
Abstract
A number of mammals undergo hibernation during their lifespan. The euthermal condition that occurs prior to hibernation is characterized by inhibition of cellular proliferation, reduction of stress reactivity, a decrease in metabolism, and changes in the immune system (seasonal thymus involution). Analysis of data of young red voles (Clethrionomys glareolus), common shrews (Sorex araneus), and fat dormice (Glis glis) while entering into the euthermal condition shows characteristic biphasic growth with a delay that can vary in its periods. A biological mechanism for reversible growth arrest effectively “stops the clock” of these animals. It is suspected that this mechanism is triggered by an increase in somatostatin synthesis with inhibition of growth hormone, thyroid-stimulating hormone, and melatonin, and with a loss of circadian function in the suprachiasmatic nuclei of the hypothalamus.
Introduction
Fat dormice Glis glis, small rodents of the Gliridae family, occupy an extensive area around all of Europe and its adjoining territory (Caucasus, Asia Minor, and the Kopet Dag, a mountain range between Turkmenistan and Iran). Fat dormice are obligatory hibernators, with a long (up to 8 months per year) hibernation period. 4,5 Juvenile fat dormice stop growing shortly before hibernation. Their reversible growth inhibition in a euthermal condition can be prolonged for a variable period (for more than 1.5 years in our experiments). Because the animals retain juvenile features during the time of growth delay, including size and the type of pelage (hair) that does not change throughout molting, we consider this a reversible arrest of the animals' “ontogenesis clock”—a “freezing” of their biological age.
We first noted in 2004 that young fat dormice that were exposed from the moment of birth to a natural photoperiod indoors in temperature 18–25°C would stop growing in autumn and winter and would remain at a developmental stage resembling 2-month-old individuals. We decided to probe this phenomenon.
Materials and Methods
Animals
Animals used in the present study belonged to a subspecies of fat dormice Glis glis glis Linnaeus 1776 and were derived from pregnant females caught in the Matveevski Forest (in the Chigirin area of the Cherkasy region; license #198 of the Ministry of Environmental Protection of Ukraine).
Two groups of mixed sexual composition were formed from a number of young animals (18 subjects, aged 1 month), born during the period from July 26 until August, 1, 2005: Group 1 (8 subjects) and group 2 (10 subjects), which were housed in two rooms, called modes B (group 1) and A (group 2). Six months later, subjects from group 2 were sorted into the followings groups (5 subjects in each), contained in mode B (group 3) and in mode A (group 4). We obtained 13 young subjects during period from July 1 to July 14, 2007 and used them in research in during the period 2007–2008.
Fat dormice were housed in spacious cages 490 × 470 × 300 mm made of zinc–steel net with 20-mm cells, equipped with suspended nest boxes (100 × 100 × 150 mm) constructed from multilayered plywood. Animals were maintained on a waterless diet with free access to food. Different fruits (mainly apples), oat flakes, dried fruits, protein granular fodder (Nutra Nuggets Professional for Cats; 32% protein, 21% fat, 3% carbohydrates [cellulose], 5.5% ash, 10% humidity) completed their rations.
The work was carried out in compliance with the principles of the Helsinki Declaration on Humane Treatment of Animals.
Light and temperature regimes (modes)
The following regimes were employed: (A) Natural light mode (NL), which constituted illumination through the windows, on the latitude 49°N. Bright light was light exposure at a level of cages in the afternoon 500–1000 lux, temperature 18–25°C, humidity 50–70%. (B) Dim (having no connection with the season of the year) light mode (DL), was natural light plus additional artificial illumination after dawn, with lights on during the evening for on average 15 h (minimal duration was 12 h, maximal was 18 h), soft light (luminescent lamps of a daytime spectrum). Light exposure was 100–300 lux, temperature 18–25°C, humidity 50–70%. (C) Light deprivation (DD) was light exposure less than 1 lux during the day, temperature 18–25°C, humidity 50–70%. (D) Overwintering in conditions of light deprivation included illumination less than 1 lux, temperature 2–5°C, humidity 90–100%. (E) Overwintering in conditions of natural illumination included illumination through the windows, bright light (500–1000 lux), temperature 2–10°C, humidity 60–80%.
