
Abstract
Bone marrow mesenchymal stem cells (BMSCs) have been reported to possess low immunogenicity and cause immunosuppression of recipients when allografted. They can differentiate into insulin-producing cells and may be a valuable source for islet formation. However, the extremely low differentiating rate of adult BMSCs toward insulin-producing cells and the insufficient insulin secretion of the differentiated BMSCs in vitro prevent their clinical use in diabetes treatment. Little is known about the potential of cell replacement therapy with human BMSCs. Previously, we isolated and identified human first-trimester fetal BMSCs (hfBMSCs). Under a novel four-step induction procedure established in this study, the hfBMSCs effectively differentiated into functional pancreatic islet-like cell clusters that contained 62 ± 14% insulin-producing cells, expressed a broad gene profile related to pancreatic islet β-cell development, and released high levels of insulin (2.245 ± 0.222 pmol/100 clusters per 30 min) and C-peptide (2.200 ± 0.468 pmol/100 clusters per 30 min) in response to 25 mmol/L glucose stimulus in vitro. The pancreatic islet-like cell clusters normalized the blood glucose level of diabetic model mice for at least 9 weeks when xenografted; blood glucose levels in these mice rose abnormally again when the grafts were removed. Examination of the grafts indicated that the transplanted cells survived in recipients and produced human insulin and C-peptide in situ. These results demonstrate that hfBMSCs derived from a human first-trimester abortus can differentiate into pancreatic islet-like cell clusters following an established four-step induction. The insulin-producing clusters present advantages in cell replacement therapy of type 1 diabetic model mice.
Introduction
Our previous studies have verified that BMSCs isolated from human first-trimester abortuses (hfBMSCs) expressed Oct-4, stage-specific embryonic antigen-3 (SSEA3), and SSEA4, showed a diploid karyotype, and formed no tumors in nude mice. Their population doubling times were 34, 36, and 40 h at passages 6, 12, and 24, respectively. 32 In addition, hfBMSCs can differentiate into mesodermal cell types and possess prodigious transdifferentiation potentials. 33 Here, a novel four-step induction procedure was established to induce hfBMSCs into insulin-producing cell clusters similar to the islets of Langerhans that expressed remarkable levels of insulin and C-peptide under glucose stimulation and successfully cured diabetes in nude mouse models when xenografted.
Materials and Methods
Preparation of hfBMSCs
In our previous study, five human abortuses at the age of 10–12 weeks, with permission from the patients, hospital, and the Ethics Committee of Northwest A & F University, were obtained from a local hospital and used to isolate hfBMSCs. 32 Briefly, the limb long bones of five human abortuses were taken out on a germ-free bench and washed with phosphate-buffered saline (PBS), 100 IU/mL penicillin, and 100 mg/mL streptomycin in one 9-cm Petri dish. The two cartilage ends of the bones were cut, and the medullary cavity was opened with scissors lengthwise followed by a calm swirl with minimum essential medium alpha (α-MEM, Gibco, Billings, MT), 100 IU/mL penicillin, and 100 mg/mL streptomycin. Bone bits were removed, and the cell suspension was filtered through a 200-mesh nylon net. The filtrate was centrifuged for 5 min at 1,500 rpm, and the supernatant was discarded. The deposited marrow cells were cultured in one 9-cm plastic culture dish with α-MEM, 10% fetal calf serum (FCS; Stemcell Technologies Inc., Vancouver, Canada), 100 IU/mL penicillin, 100 mg/mL streptomycin, and 0.1mmol/L β-mercaptoethanol (Sigma, Loveland, CO). The culture medium was changed every 2 days, and blood cells were gradually removed by the medium changes. When they grew to monolayer and 80% confluency, fusiform cells were digested with 2.5 g/L trypsin (Gibco) and 0.4 g/L edetic acid (Invitrogen, Carlsbad, CA) and passaged with the same medium. Isolated cells of passage 3 were identified with flow cytometer (Beckman Coulter Inc, Fullerton, CA) for antigen markers of the putative BMSCs, CD29, CD44, CD71, CD105, and CD166, and those of hematopoietic stem cells, CD11a, CD14, CD34, and CD45 (all from Beckman Coulter Inc). 32 Then the isolated hfBMSCs were cultured to passage 6 and cryopreserved in liquid nitrogen.
In vitro induction of hfBMSCs toward insulin-producing cells
Passage 6 of the pooled hfBMSCs was thawed, proliferated to passage 8 in α-MEM, 20% FCS, and 0.1 mmol/L β-mercaptoethanol, and induced using a four-step procedure designed according to the characteristics of different inducers and the induced differentiation of ES cells, 5 –11 pancreatic progenitor/stem cells, 13 –16 hepatic stem cells, 17 –19 neural progenitor cells, 20 and BMSCs 26 –31 into insulin-producing cells. In step 1, the hfBMSC population was adapted to high-glucose environment through a 6-day culture in Dulbecco modified Eagle medium–high glucose (DMEM-HG; containing 25 mmol/L glucose, HyClone, Logan, UT), 20% FCS, and 0.1 mmol/L β-mercaptoethanol. The medium was changed every 2 days. In step 2, the adapted hfBMSCs were induced for 6 days to produce nestin-positive cells in DMEM-HG, 10 ng/mL basic fibroblast growth factor (bFGF, Invitrogen), 10 ng/mL epidermal growth factor (EGF; Chemicon, Temecula, CA), 2% B27 supplement (Stemcell Technologies Inc.), 0.5% bovine serum albumin (BSA; Invitrogen), and 0.1 mmol/L β-mercaptoethanol, with medium changes every 2 days. Step 3 was for the differentiation of nestin-positive cells into insulin-producing cells via a 6-day induction in DMEM-HG, 10 ng/mL EGF, 10 ng/mL hepatocyte growth factor (HGF; Sigma), 10 mmol/L nicotinamide (Sigma), 2% B27, 0.5% BSA, and 0.1 mmol/L β-mercaptoethanol, with medium changes every 2 days. Step 4 was for the maturation of the expected insulin-producing cells via a 4-day culture with DMEM–low glucose (DMEM-LG containing 5.6 mmol/L glucose, HyClone), 10 ng/mL EGF, 10 nmol/L exendin-4 (Sigma), 10 ng/mL betacellulin (Sigma), 25 μmol/L zinc acetate (Sigma), 2% B27, 0.5% BSA, and 0.1 mmol/L β-mercaptoethanol, with medium changes every 2 days. In the four-step protocol, 10 batches (n = 10) were designed with nine replications each, and each replication covers one dish containing over 1 × 106 hfBMSCs.
