
Abstract
Parkinson disease (PD) is a neurodegenerative disorder that provides a useful model for testing cell replacement strategies to rejuvenate the affected dopaminergic neural systems, which have been destroyed by aging and the disease. We first showed that grafts of fetal dopaminergic neurons can reverse parkinsonian motor deficits induced by the toxin, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP), validating the feasibility of cellular repair in a primate nervous system. Subsequent clinical trials in Parkinson patients showed encouraging results, including long-term improvement of neurological signs and reduction of medications in some patients. However, many experienced little therapeutic benefit, and some recipients experienced dyskinesias, suggesting a lack of regulated control of the grafts. We have since attempted to improve cell replacements by placing grafts in their correct anatomical location in the substantia nigra and using strategies such as co-grafting fetal striatal tissue or growth factors into the physiologic striatal targets. Moreover, the use of fetal cells depends on a variable supply of donor material, making it difficult to standardize cell quality and quantity. Therefore, we have also explored possibilities of using human neural stem cells (hNSCs) to ameliorate parkinsonism in nonhuman primates with encouraging results. hNSCs implanted into the striatum showed a remarkable migratory ability and were found in the substantia nigra, where a small number appeared to differentiate into dopamine neurons. The majority became growth factor–producing glia that could provide beneficial effects on host dopamine neurons. Studies to determine the optimum stage of differentiation from embryonic stem cells and to derive useful cells from somatic cell sources are in progress.
Introduction
PD has been treated pharmacologically with the amino acid precursor of dopamine,
Efforts have been made to develop "cell replacements" for the missing and damaged dopamine neurons as an alternative that might fully restore function, consistent with strategies for engineered negligible senescence. Although pathological processes and aging might eventually have deleterious effects on replaced cells, they offer the prospect of full restoration of the functional losses and potential rejuvenation. The earliest reported effort to transplant adult neural tissue (in 1890) was completely unsuccessful. 25 Slow progress began with the discovery in 1917 that embryonic tissue could survive grafting into the brain. 26 Neural grafts of primary fetal dopamine neurons and immediate precursors have been tested as an experimental approach for therapy in PD since 1979, when the first encouraging results were reported in dopamine-depleted rodents. 27,28 Our first work in nonhuman primates established an encouraging degree of efficacy in severely debilitated animals, 29 proving that functional brain cell transplantation was not unique to rodents. We used the neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) to induce all of the classic signs of parkinsonism and create a model for study. Animals that received multiple grafts of carefully dissected fetal dopamine neurons demonstrated a remarkable reversal of motor signs. Survival of grafted dopaminergic neurons was robust and was characterized by the presence of extensive neuritic outgrowth into the host striatum and increased dopamine in the vicinity of the grafts. 30 The survival of cryopreserved human fetal neural tissue transplanted into monkeys, with careful respect for the various ethical concerns, paved the way for human studies. 31
Several clinical trials were conducted and some positive effects were seen in reducing the symptoms of the disease (for reviews, see refs. 32 and 33). Importantly, many subjects showed a reduction in
Stem cells offer a unique opportunity to implant cells of known characteristics and in numbers that can be determined based on the severity of the disorder. Neural stem cells also are known to be prolific producers of neuronal growth factors such as glial-derived neurotrophic factor (GDNF), nerve growth factor (NGF), and others that promote survival of neurons in the central nervous system (CNS). 37 Accordingly, they offer another opportunity for cellular repair in addition to cell replacement. The present contribution reviews our recent work in this area and offers suggestions for future trials based on findings in a translational model that is highly relevant to human trials. We briefly describe our efforts to develop strategies for more physiologic circuit restoration of the nigrostriatal system using different combinations of fetal cells and gene therapy, as well as our studies using neural stem cells in nonhuman primates.
Almost all of the preclinical and clinical studies of transplantation for PD have placed the grafts directly into the putamen and/or the caudate, the presumed targets of the dopamine-producing cells of the substantia nigra (SN). This placement was due to the belief that the distances between the structures were too great for grafted cells to bridge, especially in the absence of the various factors that directed such outgrowth during fetal development. Grafts placed into the SN were shown to have functional effects and projections to the striatum at postnatal days 3 and 10, but not by day 20 in the neonatal rat, 38 confirming the belief that placements in the SN would not be effective in adult animals or patients. Mendez and Hong 39 and Mendez et al. 40 have suggested that the double ventral mesencephalon (VM) grafts were producing unknown growth factors and that the presence of the VM grafts in the SN added to the functional benefits, a procedure that has also been taken into the clinic. 41 It was still believed unlikely, however, that any improved results were due to outgrowth of cells from the SN, especially because dopamine precursors were also placed directly into the striatum.