Testing reversible growth arrest
For testing RGA of the dormice during the experiment, the length of the head (Lc) was measured every 2 weeks, because it provides a good estimate of the size of the skull and is more accurate than the measurement of body length in live animals. Measurements of Lc were made with a special ruler with a staple fixed on the end, from the most protruding part of an occipital bone to the tip of the nose, with accuracy ±0.5 mm. Body weight as a parameter of age changes in dormice is poorly informative because of significant seasonal changes in adiposity. 6
In addition to head measurements, age-related changes in growth and development (“biological age”) were monitored by recording the type of pelage as juvenile or adult. The first cycle of growth of a juvenile pelage is completed by 1 month of age and produces a coat of very short and soft hair. Juvenile pelage consisting of longer hair is formed during the second month of postnatal development.
Statistical analysis
Statistical analyses were conducted using SPSS 16.0 for Windows. Data were expressed as mean ± standard error of the mean (SEM). The Student t-test and one-way analysis of variance (ANOVA) test were used to compare the differences in Lc of dormice between groups 1 and 2 and groups 3 and 4. The level of significance was set at p < 0.05. Graphs were plotted using the master of diagrams in Excel 2003.
Results
During the first month of exposure to regimes A and B, the growth rate of the animals (estimated by measurements of head length) did not differ between groups 1 and 2 (38.53 ± 0.41 mm, t = 1.78, p > 0.05) (Fig. 1A). At the beginning of October, 2005, growth of young dormice in group 2 had almost stopped, whereas in group 1 an increase in the length of the head was observed for 1.5 months, reaching final values of 42.50 ± 0.42, F(2, 14) = 12.3, p < 0.001), and change of the pelage from juvenile to definitive had begun (Fig. 1A). The molting in group 2 started at the age of 7 months (the type of pelage had not changed), without any changes in the linear size of the animals.
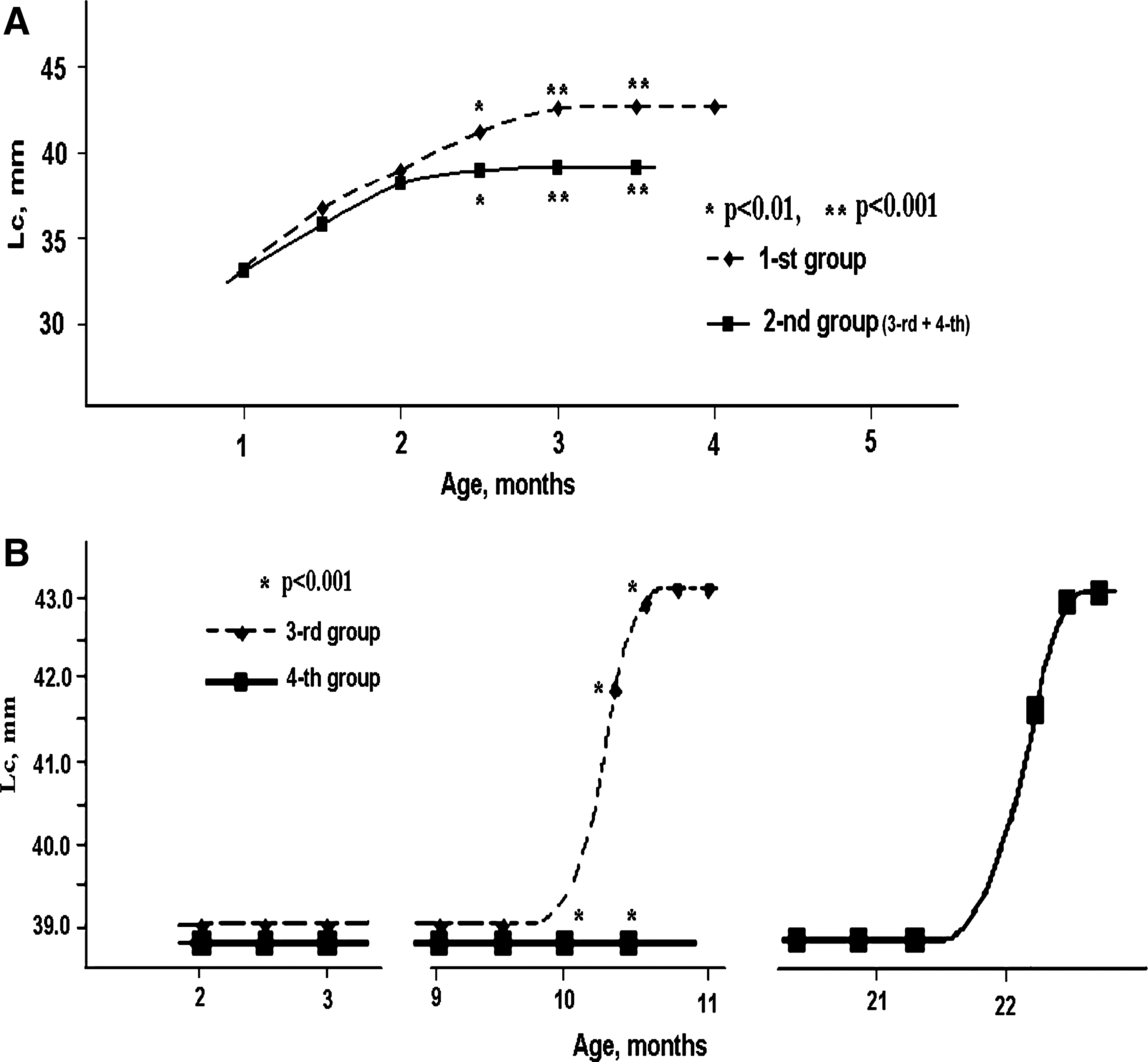
Length of the head (Lc) in the fat dormouse as a marker of reversible growth arrest (RGA). (
On March 25, 2006, when the animals were approximately 8 months old, group 2 was randomized into two groups (3 and 4) of 5 animals each. Group 4 remained in the conditions of mode A, whereas group 3 was moved to a location with mode B (Fig. 1B). After a little more than 2 months (May 30, 2006) in conditions of varied light mode, group 3 had its RGA interrupted, as indicated by a significant increase in head size (42.73 ± 0.30, F(2, 8) = 21.7, p < 0.001) (Fig. 1B). On February 15, 2007, animals from group 4 were placed in hibernation mode (mode D) and were in a condition of hypothermia until May 5, 2007.
