
Abstract
We investigated the effect of the phytocompound Denshici-to-Chiusei (DTS) on the atherogenesis in apolipoprotein E−/−/low-density lipoprotein receptor−/− (apoE−/−/LDL receptor−/−) mice (E0). E0 mice were fed for 16 weeks with: (1) placebo or (2) 25 mg or (3) 50 mg of DTS/day. Aortic lesions were reduced by 38% (p < 0.01) in mice fed 50 mg/day, whereas peritoneal macrophages after both dosages had a 45%–60% lower (p < 0.01) capacity to oxidize LDL and to degrade it. This was associated with reduced LDL-associated lipoperoxides and a 22% inhibition (p < 0.05) in LDL aggregation. Tumor necrosis factor-α (TNF-α) expression and immunoreactivity in the aortic media increased five-fold, but this was significantly mitigated by DTS (50 mg > 25 mg) (p < 0.05). DTS significantly attenuated inflammatory mechanisms preceding atherogenesis with reduced LDL susceptibility to oxidation–aggregation.
Introduction
Materials and Methods
Male apolipoprotein E and LDL receptor gene double-knockout mice (apoE−/−/LDL receptor−/−) were fed standard chow. E0 mice (n = 30; 6 weeks old) were divided into three groups of 10 and fed for 16 weeks via their drinking water with the following: group A, 1.1% alcohol and water (control); group B, 25 mg of DTS/day; and group C, 50 mg of DTS/day in 1.1% alcohol and water. Mice were killed by intraperitoneal injection of Hypnorm:Dormicum in week 16. Their blood was immediately withdrawn, and entire aortas were surgically removed, washed in 0.9% saline containing 0.02 mM butylated hydroxytoluene, and fixed in 4% formaldehyde for 12 h.
Histopathology of aortic atherosclerotic lesions
The aorta was rapidly fixed in 3% glutaraldehyde in 0.1 mol/L sodium cacodylate buffer with 0.1 g/L calcium chloride, pH 7.4, at room temperature and then cut transversely into blocks of 1 mm each.
Peritoneal macrophage preparation and functional studies
Peritoneal macrophages (PM) were harvested from the peritoneal fluid of the E0 mice, washed, and centrifuged with phosphate-buffered saline (PBS) and then resuspended in Dulbecco modified Eagle medium (DMEM). The cell suspension was incubated for 2 h and washed with 5 mL of DMEM; the monolayer was incubated for 18 h before the experiment. PM were incubated for 5 h at 37°C with supplemented DMEM medium in the presence of 125I-labeled oxidized (Ox)-LDL (10 mg of protein/L). Macrophage degradation of the labeled lipoprotein was measured by trichloroacetic acid-soluble noniodide radioactivity. The cells were washed with PBS, and aliquots of 0.5 mL were taken for radioactivity counting to measure cell-associated lipoprotein. Oxidation of LDL was performed by incubating the LDL with CuSO4 (5 mmol/L). Oxidation was terminated by refrigeration at 4°C and addition of 0.1 mmol/L of Na2 EDTA to chelate the copper ions. LDL oxidation was determined by measuring thiobarbituric acid-reactive substances (TBARS). 10 Formation of conjugated dienes was monitored by measuring the increase in absorption at 234 nm. LDL-associated lipid peroxide formation was determined with a cholesterol color reagent, and LDL aggregation after vortexing was confirmed by observation at an optical density of 680 nm.
Determination of LDL oxidation susceptibility
LDL was isolated by a density-gradient ultracentrifugationin The total LDL cholesterol (LDL-C) level was measured using a commercial kit (Boehringer-Mannheim). The susceptibility to oxidation was tested by determinations of TBARS and conjugated dienes. Two hundred milligrams of LDL protein per liter was oxidized in the presence of 25 mol/ Cu2 in PBS for 6 h at 37°C. The lipoperoxide content of oxidized LDL was determined as above. Conjugated dienes in LDL were measured at 234 nm in a Perkin-Elmer UV-VIS spectrometer.
Immunohistochemistry
Tissue sections were blocked with 2–5% rabbit serum in PBS for 30 min. Primary antibodies against tumor necrosis factor-α (TNF-α or TNF receptors p55 and p75 were diluted in PBS and incubated at 4°C in a humidified chamber. Sections were washed and incubated with biotinylated secondary rabbit anti-goat antibody diluted in PBS and then washed. The sections were incubated for 30 min with a peroxidase-labeled avidin–biotin complex and then developed using a DAB Detection Kit and counterstained in Hematoxylin. Negative controls included substitution of the primary antibody with PBS. The TNF immunoreactivity was evaluated by image analysis and expressed as percent positive immunostaining of total media or plaque area. The mean medial staining and plaque area for each animal were based on four sections from each main district.