Our group has used fetal lateral ganglionic eminence (anlagen of the striatum) in a series of experiments to determine the potential attractant effect of placements at increasing distances from VM grafts into the SN. Beginning with graft placements in close proximity, it was apparent that the "striatal" grafts attracted outgrowth from the VM tissue. In a series of studies, 42 we showed that neurites from grafted dopaminergic neurons extended 5–7 mm, almost the full distance to the caudal portion of the caudate in the St. Kitts green monkey 43 (see Fig.1A–D).
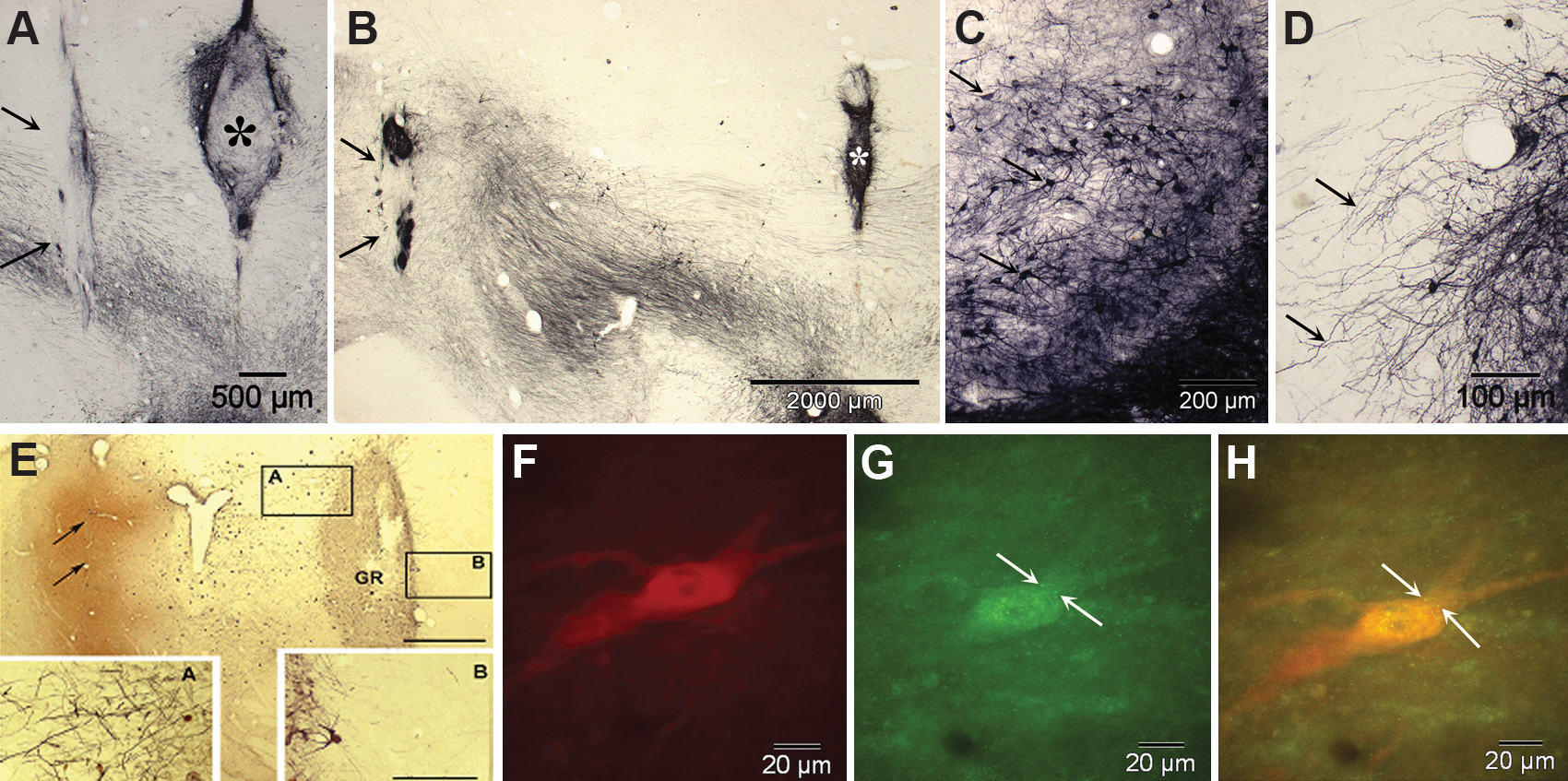
(
In studies of GDNF, which is essential for the survival and outgrowth of developing midbrain dopamine neurons, we confirmed prior studies that GDNF elicits directional neuritic outgrowth of fetal VM neurons grafted into the striatum when GDNF overexpression was induced by an adeno-associated virus (AAV2) vector harboring the GDNF gene. 44 Because GDNF is released into interstitial fluid, we wondered whether this effect might be sufficient to attract neurite outgrowth over the distance from the SN to the rostral caudate.
