
Abstract
The local renin–angiotensin system (RAS) in the brain is a multitasking system controlling a plethora of essential functions such as neurogenic hypertension, baroreflexes, and sympathetic activity. Aside from its vasoactive actions, brain angiotensin II (AT-II) has also been implicated in the pathogenesis of cognitive decline, and beneficial effects of angiotensin receptor blockers (ARBs) in Alzheimer (AD) and Parkinson diseases (PD) are suggested. However, the use of ARBs at antihypertensive dosages would lead to unwanted hypotensive reactions in AD patients. Here we treated the APP/PS1 transgenic mouse model of AD with the ARB losartan (10 mg/kg body weight) to determine whether blockade of the AT-II receptor subtype 1 (AT1-R) with intranasal losartan, using at a dosage far below its systemic antihypertensive dose, could maintain its neuroprotective effects independent of its systemic vasoactive action. Intranasal losartan treatment (10 mg/kg every other day for 2 months) of APP/PS1 mice decreased amyloid β (Aβ) plaques 3.7-fold. Blood serum levels of interleukin-12 (IL-12)p40/p70, IL-1β, and granulocyte-macrophage colony-stimulating factor (GM-CSF) were increased in the vehicle-treated APP/PS1 mice. Intranasal losartan not only decreased IL-12p40/p70, IL-1β, and GM-CSF, but also increased IL-10, which suppresses inflammation. Furthermore, losartan markedly increased tyrosine hydroxylase expression in the striatum and locus coeruleus. In conclusion, losartan exerts direct neuroprotective effects via its Aβ-reducing and antiinflammatory effects in the central nervous system (CNS). Therefore, intranasal losartan and potentially other ARBs, at concentrations below their threshold for altering systemic blood pressure, offer a new approach for the treatment of AD.
Introduction
Here we show that chronic intranasal losartan treatment decreases amyloid β (Aβ) plaque formation, blood serum inflammatory cytokines, and catecholaminergic activity reflected by tyrosine hydroxylase (TH) expression independently from its systemic antihypertensive action.
Materials and Methods
Double transgenic APP/PS1(APPswe/PS1dE9) female mice expressing a chimeric mouse/human amyloid precursor protein (Mo/HuAPP695swe) and a mutant human presenilin 1 (PS1-dE9), both directed to CNS neurons, and their wild-type littermates were obtained from Jackson Laboratories (Bar Harbor, ME). Animals were housed individually in the central animal facility with controlled temperature and an artificial light/dark cycle. Food and water were available ad libitum. Animal experiments were performed in accordance with the guidelines of the University of Tübingen for the experimental use of animals and German law for the protection of animals. All experiments were approved by the local Animal Protection Committee, University of Tübingen.
Seven month-old mice were divided into losartan- and vehicle-treated (0.9% NaCl) APP/PS1 groups and a vehicle-treated wild-type (WT) control group (n = 8 in each). Ten days prior and during the first 10 days of losartan treatment, blood pressure was measured by the tail cuff method using a blood pressure analyzer (Hugo Sachs, March-Hugstetten, Germany) as described elsewhere. 12 Losartan (MSD, Haar, Germany) dissolved in in 24 μL of 0.9% NaCl was applied intranasally (10 mg/kg body weight) every other day for a period of 2 months. Thereafter, the mice were sacrificed under ketamine anesthesia (75 mg/kg, intraperitoneally [i.p.]; Delta Select, Pfullingen, Germany). The brains and serum samples were frozen at −80°C and processed as described below.