In the first days after a switch from wintering to mode A, an increase in the Lc was detected in all 5 animals. Simultaneously with renewal of growth, the juvenile pelage started to change into the definitive pelage. Thus, in these 5 individuals, the RGA was maintained for 16.5 months (from the beginning of October, 2005, to February 15, 2007), and its interruption provoked hibernation.
From July 1 to July 14, 2007, 11 newborn dormice were obtained. These animals were maintained in mode A from their date of birth. Delay of growth (occurrence of RGA) was noted at the beginning of October, 2007. Induction of hibernation was carried out in mode E from December 19, 2007. Starting on January 10, 2008, 2 subjects that failed to enter hibernation were placed in a lightproof box, where they remained until January 20 (10 days). As a result of maintenance in conditions of light deprivation (DD, light modes C), there was rapid onset of RGA in 1 individual (increase in the Lc up to 41 mm [ + 3 mm] and the fur changed into the definitive form), whereas in the other animal, the exit from RGA had a latent period of 1.5 months.
Removal of the basic group of animals from hibernation was begun on February 4 by increasing the temperature to 22°C. Within 3–5 days, the activity of the animals was completely restored. Of the 9 animals that stopped hibernation at this time, interruption of the RGA condition in the first days after arousal was noted in 1 individual (11.1 %). In 7 animals (77.7%), interruption of growth inhibition was terminated between March and May, and RGA was maintained until the following autumn in 1 dormouse (11.1%).
Discussion
On the basis of the present findings and previously published data, it can be proposed that the mechanism of RGA in mammals is activated by various exogenous factors, including photoperiod durations, light exposure and temperature, availability of food, and density of population. The high level of light exposure promotes RGA because, on the one hand, it increases the influence of the photoperiod length on the organism, and, on the other hand, it promotes reduction of melatonin synthesis.
Our results indicate that the exit from a condition of growth inhibition is realized in various conditions (hibernation, light deprivation, dim light mode) under various regimens (within several days or with the latent period, up to 2 months). The increase in melatonin can be the key factor in the interruption of a condition of the RGA (melatonin concentration in hibernating mammals increases intermittently during their periodic arousals).