RNA isolation and cDNA synthesis
Total RNA was isolated from the aorta by applying the protocol for FastPrepTM system (Carlsbad, California). The synthesis of cDNA was performed by mixing 1 μg of total RNA with random primers, dithiothreitol (DTT), deoxyribonucleotide triphosphate (dNTP), RNase inhibitor (INH), and Moloney murine leukemia birus reverse transcriptase (Mo-MLV RT) (GibcoBRL).
Real-time polymerase chain reaction
TaqMan Universal PCR Master Mix (TM-PCR-MM, Applied Biosystems, USA) was used in an ABI PRISM 7700 Sequence Detection System (Perkin-Elmer, Applied Biosystems) for amplification of cDNA. Samples without Mo-MLV RT in the cDNA synthesis and wells with no template were used as controls. All samples were run in triplicate. Amplification of TNF-α and 18S rRNA was carried out by using cDNA and TaqMan predeveloped assay reagents and 2 × M-PCR-MM . Amplification of TNF (TNFα-R1 and TNFα-R2) p55 and p75 receptors was carried out by using appropriate concentrations of specific primer pairs and probes and 2 × TM-PCR-MM. The endogenous control 18S rRNA was used for normalization of data. Calculations were performed using standard curves obtained by serial dilutions of cDNA synthesized from RNA entracted from aortas of 7-month-old apoE−/− mice.
RT-PCR analysis of TNF-α mRNA
RNA samples prepared from the livers of oxidized LDL-treated animals served as positive controls for PCR analysis. Total RNA was purified by using RNAgents total RNA isolation system for reverse transcription (RT)–PCR (Promega); afterward each RNA sample was treated with DNase. Two micrograms of total RNA was reverse-transcribed by the random-priming method with Pd(N)6 hexamers (Pharmacia Biotech). Analysis of specific mRNAs by RT-PCR was accomplished using a Clontech amplimer set. An aliquot of each reaction underwent electrophoresis on 1.8% agarose gels and was analyzed by ethidium bromide staining. Quantification of TNF-α was expressed as the ratio of intensity of the TNF-α to the β-actin bands.
Results
Aortic atherosclerotic lesion areas were reduced 38% (p < 0.01) in mice that consumed 50 mg of DTS/day. DTS did not significantly change cholesterol or triglyceride plasma levels. However, peritoneal macrophages harvested from E0 mice after consumption of 25 or 50 mg of DTS/day had a lower (p < 0.01) capacity to oxidize LDL (by 45% and by 60%, respectively), and to take up and degrade oxidized LDL (by 41% and 68%, respectively). Consumption of either 25 and 50 mg of DTS/day also reduced the basal level of LDL-associated lipid peroxides by 26% and 62%, respectively (p < 0.01) with a 19%–22% inhibition (p < 0.05) in LDL aggregation. From ex vivo study, it appeared that the aortas of the control 8-week-old apoE−/−/LDLreceptor−/− mice displayed immunoreactivity for TNF-α as well as for TNF p55 and p75 receptors (Fig. 1). At 16 weeks, TNF-α expression in the medium was increased almost five-fold with widespread atherosclerosis and TNF-α immunoreactivity in all plaques and increased TNF-α mRNA levels. Mice with both levels of DTS showed a significant (50 mg > 25 mg) decrease of TNF-α immunoreactivity and mRNA (p < 0.01).