Horizontal cryosections (10 μm in thickness) were prepared from the brains of mice and fixed with methanol at −20°C, washed, and incubated with antibody against the human sequence of Aβ (mouse monoclonal 6E10 antibody, dilution 1:100; Signet, Dedham, MA) or TH (mouse monoclonal, dilution 1:200, Millipore, Billerica, MA) and glial fibrillary acidic protein (GFAP, rabbit polyclonal, dilution 1:400; DacoCytomation, Glostrup, Denmark) for 2 h at room temperature (RT). Sections were washed with phosphate-buffered saline (PBS) and further incubated with the respective Cy3-conjugated goat anti-mouse (diluted 1:500) or fluorescein isothiocyanate (FITC)-conjugated anti-rabbit immunoglobulin Gs (IgGs, 1:100, Dianova, Hamburg, Germany) for 1 h at RT. Thereafter, samples were washed with 0.1% Triton (Sigma, Taufkirchen, Germany) in PBS, coated with Vectashield mounting medium (Vector Laboratories Burlingame, CA) containing 4′,6 diamidino-2-phenylindole (DAPI) and assessed by fluorescent microscopy. The quantification of Aβ plaques (total surface and number of plaques) was performed in the hippocampal area from 10 sections between −3 to −6 mm from bregma using fluorescence microscopy and the CellP software (Olympus SIS, Hamburg, Germany).
The measurement of interleukin-α (IL-1α), IL-1β, IL-4, IL-5, IL-6, IL-10, IL-12p40/p70, tumor necrosis factor-α (TNF-α), interferon-γ (IFN-γ; granulocyte-macrophage colony stimulating factor, GM-CSF) was performed with a Beadlyte® mouse Multi-Cytokine BeadmasterTM Kit (Upstate, Lake Placid, NY) and a Luminex-100 system (Luminex Corporation
Results and Discussion
Measurement of systolic (SBP) (Fig. 1A) and diastolic blood pressure (DBP) (Fig. 1B) 10 days prior to and during the first 10 days of losartan treatment showed no significant difference between the groups (cf. losartan- and 0.9% NaCl-treated groups in Fig. 1A,B). The increase in formation of Aβ plaques, previously reported to begin from 6 to 7 months of age, 4 was observed in 9-month-old APP/PS transgenic mice with a 13- and 17-fold increase in the number and total surface area of plaques, respectively (cf. WT vs. control in Fig.1C,D). Two months of intranasal losartan treatment decreased both the total number (Fig. 1C, cf. losartan with control) and surface area of Aβ plaques (Fig. 1D, cf. losartan with control) in the hippocampus. Immunohistochemical analysis of Aβ and GFAP staining (Fig. 2A–F) shows a strong expression of both Aβ and GFAP in APP/PS1 control groups in the hippocampus (Fig. 2E) and entorhinal cortex (Fig. 2B), whereas the levels of Aβ and GFAP in the losartan group (Fig. 2A,D) are nearly equal to those of the WT controls (Fig. 2C and F). Among the cytokines detected by multiplex analysis, IL-1β, IL-12p40/p70, and GM-CSF were elevated in the APP/PS1 controls in comparison with the WT animals (cf. control vs. WT in Fig. 3A,C,D) whereas only negligible amounts of IL-1α, IL-4, IL-5, IL-6, TNF-α, and IFN-γ were observed in all analyzed groups (data not shown). Intranasal losartan decreased IL-1β, IL-12p40/p70, and GM-CSF in the blood serum of APP/PS1 mice (cf. losartan vs. control in Fig. 3A,C,D) down to the levels observed in WT mice (WT in Fig. 3A,C,D). Interestingly, in the intranasal losartan-treated group, IL-10 was significantly upregulated in comparison with vehicle-treated transgenic and WT control animals (cf. losartan vs. control and WT in Fig. 3B). A prominent increase in TH expression was seen after intranasal losartan treatment in both the striatum (Fig. 4A) and locus coeruleus of the APP/PS1 mice (Fig. 4D), whereas the vehicle-treated APP/PS1 mice (Fig.4B,E) had reduced TH intensity than that of the WT controls (Fig. 4C,F).