After the four-step induction, 10 clusters from each of the five batches (n = 50) were photographed, and their diameters were measured under an inverted microscopy (Leica, Wetzlar, Germany). To separate cell clusters from single cells, both in adherence, the cultures were scraped with a plastic cell slicker, collected in a centrifuge tube, and allowed to stand for 10 min at 37°C. Then the supernatant was transferred to another tube to obtain single cells by centrifugation, while the cell clusters in sediment from standing were digested with 2.5 g/L trypsin containing 0.4 g/L EDTA at 37°C for about 2 min. The single cells from the supernatant after that standing and from the digested clusters were counted, the average cell number of clusters was calculated from 10 clusters of each of five batches (n = 50), and the rate of cell clusters formation in this research was defined as the proportion of the cell number of clusters to the total cell number of the adhered culture in six batches with one dish each (n = 6).
Ultrastructural observation of transdifferentiated cells in islet-like clusters
After scraping and standing, some clusters in the sediment were centrifuged for 10 min at room temperature, fixed in 0.1% glutaraldehyde/2% formaldehyde in 0.1 mol/L cacodylate buffer, transferred to 0.1 mol/L cacodylate buffer, and embedded. Samples were sectioned ultrathinly, observed, and photographed under a transmission electron microscopy (JEM-100SX, Japan).
Fluorescent quantitative RT-PCR assay
To clarify the in vitro expression levels of nestin, Pdx1 (pancreatic and duodenal homeobox-1), Ngn3 (neurogenin-3), Pax4 (paired box gene-4), NeuroD1 (neurogenic differentiation-1), Nkx2.2, Nkx6.1, PCSK1 (proprotein convertase subtilisin/kexin type 1), insulin, glucagon, SST (somatostatin), and PP (pancreatic polypeptide) of the hfBMSCs after four-step induction, fluorescent quantitative RT-PCR was conducted, with human fetal pancreatic tissue used as a positive control and noninduced hfBMSCs as negative control. Total RNA of the hfBMSCs after four-step induction and those of the positive and the negative controls were each reverse-transcribed into cDNA with PrimerScript RT reagent kit (TaKaRa) according to manufacturer's manual. The forward and reverse primers for 12 target genes were designed using Primer Express 2.0 according to relevant genes obtained from GenBank. The expression levels of all the genes were analyzed with a fluorescent quantitative PCR instrument (ABI 7500 FAST, Applied Biosystems, USA), in which a 20-μL PCR system, i.e., 10 μL of SYBR Green Realtime PCR Master Mix (TaKaRa, 2 ×), 0.4 μL of forward and reverse primers, 1.0 μL of cDNA templates, 0.4 μL of ROX Reference Dye II (50 ×), and 7.8 μL of dH2O, was adopted. The sequences of primer pair, product sizes, and amplification conditions of 18S gene and target genes are listed in Table 1. Each gene was tested with three independent replications (n = 3).
SST, Somatostatin; PP, pancreatic polypeptide.
Immunofluorescence assay
Cell samples following every induction step were fixed in 4% paraformaldehyde for 20–30 min, washed three times with PBS, and stained immunofluorescently with primary antibodies including mouse anti-human nestin, Ngn3, NeuroD1, PCSK1, glucagon (Abcam, Cambridge, MA), and insulin (Invitrogen), rabbit anti-human Pdx1, MafA (v-maf musculoaponeurotic fibrosarcoma oncogene homolog A), C-peptide (Abcam), SST, and PP antibodies (Invitrogen), and then with secondary fluorescent antibodies, including fluorescein isothiocyanate (FITC)-conjugated donkey anti-mouse immunoglobulin G (IgG) and Alexa Fluor 594–conjugated goat anti-rabbit IgG (Invitrogen). Images were captured with an inverted microscope. Omission of primary antibodies served as a negative control and resulted in no detectable staining. To investigate the rate of insulin-producing cells and other endocrine cells in a cell cluster after induction, cell clusters from 10 batches of hfBMSCs following induction (n = 10) were, respectively, digested, subcultured for 24 h, fixed, and stained immunofluorescently with PCSK1, glucagon, C-peptide, SST and PP antibodies, and relevant IgGs conjugated with fluorophor. Ten non-overlapping visual fields were randomly chosen from each staining of the test and control samples under microscopy, and representative stained and nonstained cells were observed and photographed using an inverted microscope. The proportion of cells immunoreactive to particular antigens was quantified by counting 100 cells per field.
Assay of insulin and C-peptide secretion and intracellular content of proinsulin in vitro
In test groups, islet-like cell clusters from scraped cultures after four-step induction were transferred into 12-well culture plates, 100 clusters per well, and precultured in DMEM-LG, 10 ng/mL EGF, 2% B27, 0.5% BSA, and 0.1 mmol/L β-mercaptoethanol in for 24 h, washed three times with PBS, and stimulated with 1 mL of either 5, 10, or 25 mmol/L glucose in PBS solution containing 1% BSA, with three replication wells for each glucose level and 1 mL of medium per well. In parallel, 100 fetal pancreatic islets isolated from the pancreata of another three human abortuses at the age of 16–20 weeks (positive control) and 1.5 × 104 hfBMSCs at passage 8 (equivalent 100 islet-like clusters) without induction (negative control) were treated in the same way with all three glucose levels as in test group. Fetal pancreatic islets were isolated according to a method reported by Kover et al. 34 Briefly, pancreata from human abortuses were minced and digested with collagenase for 14 min, and pancreatic islets were selected with a filter screen and checked with dithizone staining. The supernatants from all treatments were collected after 30 min of stimulation, and insulin and C-peptide were measured via radioimmunoassay (RIA) with a human insulin-specific RIA kit and a human C-peptide RIA kit (LINCO Research Inc., St. Charles, MO) and a liquid scintillation counter (LS6000, Beckman Co., USA). Either in test groups or in two control groups, 10 batches (n = 10) of experiments, each with three replications, were designed for every glucose level treatment. After four-step induction, 100 pancreatic islet-like clusters were precultured for 24 h followed by washing three times, just as those in test groups before glucose stimulation. After removing the supernatants of the test and control groups, the cells in all treatments were washed with PBS and intracellular proinsulin was extracted with lysis buffer and determined with the human proinsulin RIA kit (LINCO Research Inc.).