Characteristics for hibernating mammals are clearly defined by a major restructuring of the organism's physiological and endocrine functions in the period prior to hibernation, including inhibition of melatonin production, thyroid-stimulating hormone (TSH) and growth hormone (GH) synthesis, decrease of cellular proliferation, changes in the immune system (seasonal involution of thymus), and an overall decrease of metabolism. 7
The growth arrest in young mammals is a possible consequence of total inhibition of TSH, GH, and melatonin synthesis. One basic factor of this transition is probably somatostatin (SST). SST synthesis increases in the hypothalamus as environmental conditions deteriorate. SST inhibits synthesis of many hormones, including serotonin (the melatonin predecessor), TSH, and GH, and also cellular proliferation. 8,9 This assumption has been confirmed by data showing a high level of SST concentration in the hypothalamus of hibernating mammals during hibernation. 10
We propose considering “neobiosis” (a new term) as a euthermal condition of mammals preceding hibernation or substituting for it. Neobiosis is accompanied by elimination of the age-related (i.e., irreversible) changes of an organism. The convertibility of changes that would be irreversible in a majority of mammals is a characteristic of neobiosis and takes place during development (e.g., growth arrest, involution of thymus).
What are the evolutionary preconditions of neobiosis? We consider that the rate at which an organism develops is maximal in an optimal environmental zone that, for the given organism, is defined by environmental parameters, including physical, chemical, and biological factors. The deviation of these parameters values from optimum leads to a reduction of the speed of development and an increase in the death rate that is unrelated to age but related to a reduction of adaptation. A decrease in rates of individual development under deteriorating environmental conditions is the basic possible precondition for the emergence of neobiosis during evolution.
In sum, neobiosis in mammals with a short life can be described as an adaptive mechanism as a result of natural selection allowing support for a population for long periods (winter, drought) that are unfavorable for reproduction by a significant increase in life expectancy of young animals within the population. It is important, to note that the increase in lifespan is achieved by prolongation of run up to definitive animal size and puberty.
What are the central mechanisms responsible for the neobiosis? It is hard to imagine that the processes of individual development of an organism are not coordinated by a common center, just as an orchestra is controlled by a conductor. Perhaps the answer to this question can be explored by studying the hibernation of mammals. In some species of mammals, duration of hibernation may be more than half a year. Hibernation is an interrupted process, consisting of periods of torpor (hypothermia) and periodic arousals. The biological significance of periodic arousals—the rapid heating of animals—is still not clear. Most likely, animals control the environment and the removal from the organism products of metabolism. A circadian rhythm is not characteristic for periodic arousals. In the same animal during hibernation, the length of the waking state and the periods of hypothermia among them can change. In addition to hibernation to hypometabolic states, mammals can be called during the daily torpor; this occurs in many species (dormice, bats, hamsters). The main difference between daily torpor and hibernation is to maintain the daily rhythmic activity and periods of rest of the animals. The state of daily torpor constitutes energy savings at the time of day when the animals rest.
The circadian rhythm of activity and rest in mammals is controlled by the suprachiasmatic nuclei of hypothalamus (SCN), thus it is not difficult to conclude that the circadian function of SCN is counterproductive during hibernation. Its retention would serve as an “alarm” that compels animals to wake up every day at about the same time. Research profiles of “clock” gene expression in mammals during hibernation and daily torpor confirm this viewpoint, suggesting that the molecular mechanisms of the “circadian clock” stop during hibernation, but continue to work in a state of daily torpor. 11,12
We postulate a hypothesis in which neobiosis is a “by-effect” of the circadian clock stop in hibernating mammals. In most species, neobiosis is not age-related. However, in short-lived species (voles, shrews), neobiosis represents an adaptive mechanism observed only in young animals. Once the mechanisms of neobiosis are identified, it may be possible to try to modeling neobiosis in laboratory animals, with the aim of prolongating their lifespans. Hypothetically, it could be possible to create such a method of lifespan extension in humans (ideally, to achieve negligible senescence).
Footnotes
Acknowledgments
This research work was carried out within the framework of the project “Practical Use of Skulachev's Ions,” MSU. We thank Academician V.P. Skulachev, Prof. V.N. Anisimov, Prof. A.A. Bartke, and Dr. A.M. Olovnikov for help and support.