We then studied adult male monkeys (Chlorocebus sabaeus), with different combinations of VM tissue placed immediately dorsal to or within the SN and either striatal co-grafts or injections of the AAV2/GDNF vector into the target regions. 45 In this experiment, 7–20 days prior to sacrifice, the retrograde tracer Fluoro-Gold (FG) was injected into the identical striatal targets on the ipsilateral and contralateral sides to the SN grafts in 2 animals. Several animals received striatal and nigral co-grafts at increasing distances along the trajectory of the ascending pathway to the striatum from the SN. After 6 months, the animals were killed and the brains were studied using immunohistochemical methods with unbiased stereological counting. FG-labeled tyrosine hydroxylase–positive (marker for dopamine neurons) cells were found in the host SNs as well as labeling small numbers of cells in the grafts, but not in control animals. There was no FG labeling of any of the co-grafts, suggesting that the attractant effect did not extend beyond the striatal co-grafts location substantially into the striatum where the FG injections were made. These results, we believe, support the interpretation that VM grafts placed in the region of the SN have the potential to extend neurites to their physiological targets in the striatum, if growth factors and other conditions are suitable (see Fig. 1E–H).
In parallel to these studies, we have also studied human stem cells, derived from a neuroectoderm-derived structure, the telencephalic ventricular zone of an early second-trimester human cadaver, as an alternative to primary fetal cells (human neural stem cells [hNSCs]). These cells have the potential to be maintained and expanded in culture and to differentiate into all of the cell types of the CNS. Implanting stem/progenitor cells constitutively capable of multiple actions, including differentiation into various cells and secreting cytokines, might allow them to develop in a parkinsonian brain to yield the most appropriate types, numbers, and locations of cells as determined by the host milieu. 37 We studied 27 adult male monkeys (Chlorocebus sabaeus) in several cohorts with different outcome periods up to 8 months after injection into the nigrostriatal system of MPTP-injected and uninjected normal monkeys. 46 –48 Briefly, we found three categories of effects: (1) Differentiation of some NSCs into cells that show classic markers of dopamine neurons; (2) significant migration of hNSCs; and (3) normalizing effects of hNSCs on MPTP-induced abnormalities in the host, as illustrated in Fig. 2. In addition to these potentially normalizing effects, we found a substantial improvement in the parksonian behaviors induced by MPTP in 5 severely affected monkeys, compared with 3 controls. This improvement did not result in a complete recovery from parkinsonism, although it was highly significant statistically as well as functionally, allowing animals to move and feed themselves, which previously they had been unable to do. It is also not clear from these studies which of the mechanisms might be responsible—the homeostatic and normalizing effects observed in the host brain associated with the various types of hNSCs present or the small number of apparently dopamine cells that had differentiated from the hNSCs in the SN.

Over the course of three studies, spontaneous human neural stem cell (hNSC) differentiation (