Preparation of type 1 diabetic model mice
These experiments were approved by the Institutional Animal Care and Use Committee of Northwest A&F University. A total of 30 adult male nude mice (BALB/c nu/nu) were provided by Experiment Animal Centre of The Fourth Military Medical University (Xi'an, China), each with normal blood glucose levels. Streptozocin (STZ; Sigma) was administered daily by intraperitoneal injection at 50 mg/kg for 5 consecutive days to selectively destroy the pancreatic β-cells. At 72 h after the last STZ injection, fasting blood glucose levels were determined using a standard blood glucose meter (Sure StepTM Plus, LifeScan Inc., Milpitas, CA) once a day for 3 successive days. Only mice retaining higher than 18 mmol/L fasting blood glucose levels over successive 3 days were defined as diabetic model animals; 25 eligible diabetic model mice successfully prepared in this study.
Transplantation of CM-DiI-labeled islet-like cultures and examination of blood glucose, body weight, and immunohistochemistry in recipient model mice
Cultures of hfBMSCs with and without four-step induction were incubated individually in a working solution of Cell Tracker™ CM-DiI (Molecular Probes, Invitrogen) for 5 min at 37°C and for another 15 min at 4°C according to the manufacturer's instructions, followed by a wash with PBS and a check under a fluorescence microscope. In the test group, 600 CM-DiI–labeled and induced cell clusters (about 9 × 104 cells) were transplanted into the right testes of 10 diabetic model mice (n = 10). In parallel as the cell control group, the CM-DiI–labeled but noninduced 9 × 104 hfBMSCs were transplanted into the right testes of another 10 diabetic model mice (n = 10). The same volume of PBS was injected into the right testes of another 5 diabetic model mice as a blank control (n = 5). Blood glucose levels of each treated animal in three groups were measured every 4 days using the standard blood glucose meter and the body weight was measured every 8 days using an electronic balance (Mettler-Toledo Inc., Columbus, OH).
To examine the effects of graft removal on blood glucose fluctuation and the survival and insulin production of the xenografted cells in recipients, the right testes of 3 mice in the test group and of 3 mice in the cell control group, both randomly selected, were removed 30 days posttransplantation; their blood glucose fluctuations and body weights were measured similarly. Histological sections of all testicular grafts were dewaxed and stained with mouse monoclonal antibodies (Abcam) against human insulin or human C-peptide, and then with FITC-conjugated donkey anti-mouse IgG. They were examined using a laser scanning confocal microscopy (Carl Zeiss, Oberkochen, Germany).
To evaluate the glucose clearance effect of the transplanted pancreatic islet-like cell clusters, the intraperitoneal glucose tolerance (IPGT) test was performed on 5 test mice after 12 days of glucose level normalization following the transplantation and 5 control (nondiabetic) mice, each intraperitoneally injected with glucose at 2 mg/g body weight under fasting conditions and tested for blood glucose at 0, 30, 60, 90, 120, and 150 min after the injection. 5
Data processing
Data were subjected to an independent-samples t-test (two-tailed, type 2) using SPSS 15.0, and all values are presented as means ± standard deviation (SD). The statistical difference in means was defined at p < 0.05.
Results
Differentiation of hfBMSCs into pancreatic islet-like cell clusters