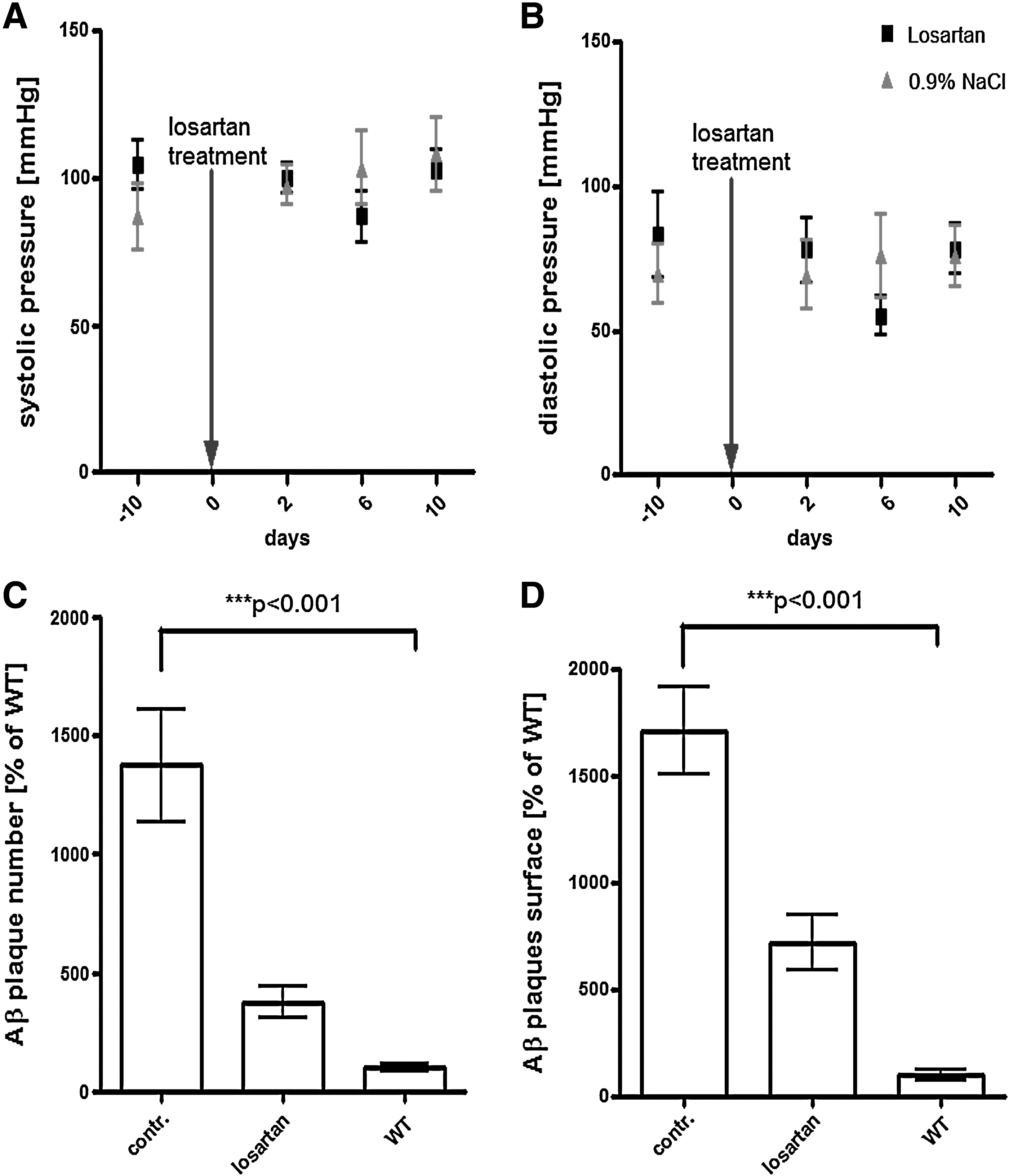
Blood pressure measurement and quantification of Aβ plaques in APP/PS1 mice
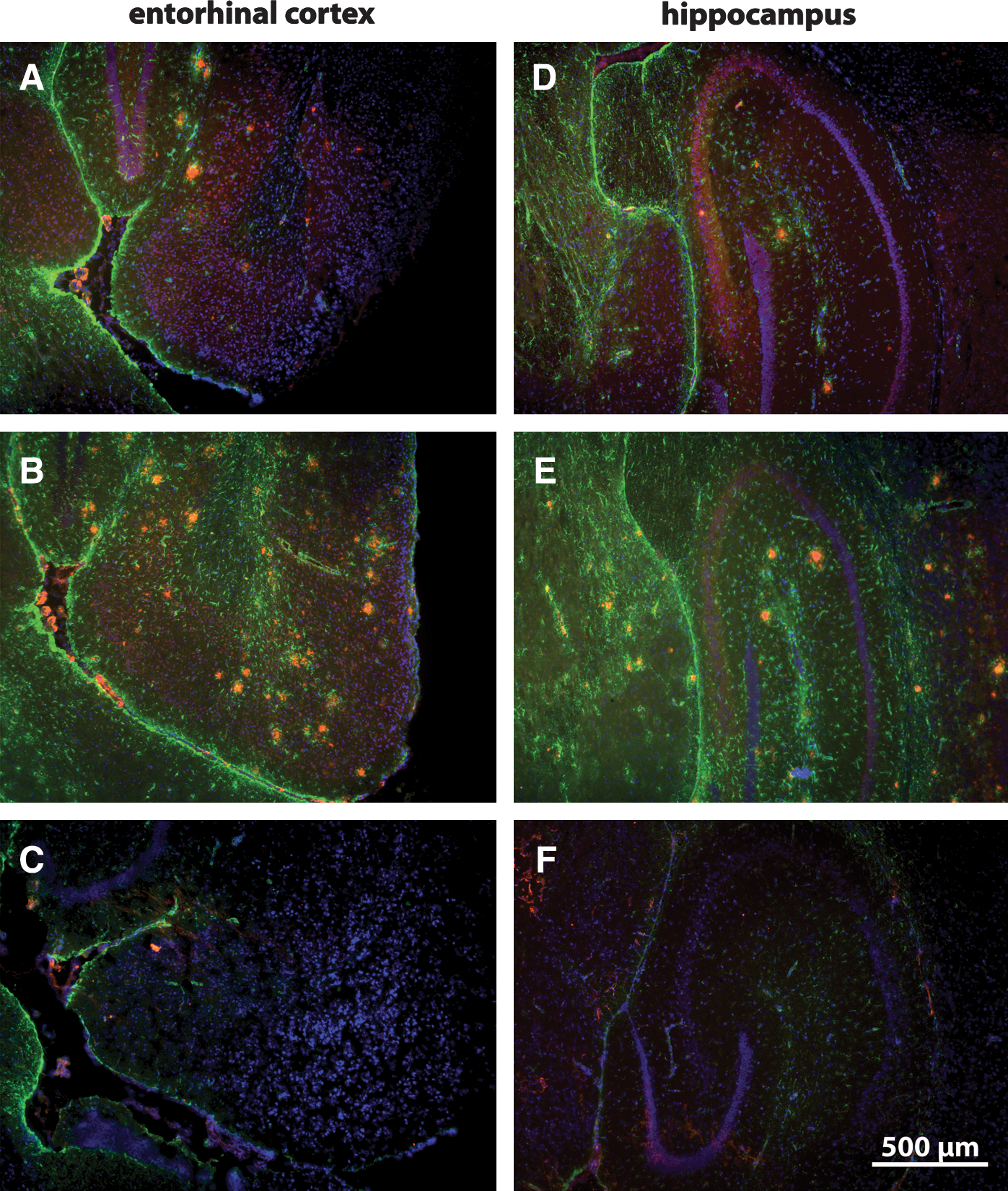
Expression of Aβ and glial fibrillary acidic protein (GFAP) in the entorhinal cortex and hippocampus of APP/PS1 mice. Immunostaining of Aβ (red) and GFAP (green) in the respective entorhinal cortex and hippocampus of losartan- (

Cytokine analyses in the blood serum of APP/PS1 and wild-type (WT) mice. Quantification of granulocyte-macrophage colony-stimulating factor (GM-CSF) (
