
Abstract
Pluripotent stem cells offer unique opportunities for curing debilitating diseases. However, further comprehensive research is needed to better understand cell signaling during the differentiation of pluripotent cells into different cell lineages and accordingly to develop clinically applicable protocols. One of the limiting steps for differentiation studies is proper culture and expansion of pluripotent stem cells, which is labor intensive, expensive, and requires a great deal of expertise. This limiting step can be overcome by successful banking and distribution of embryoid bodies (EBs), which are aggregates of pluripotent stem cells and typically the starting point of differentiation protocols. The objective of this study was to investigate the feasibility of EB banking by studying survival and functionality of cryopreserved EBs. To this end, EBs were formed by culturing mouse 129 embryonic stem (ES) cells in the absence of leukemia inhibitory factor (LIF) in hanging drops and then subjected to different cryopreservation protocols. In a series of experiments, we first tested the postthaw survival of EBs as a function of dimethylsulfoxide (DMSO) and extracellular trehalose concentrations and cooling rates. Next, we studied the functionality of cryopreserved EBs by assessing their postthaw attachment, growth, and differentiation into various cell types. Higher (≥5%) DMSO concentrations alone or in combination with trehalose (0.1 M and 0.2 M) yielded good postthaw survival rates of >80%, whereas cooling of EBs at 1°C/min in the presence of 5% DMSO +0.1 M trehalose gave the best attachment and growth rates, with differentiation into cell lineages of three germ layers. Taken together, our results suggest that EBs are tolerant to cryopreservation-associated stresses and retain their differentiation potential after freezing and thawing. Furthermore, our experiments with dissociated EB cells and nondissociated EBs suggest that the extracellular matrix may play a beneficial role in the cryotolerance of EBs. Overall, our data support the feasibility of EB banking, which would facilitate advancement of cell-based therapies.
Introduction
Recently, somatic cells have been reprogrammed into a pluripotent state by deliberately expressing various combinations of transcription factors, such as Oct4, Sox2, Klf4, c-Myc, Nanog, and Lin28. 15 –19 These so-called induced pluripotent stem cells (iPSCs) are similar to ES cells in terms of unlimited self-renewal capacity and differentiation into cell types from three germ layers (i.e., ectoderm, endoderm, and mesoderm). Hence, this reprogramming approach can be used for generation of disease- and patient-specific pluripotent stem cells. In fact, this has already been accomplished by several groups. 20 –24 Taken together, pluripotent stem cells offer enormous potential for translational research and treating diverse diseases. However, further fundamental research is needed to understand signaling pathways controlling differentiation of pluripotent stem cells into different cell lineages and accordingly to develop directed differentiation protocols. The quality and homogeneity of the differentiated cells and related biosafety issues also require comprehensive studies. One of the limiting steps for such research is proper culture and expansion of pluripotent stem cells, which is labor intensive, expensive, and requires a great deal of expertise. This limiting step can be overcome by successful banking and distribution of embryoid bodies (EBs), which are aggregates of pluripotent stem cells and typically the starting point of differentiation protocols. Particularly, the easy availability of consistently produced and characterized high-quality EBs in large quantities would ensure a uniform starting point for various differentiation protocols and facilitate the reproducibility of the outcome. The objective of this study was to investigate the feasibility of EB banking. For this purpose, EBs generated from mouse ES cells were used as models and subjected to different cryopreservation protocols to study their postthaw survival and functionality.