In the four-step induced differentiation, some of hfBMSCs died and floated in the culture medium, whereas the survivals adhered, proliferated, and differentiated. They changed in shape from long to short spindles following step 1; the short spindle-shaped cells gradually congregated and formed cell clusters during step 2 (Fig. 1a–d), and the cell clusters gradually became globe clusters during step 3 (Fig. 1e–h). The globe clusters gradually became compact during step 4 (Fig. 1i), and were similar to the islets of Langerhans after the end of the induction procedure.
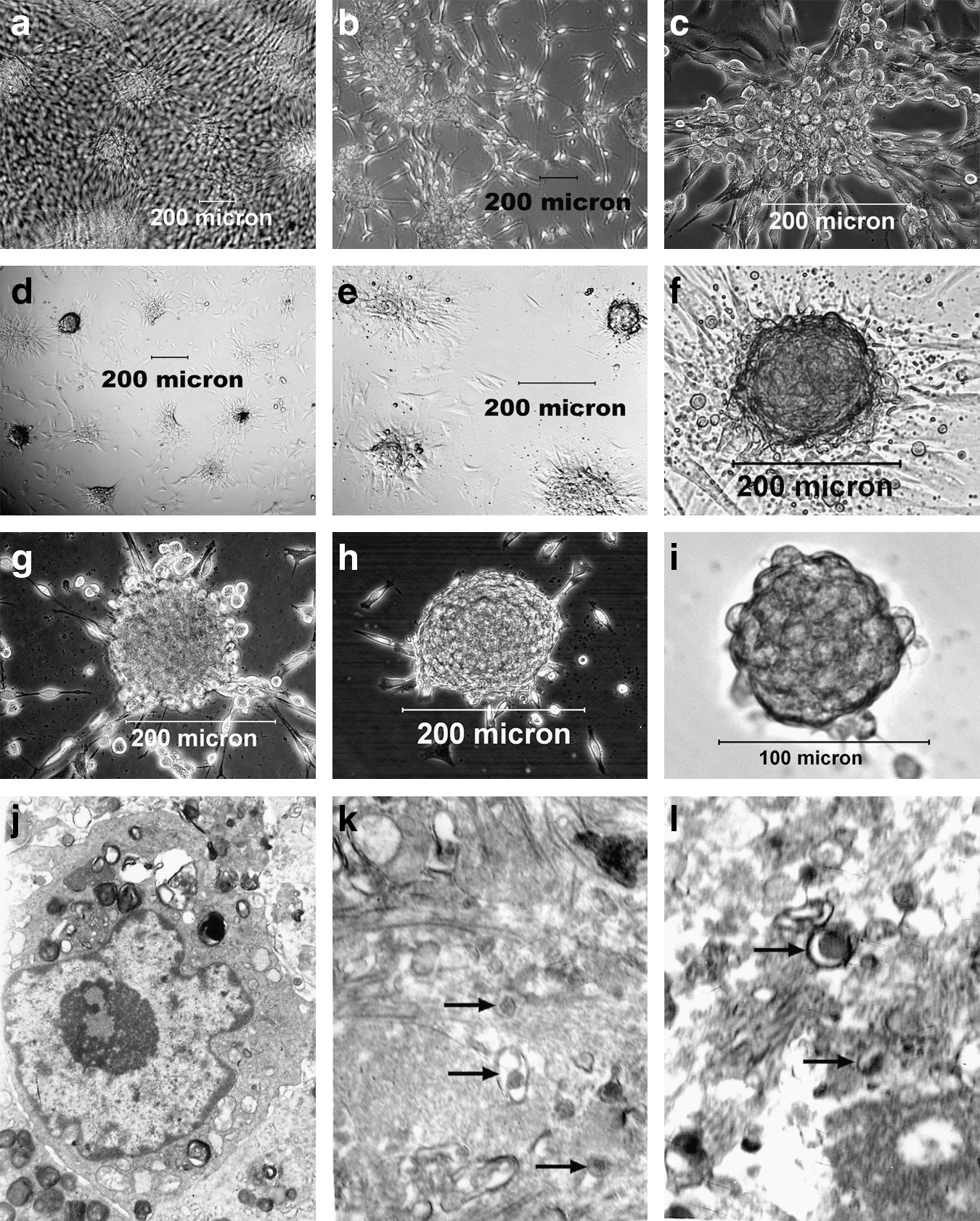
Formation of pancreatic islet-like cell clusters during four-step induction. Pancreatic islet-like cell clusters initially formed after step 2 (
According to counting and measuring methods, the formation rate of the islet-like cell clusters was 50.3 ± 9.4% (n = 6), with a cluster diameter of 125.84 ± 53.45 μm (n = 50) and 148.6 ± 63.4 cells in each cluster (n = 50) on average. Under transmission electron microscopy, a large number of endocrine cells (Fig. 1j) and secretory granules in cytoplasma were observed in the sections of the islet-like clusters (Fig. 1k), which were all histologically similar to those in fetal pancreatic islets in the positive control (Fig. 1l).
Fluorescent quantitative RT-PCR
Fluorescent quantitative RT-PCR of 12 genes showed that, compared to fetal pancreatic tissue, the cells after four-step induction expressed significantly more Ngn3 and Pax4 (147.12 ± 33.27 vs. 64.79 ± 13.19; 138.16 ± 33.20 vs. 61.18 ±8.63, both at p < 0.01, n = 3), more nestin (0.32 ± 0.05 vs. 0.24 ± 0.02, p < 0.05, n = 3), but significantly less Pdx1, insulin, SST and PP (78.63 ± 2.18 vs. 368.90 ± 38.08; 68.27 ± 30.85 vs. 47,538.89 ± 20,746.25; 31.71 ± 7.50 vs. 18,623.31 ± 6,955.19; 7.33 ± 4.00 vs. 1,362.48 ± 237.46, all at p < 0.01, n = 3) and less NeuroD1, PCSK1, and glucagon (112.21 ± 62.12 vs. 776.91 ± 517.69; 28.52 ± 5.81 vs. 19.62 ± 0.59; 57.45 ± 36.57 vs. 23,544.61 ± 18,905.10, all at p < 0.05, n = 3). However, the expression of Nkx2.2 and Nkx6.1 is not statistically different between the induced cells and the positive control (both at p > 0.05, n = 3). The expression profiles of 12 genes are showed in Fig. 2.

Fluorescent quantitative RT-PCR analysis of differentiated human fetal bone marrow mesenchymal stem cells (fBMSCs). After four-step induction, the hfBMSCs expressed high-levels of the specific regulation genes for neogenesis and development of pancreatic β-cells, including Pdx1, Ngn3, Pax4, NeuroD1, Nkx2.2, Nkx6.1, PCSK1, and of hallmark genes for mature islet cells, including insulin, glucagon, somatostatin (SST), and pancreatic polypeptide (PP). Compared to those of fetal pancreatic islets, hfBMSCs expressed more Ngn3 and Pax4 (p < 0.01, n = 3), less NeuroD1, PCSK1, and glucagon (p < 0.05, n = 3) and much less Pdx1, insulin, SST, and PP (p < 0.01, n = 3) after four-step induction. Each gene was tested with three replications.
Immunofluorescence assay
The immunofluorescence assay indicated that the pancreatic islet-like cell clusters expressed nestin, Pdx1, Ngn3, NeuroD1, MafA, insulin, C-peptide, PCSK1, SST, and PP during the cluster formation (Fig. 3), i.e., nestin was strongly expressed after two-step induction (Fig. 3a–c) while feebly expressed after step 1 and 3 of induction (data not shown). Ngn3, Pdx1, NeuroD1, and MafA were expressed after three-step induction (Fig. 3d–k), and insulin and C-peptide (Fig. 3l–o) strongly expressed after all four-step inductions. In contrast, noninduced hfBMSCs expressed none of the proteins listed above except for nestin (figures not shown). After the digestion of pancreatic islet-like cell clusters, the single cells adhered during 24-h subculture and were fixed and stained for PCSK1, PP, glucagon, SST, and C-peptide (Fig. 3p–z). The positive cell percentages for C-peptide, glucagon, SST, and PP were 62 ± 14%, 3 ± 1.4%, 4 ± 3.2%, and 2 ± 2.3%, respectively (n = 10).
