, or astrocytes, respectively. In agreement with our working hypothesis, the resulting profile of population responses revealed the efficiency of the procedure. In conclusion, the technological platform described in this study is relevant in the context of drug discovery and development in the general area of regenerative medicine of the brain.
Footnotes
Acknowledgments
We thank Dr. João Relvas for scientific advice and Dr. Elisabete Ferreiro and Dr Cláudia Pereira for their gift of U73122. This work was supported by FCT-PTDC/SAU-NEU/68465/2006 and SFRH/BD/32953/2006.
Author Disclosure Statement
The method described in this work is under patent protection (U.S. Provisional 16/10/2074-PAT 2008100089504/0198).