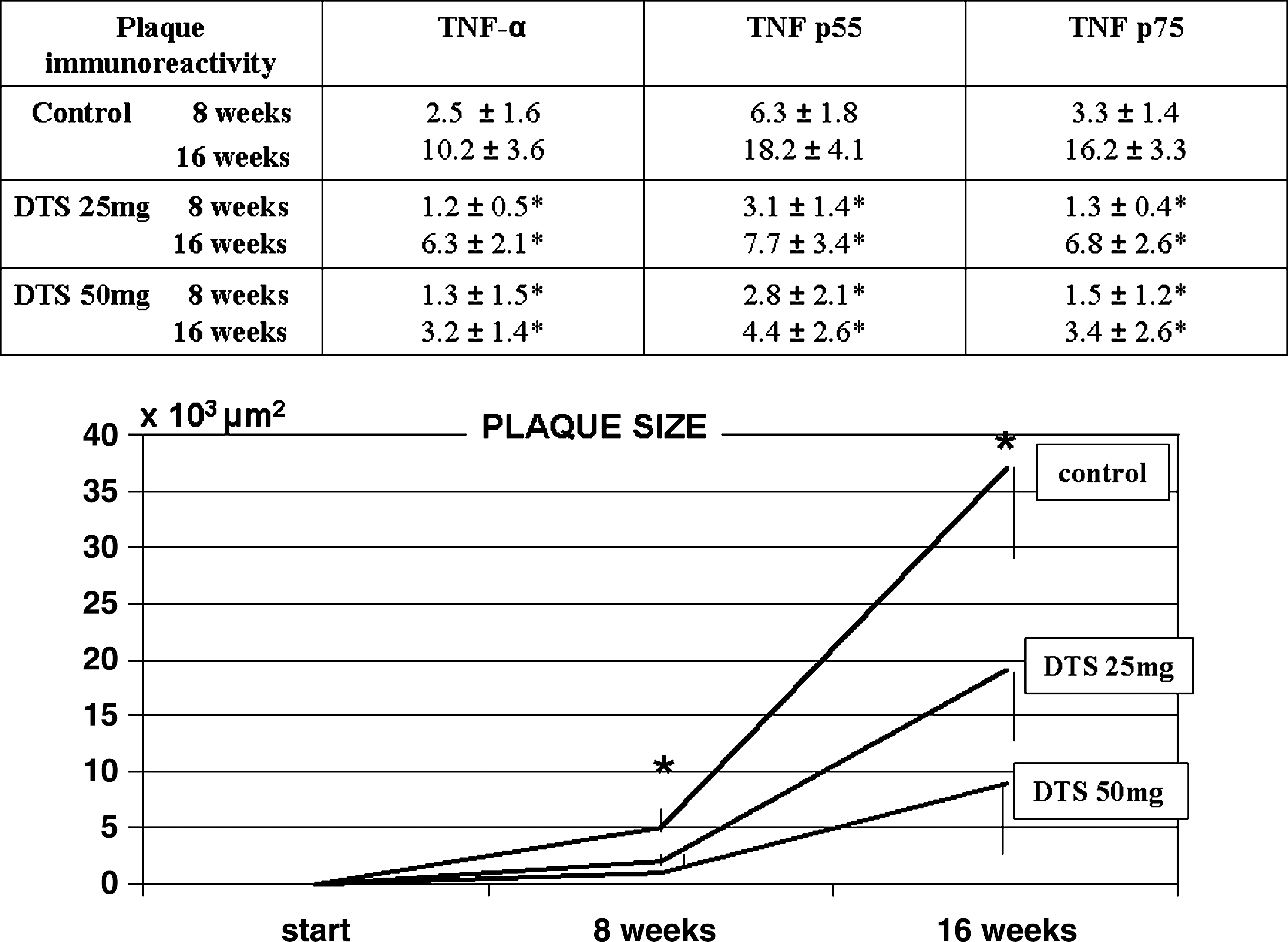
Plaque size, tumor necrosis factor, and tumor necrosis factor receptor expression in apolipoprotein E−/−/low-density lipoprotein receptor−/− (apoE−/−/LDL receptor−/−) mice. Effect of Denshici-to-Chiusei (DTS) supplementation. Values are expressed as mean ± standard deviation (SD) of plaque size (*p < 0.01 vs. both DTS dosages) in experimental animals and immunostaining for tumor necrosis factor-α (TNF-α) and TNF receptors as determined as percentage of plaque area (*p < 0.01 vs. control).
Discussion
The inflammatory nature of atherosclerosis has prompted efforts to prevent development and/or progression of disease by targeting inflammatory mediators. The presence of TNF-α has been demonstrated in human atherosclerotic plaques; it is expressed during intimal thickening and in atherosclerotic plaques and, to a lesser extent, also in the underlying media. Indeed, our study showed that proinflammatory mechanisms take place early in the process. Experimental studies have identified several mechanisms by which TNF-α may promote atherogenesis, including endothelial adhesion molecule expression and activation of macrophages, 11 –13 which also are impaired in their apoptotic capacity. 14 This is confirmed in clinics because there are indications that factors having a direct influence on development and progression of atherosclerotic lesions and circulating TNF-α levels are significantly increased in patients with premature coronary heart disease compared with age-matched healthy controls. 15
Drugs lowering plasma lipids and blood pressure are the fundamental treatments for cardiovascular disease. However, the exact interplay between the plaque-based inflammatory processes of atherogenesis and conventional risk factors need further eludication. 16 Although experimental data have to be interpreted cautiously, the present findings suggest that DTS supplementation is able to counteract some of the key pathogenetic mechanisms effectively, thus offering a promising integrative tool for early clinical application in high-risk patients. Interestingly, quite recently the two major components of DTS have shown effective antiinflammatory and anticomplement actions in an experimental atherogenesis model. 17 –21