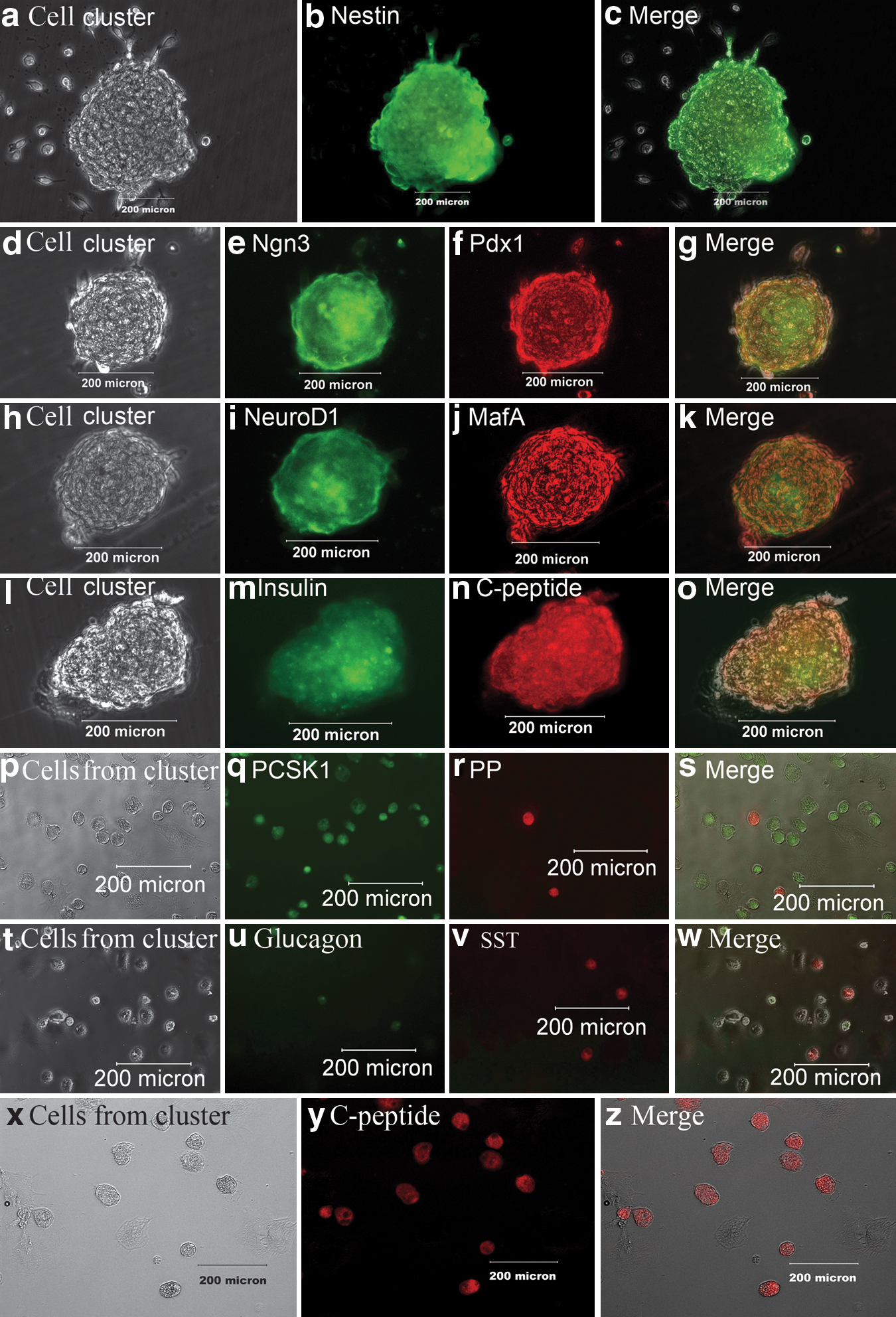
Immunofluorescence assay of pancreatic islet-like clusters during the four-step induction. Nestin was strongly positive in a cell cluster (gray scale in
Levels of insulin and C-peptide secretion and intracellular proinsulin in vitro
A total of 100 pancreatic islet-like cell clusters following four-step induction (test), 100 human fetal pancreatic islets (positive control), and 1.5 × 104 noninduced hfBMSCs (negative control) were all stimulated with glucose at different levels. The results (Fig. 4a) indicated that: (1) The pancreatic islet-like cell clusters formed following four-step induction released insulin and C-peptide at significantly higher levels than those of the negative control (p < 0.01, n = 10). (2) The insulin and C-peptide releases of pancreatic islet-like clusters in response to glucose challenge occurred in a concentration-dependent manner because there was significant difference of each release among different glucose levels (p < 0.01, n = 10). (3) The release patterns of insulin and C-peptide by the pancreatic islet-like clusters were similar to those by human fetal pancreatic islets, but the islet-like clusters secreted significantly less target proteins than those of the human fetal pancreatic islets (p > 0.01, n = 10). The 100 pancreatic islet-like cell clusters contained 5.47 ± 1.54 pmol proinsulin (Fig. 4b), significantly less than that in 100 human fetal pancreatic islets (p < 0.01, n = 10).
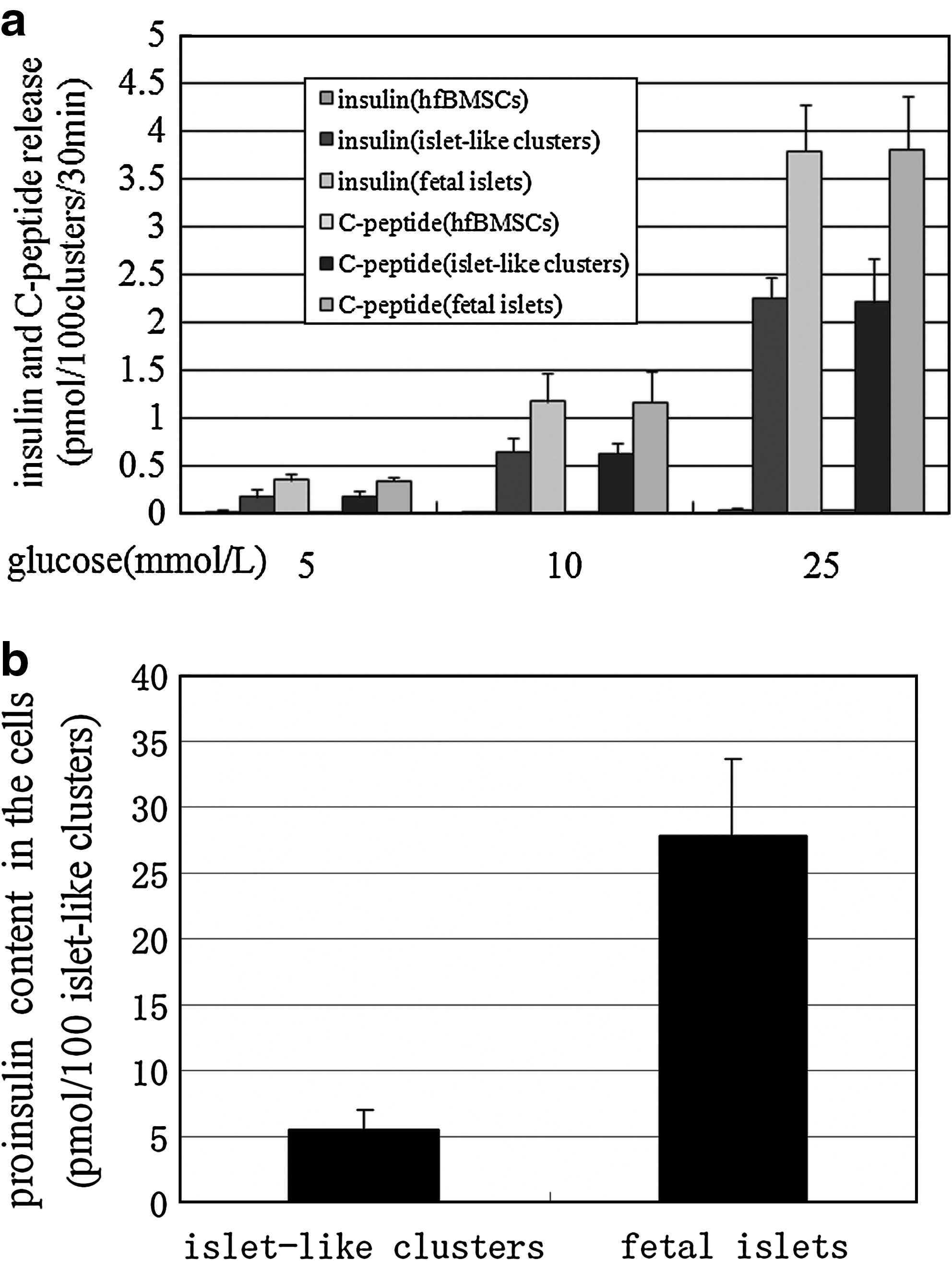
Insulin and C-peptide release and intracellular proinsulin content of islet-like cell clusters. (
Transplantation effects of induced cell clusters in diabetic mice