We and our collaborators are working now to derive therapeutic cells from embryonic stem cells differentiated to the "neural" stage or to a larger percentage of dopamine cells in culture. Several methods have been reported, with functional improvements in rodent models, but the results have been less successful than had been hoped. 49 –53 Several studies in primates nonetheless support the idea that improved methods can be found in the future to fully restore function to the level seen after the best fetal precursor grafts. 50,54 We hope to identify an optimal level of differentiation, in which cells remain migratory and have sufficient supportive factors to sustain them during a more physiologic circuit restoration, such as we saw with AAV2/GDNF injections in the target regions with fetal precursor cells. We now have cultures with about 10% of TH+ cells, which also release dopamine into the medium (see Fig. 3). We are presently investigating such dopamine-enriched cultured cells in MPTP-treated monkeys. We hope soon to study parkinsonian monkeys injected with cells derived from adult fibroblasts (induced pluripotent somatic cells [iPS cells]) that have been differentiated into neural and dopaminergic cells. 55
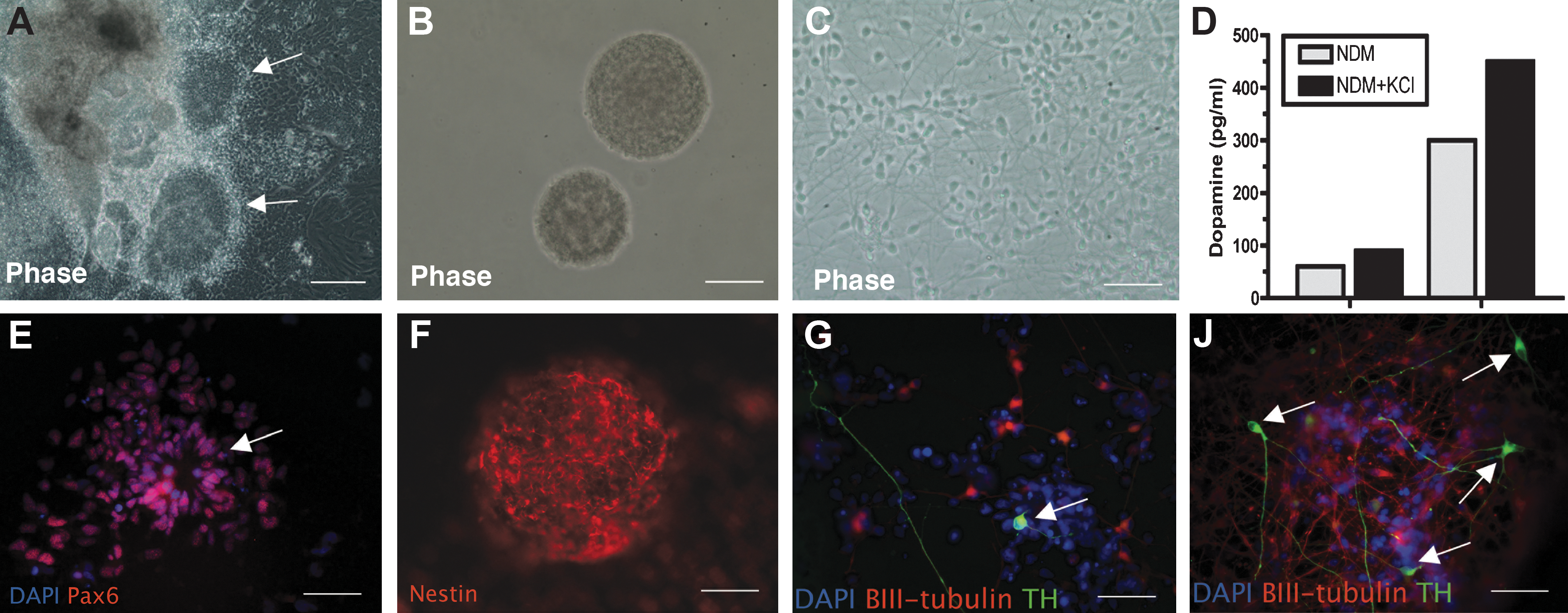
Dopaminergic neurons differentiated from human embryonic stem (hES) cells release dopamine into medium. Briefly, human embryonic stem cells (H1 from Wicell) (passages P44–P54) were cultured in an undifferentiated state on feeder-free and serum-free conditions. They were differentiated into neural precursor cells (NPCs) by culturing them as floating cell aggregates (embryonic bodies) for 3 weeks in a medium supplemented with recombinant human Noggin and basic fibroblast growth factor (bFGF). The NPCs exhibited columnar morphology, formed neural rosettes (arrows,
There is much yet to do before stem cell–derived and reprogrammed cells are ready for rational and controlled clinical trials, including additional studies of toxicity, inappropriate migration, cell overgrowth, and immune rejection. Improved reliability of efficacy and more physiologic circuit reconstruction using tropic and trophic factors are also important goals. Future studies of brain repair and rejuvenation will also benefit from new discoveries of the genetic programs for brain development and improved tools for reprogramming cells, which will make it possible in the future to repair and replace the broken circuits in neurodegenerative diseases.
Footnotes
Acknowledgments
We thank the staff at St. Kitts Biomedical Research Foundation for their invaluable contributions to these studies, especially Dr. Milton C. Whittaker, Dr. Ricaldo Pike, Ernell Nisbett, Clive Wilson I and II, O'Neal Whattley, Xavier Morton, Shervin Liddie, Steve Whittaker, and Samuel Phipps. This research was supported by National Institute of Neurological Disorders and Stroke (NINDS) grants PO1-NS044281 and UO1-NS046028, the Axion Research Foundation, and the Michael J. Fox Foundation for Parkinson's Research. Some of this material is based upon work supported by the State of Connecticut under the Connecticut Stem Cell Research Grants Program. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the State of Connecticut, the Department of Public Health of the State of Connecticut or Connecticut Innovations, Incorporated.
Author Disclosure Statement
The authors declare that no competing financial interests exist.