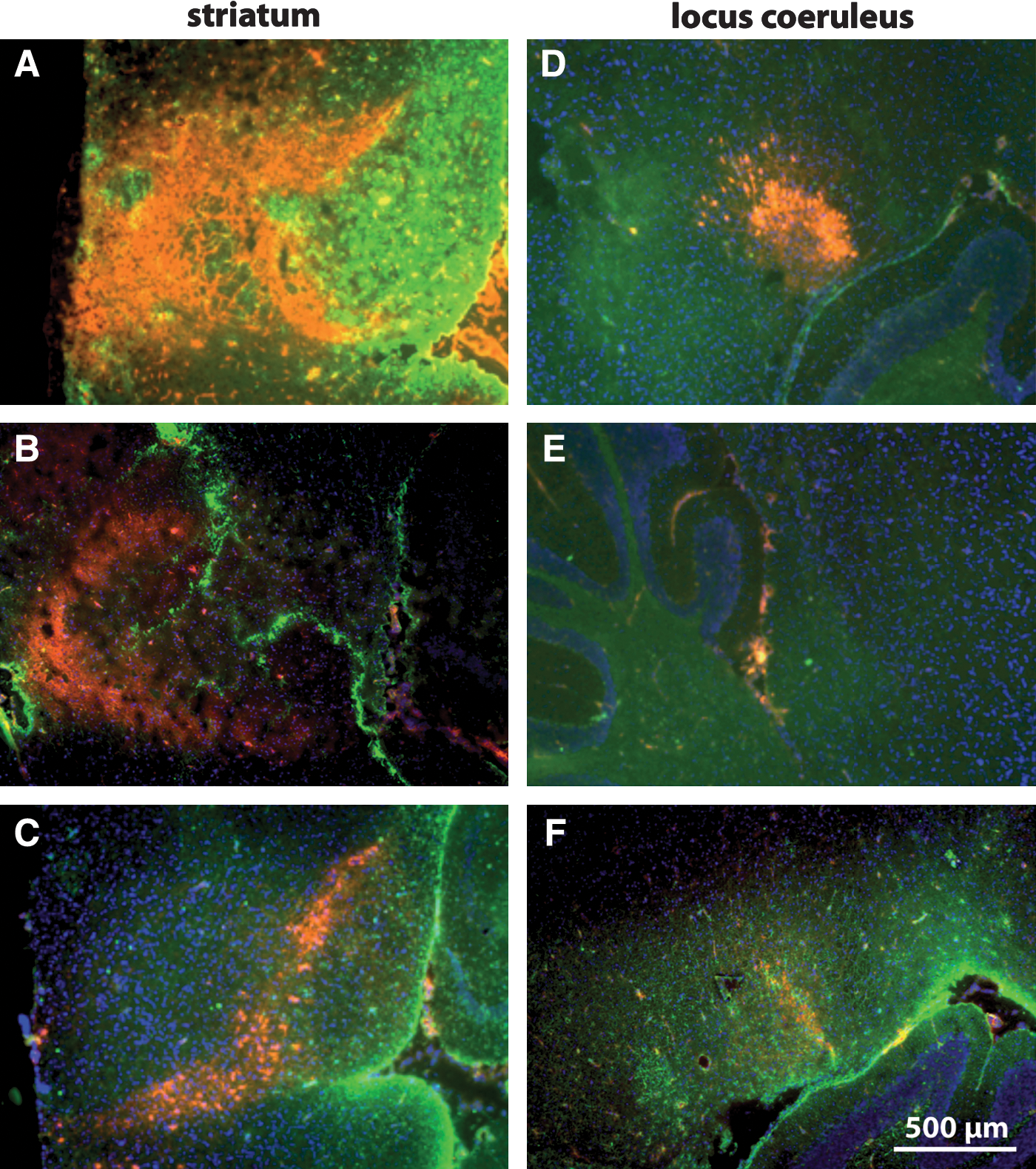
Expression of tyrosine hydroxylase (TH) and glial fibrillary acidic protein (GFAP) in APP/PS1 and wild-type mice. TH (red) and GFAP (green) in the striatum and locus coeruleus of losartan-treated (
Regarding the extensive amount of experimental and clinical trials in the past, the implication of antihypertensive drugs targeting the RAS indicates a promising future strategy in treatment of neurodegenerative disorders. However, treating normotensive patients with blood pressure-lowering drugs to achieve neuroprotection and improvement in memory function would result in unwanted hypotensive reactions and other systemic side effects. 14 Intranasal application not only avoids systemic side effects but also achieves a more targeted delivery of losartan to the brain in comparison with systemic administration. Similar benefits have been described by Hanson et al. 15 for intranasal deferoxamine, which targets deferoxamine to the brain and does not reduce blood pressure even though intravenous deferoxamine is known to lower blood pressure.