After STZ injection, 25 of 30 (83%) of the nude mice became eligible diabetic models. Following transplantation with pancreatic islet-like cell clusters, the blood glucose levels of 10 diabetic mice in test group sank to normal levels within 2 weeks. The blood glucose levels of 3 mice in test group, whose testes were removed 30 days after transplantation, rose again, and all 3 died within another 30 days. The remaining 7 mice in test group without the removal of the xenograft maintained normal blood glucose values for at least 9 weeks (Fig. 5a) and gained body weight slightly (Fig. 5b). In cell control group, 10 diabetic mice maintained high blood glucose levels after hfBMSCs transplantation (>18 mmol/L), as did the 3 mice after the removal of their testes 30 days posttransplantation. All 10 diabetic mice in cell control group continuously lost their body weight and died within 60 days after hfBMSCs transplantation. All 5 diabetic mice in the blank control group were hyperglycemic following PBS injection, lost their body weight continuously, and died within 40 days postinjection.
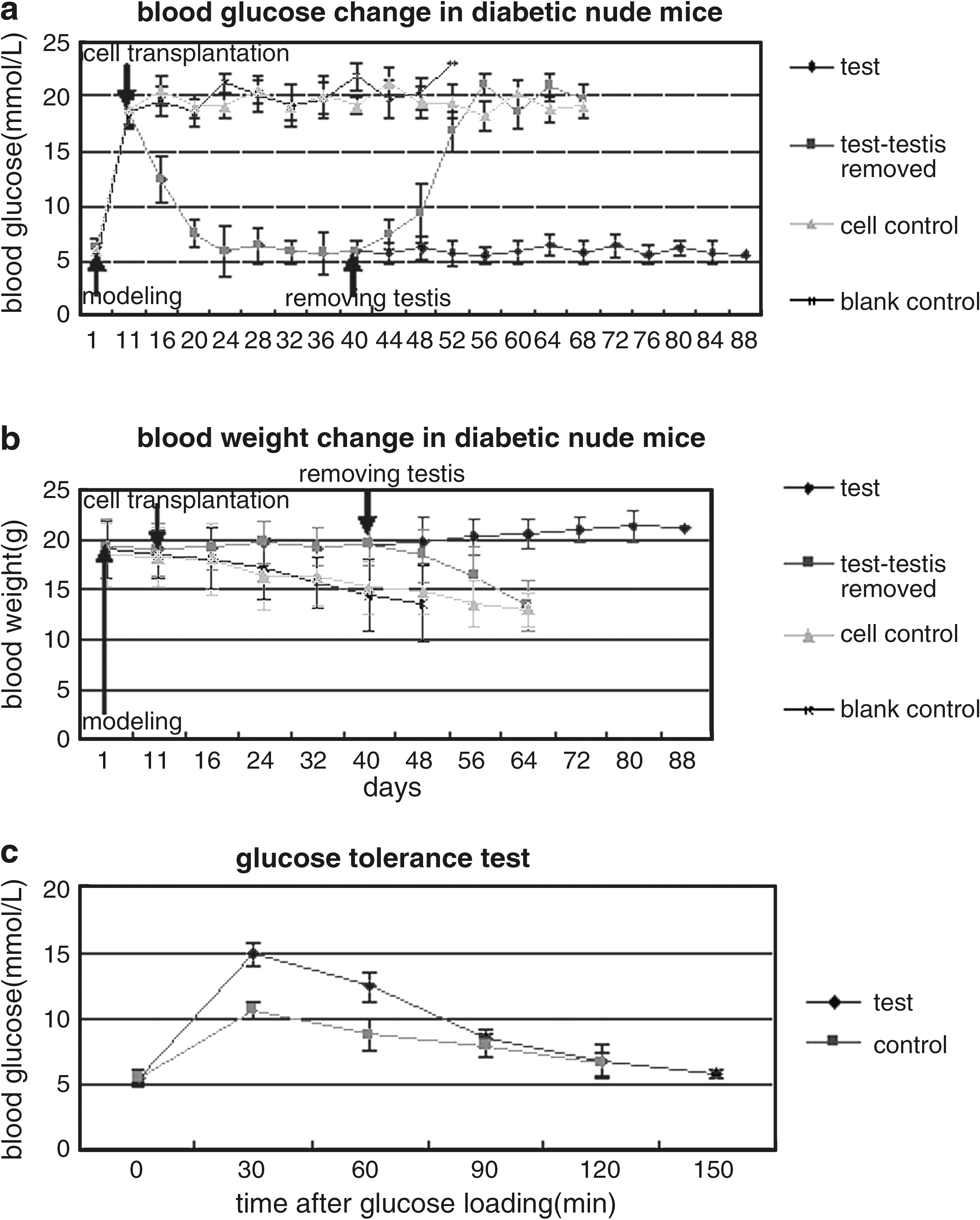
Effects of xenograft with induced cell clusters on the rehabilitation of streptozocin (STZ)-induced diabetic mice. The blood glucose levels of 10 diabetic mice in test group fell to normal within 2 weeks following the transplantation with induced islet-like cell clusters, but 3 out of the 10 mice regained hyperglycemia when their xenograft removed 30 days posttransplantation and all died within 30 days after the removal. The remaining 7 mice in the test group without the removal of xenograft maintained normal levels of serum glucose for at least 9 weeks (
The results of the IPGT test (Fig. 5c) indicated that the induced pancreatic islet-like cell clusters had a normal rate of glucose clearance in vivo, but were not as effective as native pancreatic β-cells in terms of restoring normoglycemia.
A direct check of dewaxed testis sections with laser scanning confocal microscopy showed that the CM-DiI–labeled cells were distributed mainly in the interstitial tissue between seminiferous tubules in graft (red, Fig. 6a–c). Immunofluorescent examination of the dewaxed sections indicated that the CM-DiI–labeled red cells in interstitial tissue were both positive to insulin (green, Fig. 6d–f ) and to C-peptide (green, Fig. 6g–i) in the test group instead of the cell control group (Fig. 6j–l), showing that insulin and C-peptide were produced by induced pancreatic islet-like cell clusters instead of by noninduced hfBMSCs.

Immunofluorescence examination of dewaxed sections of the grafts. Red CM-DiI fluorophore was visualized only in the interstitial tissue between the seminiferous tubules in a graft section (
Discussion
Induction strategies and principle
Previous research results have indicated that the development of pancreatic endocrine cells is similar to that of neurocytes, 35,36 and nestin is commonly expressed by precursors of neurocytes and pancreatic endocrine cells. 8,14 –16,37,38 Recent research indicates that nestin plays pivotal roles as an intermediate regulator governing both stemness and differentiation of stem cells in the process of their differentiation into insulin-secreting cells. Inhibition of nestin expression in adult rat pancreatic ductal stem cells caused a low expression of pancreatic transcription factors and islet hormones, resulting in poor β-cell development and insulin secretion. 39 Therefore, the strategy in this study was to produce nestin-positive multipotential cell clusters along with those in which there was induced differentiation into functional pancreatic islet-like cells under a four-step induction. Step 1 is an adaptive culture in a high-glucose environment. In step 2, the hfBMSCs were proliferated into nestin-positive cells in a serum-free induction cocktail where bFGF cooperated with EGF to promote the proliferation of nestin-positive cells 40 –43 and B27 served as a nutrient additive to promote the growth of the nestin-positive cells. Step 3 aimed to induce the nestin-positive cells to differentiate into insulin-producing cells with the supplementation of EGF, HGF, nicotinamide, and B27 in the culture medium, where EGF facilitated the expansion of pancreatic progenitors. 11 Nicotinamide and B27 promoted nestin-positive cells to differentiate into insulin-producing cells, 44,45 and HGF regulated the pancreatic islet cells to proliferate and differentiate. 46 Step 4 was to promote the maturation of pancreatic endocrine cells and proinsulin synthesis and to enhance the sensitivity of β-cells to glucose in presence of EGF, exendin-4, betacellulin, and zinc acetate, whereas EGF, exendin-4, and betacellulin promoted growth and maturation of β-cells, 47 –51 and zinc ion is essential to β-cells during maturation and insulin synthesis. This four-step induction strategy was well substantiated by all experimental results. Undeniably, some researchers have induced human ES cells into insulin- and glucagon-secreting cells without specifically considering the enrichment of nestin-positive stem cells, 52,53 and nestin-positive cells are believed to be insufficient for islet formation in some cases. 52 This may be related to differences in stem cell genotypes and the heterogeneity of nestin-positive cells, 54 presenting topics for further research on induction of insulin-producing cells.
Differentiating effectiveness of hfBMSCs into insulin-producing cells
Immunofluorescent staining showed that mouse BMSCs can transdifferentiate toward insulin-producing cells in vitro at the rate of 10–20%, and the induced 106 cells released 4.7 ± 0.17 ng/mL insulin after a 2-h stimulus with high-glucose medium. 26 Similarly, rat BMSCs were induced into insulin-positive cells at a rate of 19.8%, and 106 induced cells released only 2.0 ng of insulin under high-glucose medium stimulation. 28 Under the four-step induction system of this study, the hfBMSCs formed cell clusters, with a diameter of 125.84 ± 53.45 μm (n = 50), morphologically similar to the islets of Langerhans (Fig. 1). The ratio of clustering cells amounted to 50.3 ± 9.4% out of all surviving cells (n = 6), and each cluster contained 148.6 ± 63.4 cells (n = 50). The cell clusters comprised mostly endocrine cells, including 62 ±14% insulin-producing cells (n = 10). And similar to cells in fetal pancreatic islet, the cells in clusters contained secretary granules shown under electron microscopy. In addition, the gene expression profile of the induced hfBMSCs accorded with that of human fetal pancreatic islet cells (Fig. 2), and the induced islet-like cell clusters released high level of insulin at 2.245 ± 0.222 pmol/100 clusters/30 min and C-peptide at 2.200 ± 0.468 pmol/100 clusters per 30 min in response to 25 mmol/L glucose stimulus (Fig. 4a), and contained high level of proinsulin at 5.47 ± 1.54 pmol/100 clusters/30 min (Fig. 4b). All of these findings fully illustrate a high induction efficiency of the four-step procedure, an exceptional directed differentiation potential of hfBMSCs into insulin-producing cell lineage under the four-step induction, and the extraordinary insulin secretion capacity of these cells in higher-glucose medium.