Inflammation is an important contributor to the pathology of AD. Intranasal losartan decreases the concentrations of IL-1β, IL-12, and GM-CSF in blood serum of APP/PS1 transgenic mice. The elevated levels of IL-1β, IL-12, and GM-CSF in blood serum of vehicle-treated APP/PS1 mice in comparison to WT controls correlate with the elevated levels of these proinflammatory cytokines in the brain reported previously by Patel et al. 16 Therefore, measurement of these cytokines in the blood of APP/PS1 transgenic mice may serve as a useful marker of neuroinflammation in this model of AD. APP/PS1 mice fail to counteract the elevated proinflammatory cytokines (IL-1β, IL-12, and GM-CSF) induced by Aβ pathology with the antiinflammatory IL-10 and IL-4, whereas losartan increases the level of IL-10. Furthermore, the antiinflammatory capacity of intranasal losartan is reflected by significant decreases of IL-1β, IL-12, and GM-CSF. Antiinflammatory features of ARBs were also shown previously by release of immunosuppressive cytokines such as IL-10 and transforming growth factor-β (TGF-β) from CD4+ T cells and CD11b+ monocytes in a candesartan-treated animal model of multiple sclerosis 17 and by decrease in TNF-α release from losartan-treated astroglia in an in vitro model of hypoxic injury. 18
Chronic intranasal losartan treatment dramatically decreased plaque number and their overall surface area. Recent studies show the Aβ-lowering activities of ARBs in vitro 19 and in vivo. 20 ARBs generally may be more effective in terms of decreasing or preventing Aβ production and plaque formation than ACEIs because they do not inhibit ACE, the enzyme capable of not only converting angiotensin to its vasoactive form but also Aβ1–42 to Aβ1–40, which is known to be less neurotoxic than Aβ1–42. 21
TH is the first rate-limiting catecholaminergic enzyme in the dopamine and noradrenalin biosynthetic pathway and a reliable marker of dopaminergic neurons. The ability of losartan to increase TH in both striatum and locus coeruleus hints at the regulatory role of AT1-R blockade on noradrenergic/dopaminergic systems, the activity of which is decreased in striatum and locus coeruleus of APP/PS1 mice, as shown in previous studies, 22,23 and confirmed by the results described above. Numerous studies suggest the beneficial effects of RAS-acting antihypertensive drugs in mood and stress disorders 24 (for reviews see refs. 25 and 26). The increase in TH expression shown here may explain the ability of the AT1-R blocker candesartan to increase the levels of noradrenalin in the CNS of rats reported by Jenkins. 27 The TH-increasing effect of losartan in striatum of APP/PS1 mice is concordant with data showing the protective effects of ARBs on dopaminergic neurons in an in vitro MPTP model of PD. 17
We can conclude from our results that, aside from its antihypertensive action, losartan shows Aβ-lowering and antiinflammatory activities in the APP/PS1model of AD. Furthermore, losartan is capable of increasing TH expression in the locus coeruleus and striatum and therefore may be of therapeutic value not only in AD but also in other neurologic disorders associated with depleted dopaminergic and noradrenergic activities such as PD and mood disorders. In addition, intranasal delivery is an alternative method of treatment that can help to avoid the undesirable systemic effects of ARBs. Future work will address the ability of intranasal ARBs to improve cognitive function in animal models of AD.
Footnotes
Acknowledgments
We appreciate the support of Microbionix (Regensburg, Germany) in performing of multiplex analysis. This work was supported by “Dr. Karl-Kuhn Stiftung” (Tuebingen, Germany).
Author Disclosure Statement
No competing financial interests exist.