In addition, the islet-like cell clusters contained not only insulin-producing cells but also glucagon-producing cells, STT-producing cells, and PP-producing cells as well, implying that the differentiation process of insulin-producing cells is related to those of glucagon-producing cells, SST-producing cells, and PP-producing cells. Their levels of insulin and C-peptide release were less than those of the pancreatic islets from human abortuses (p < 0.01, n = 10); the rate of insulin-producing cells in the islet-like cell clusters, 62 ± 14%, was lower than 70–75% in normal pancreatic islets and insulin-producing cells in the islet-like cell clusters in initial differentiation stage before maturation.
Cell transplantation site
Diabetes can be treated either by orthotopic transplantation or by heterotopic transplantation. In the former, pancreatic islets are transplanted via the portal vein, liver, spleen, omentum majus, and peritoneal cavity, and the insulin produced by the seeded cells is in the liver or can interflow into the liver via the portal vein, according with the physiological metabolic pathway of insulin. Therefore, orthotopic transplantation is mainly practiced in the clinic, but the recovery of the graft is difficult. In the latter, however, it is easy to retrieve and examine the grafts and monitor the serum glucose of the recipient after the removal of graft because the islets are seeded into the testis or underneath the renal capsule; thus, so heterotopic transplantation is the major approach in research field.27,53 Considering the surgical difficulty and the effects of removal of graft on the recipients, the testis is more advantageous than the kidney for transplantation. Therefore, the induced clusters are seeded into the testes of model animals in this study.
Effects of islet-like cell cluster transplantation on the rehabilitation of diabetic nude mice
Although some attempts have been made to differentiate BMSCs toward insulin-producing cells, the results are not ideal. The glucose levels of STZ-induced diabetic rats began decreasing at day 6 after the allograft, with 5 × 106 induced rat BMSCs via the portal vein and kept below 15 mmol/L from day 12 to day16 posttransplantation, and then elevated again after day 20. 28 And the plasma glucose level of STZ-induced diabetic mice reduced markedly after the allograft with insulin-producing cells derived from mouse BMSCs, and the effect continued for at least 4 weeks. 29 Syndromes of STZ-treated diabetic mice were ameliorated by the transplanting insulin-producing cells derived from adult BMSCs. 31
In this study, the hyperglycemia of 10 diabetic nude mice was completely cured by transplanting induced islet-like cell clusters from hfBMSCs, but the blood glucose levels of 3 mice in the test group rose again when the grafts were removed, whereas 7 other mice in the test group maintained normal blood glucose levels for at least 9 weeks (Fig. 5a). This indicated that the transplanted cells were responsible for the recovery from hyperglycemia. This was further confirmed by the co-label of CM-DiI and of immunofluorescent antibodies against insulin and C-peptide in the grafts, demonstrating that the transplanted cells survived in the grafts and produced functional human insulin and C-peptide (Fig. 6). These results demonstrate that hfBMSCs derived from human first-trimester abortuses can differentiate into pancreatic islet-like cell clusters following an established four-step induction and the insulin-producing clusters present advantages in cell replacement therapy of type 1 diabetic model mice.
In comparison with the fetal pancreatic islets, the induced islet-like clusters expressed more Ngn3 and Pax4, the markers of early stage of β-cell development, but expressed less insulin (Fig. 2) of matured β-cells, and secreted relatively lower insulin in vitro (Fig. 4a), indicating that the induced cell clusters differentiated at the initial stage toward the β-cell. And they normalized the blood glucose levels of diabetic model mice 12 days after transplantation (Fig. 5a), implying that the induced cell clusters slowly adapted to the testicular niches of the recipients, proliferated, and/or further differentiated and matured in vivo. After removal of transplanted testes, it took another 2 weeks until the animals completely reversed to diabetics (Fig. 5a), implying that some transplanted cells possibly migrate to other locations and escape the removal of the testis, bringing on the retarded relapse to full diabetes.
Some researchers found the core cell damage in isolated adult islets of Langerhans in the first week after transplantation. 55 The cell damage may be caused by the insufficient revascularization resulting from a larger diameter of the cell mass, averaging 200 μm, which prevents adequate nutrient provision. 56 The islet-like clusters generated in this study are smaller, averaging 125.84 μm, facilitating the supply of oxygen or nutrients by diffusion and so lowering the initial degradation after transplantation. On the other hand, BMSCs have been reported to possess low immunogenicity and cause immunosuppression in recipients when allografted. 22 –25 The hfBMSCs are primordial BMSCs, both expressing hallmarks of mesenchymal stem cells—CD29, CD44, CD105, and CD166. 32 Therefore, it can be supposed that the islet-like clusters derived from hfBMSCs may possess similar properties to BMSCs. The immune elimination of type 1 diabetics will be weaker when they receive the islet-like clusters derived from hfBMSCs than isolated adult islets and insulin-producing cells derived from other adult stem cells.
Expansion and differentiation capacity of the hfBMSCs under four-step induction
In this study, a single islet-like cluster contains about 150 cells. Considering the cluster-forming rate to be around 50% and the cell mortality near 25%, 400 hfBMSCs are needed to form one islet-like cluster under the four-step induction system. Cure of a 20-g diabetic model nude mouse requires 600 islet-like clusters. Considering this body weight, 2.25 × 106 islet-like clusters, or 9 × 108 (2.25 × 106 × 400) hfBMSCs, are needed for the therapeutic transplantation for a 75-kg diabetic patient.
The pooled bone marrow cells of five abortuses in this study propagated to 1 × 106 primary fBMSCs after 2 weeks of culture and expanded to 6.56 × 109 hfBMSCs, more than seven times than 9 × 108, until passage 8, in which the hfBMSCs proliferated threefold after each passage. In other words, the hfBMSCs at passage 8 derived from five abortuses can meet the cell amount needed for therapeutic transplantation of seven 75-kg diabetic patients. Accordingly, the islet-like clusters generated from the hfBMSCs after four-step induction may relieve the problem of insufficient islets donation for transplantation in the regions where abortion is legalized, although we are not advocating abortion for the single purpose of obtaining fetal BMSCs.
Footnotes
Acknowledgment
We thank the Key Program of National Ministry of Education (03160) and the National Natural Science Foundation of China (30671067) for their support.