Materials and Methods
Culture of ES cells and EB formation
All cell cultures were carried out at 37°C under a humidified gas atmosphere of 5% CO2 in air. The mouse 129SvJ ES cells (GSI-1, Genome System Inc., St. Louis, MO) were expanded on a feeder cell layer of irradiated mouse primary fetal fibroblasts that were plated on gelatin-coated six-well dishes containing Knockout Dulbecco modified Eagle medium (DMEM; Invitrogen, Carlsbad, CA) supplemented with 20% (vol/vol) fetal bovine serum (FBS; Atlanta Biologicals, Lawrenceville, GA), 1,000 IU/mL recombinant murine leukemia inhibitory factor (LIF; ESGRO, Chemicon), 2 mM
Experimental groups and cryopreservation of EBs
To study the feasibility of EB banking, we used dimethylsulfoxide (DMSO) as a model permeating cryoprotective agent (CPA). In the first set of experiments, postthaw survival of EBs was studied as a function of DMSO concentrations by cryopreserving EBs in the presence of 2.5%, 5%, and 10% DMSO using a fixed cooling rate of 1°C/min, while unfrozen EBs served as controls. The postthaw survival was assessed based on membrane integrity as described later. The second set of experiments was designed to study the postthaw survival as a function of cooling rates (i.e., 1°C/min, 5°C/min, and 10°C/min) and extracellular trehalose concentrations (i.e., 0.0 M, 0.1 M, and 0.2 M), while the DMSO concentration was set to 5% based on the first set of experiments. Thus, the second set of experiments had nine experimental groups and unfrozen controls. In third set of experiments, we challenged intact and dissociated EBs (the latter are also referred as dissociated EB cells in the rest of this manuscript) by subjecting them to a freeze–thaw cycle in the absence of DMSO and trehalose to address the effect of extracellular matrix on the postthaw membrane integrity. The fourth and final set of experiments was carried out to address the functionality (i.e., postthaw attachment, growth and differentiation) of cryopreserved EB using the same conditions (i.e., nine experimental groups plus control) as tested in the second set of experiments.
To load cells with DMSO, the selected EBs with proper size were first rinsed in a relevant CPA solution and then exposed to the same CPA solution at room temperature for 15 min based on our preliminary equilibration experiments with 10% DMSO (see Supplementary Fig. 1, accessible at
Membrane integrity assay
To evaluate the postthaw survival of EBs, we used a membrane integrity assay based on two fluorescence dyes, SYTO-13 (Molecular Probes, Eugene, OR) and ethidium homodimer (EH, Molecular Probes). SYTO-13 permeates through both intact and compromised cell membranes, and thus labels all cells green, whereas EH can permeate only compromised cell membranes and stains nuclei of nonintact cells red. To determine the number of intact and nonintact cells based on this differential labeling, EBs were incubated in DMEM/F-12 medium containing 5 mM of SYTO 13 and 3 mM EH for 15 min and then washed in fresh DMEM/F-12. Subsequently, stained EBs were imaged using an epifluorescence microscope (Zeiss Axioskop 2 plus) and appropriate filter sets. Finally, the number of green- and red-stained cells were counted to calculate the proportion of intact EB cells using the following formula:
Because counting individual EB cells is very labor intensive, we first inspected all stained EBs under an epifluorescence microscope to confirm the uniformity within each group, and then randomly took representative images to calculate the proportion of intact EB cells.
Postthaw attachment, growth, and differentiation of EBs
To examine the postthaw functionality of cryopreserved EBs, we cultured EBs individually on poly-
To encourage the differentiation of EB cells toward neural lineage, we supplemented the differentiation medium with 1 mM all-trans retinoic acid (ATRA; Sigma, St. Louis, MO). 25 To direct their differentiation into hepatic lineage, frozen–thawed EBs were subjected to a sequential differentiation protocol modified from a previously published work. 26 Briefly, frozen–thawed EBs were first cultured in RPMI-1640 medium (CellGro) supplemented with 5% FBS and 50 ng/mL activin A to induce definitive endoderm. After 3 days of suspension culture, the EBs were next plated on collagen (type I) in the presence of RPMI-1640 containing 5% FBS, 10 ng/mL basic fibroblast growth factor (bFGF, Millipore), 10 ng/mL bone morphogenetic protein 4 (BMP4; ProSpec) to promote early hepatic differentiation. After 4 days of culture, the supplementation of the RPMI-1640 medium was changed to 5% FBS and 10 ng/mL hepatocyte growth factor (HGF; ProSpec) for a mid hepatic differentiation phase of 4 days. The final hepatic maturation was accomplished by culturing the cells in RPMI containing 10 ng/mL oncostatin M (ProSpec), 0.1 μM dexamethasone (Sigma), 5 mg/mL insulin (Gibco), 5 mg/mL transferrin (Gibco), and 5 mg/mL selenious acid (Gibco) for 3 days. Differentiated cells were fixed in 4% paraformaldehyde in PBS at room temperature for 20 min and subsequently permeabilized with 0.1% Triton X-100 in PBS for 15 min.
The fixed cells were blocked with a commercial blocking solution (Protein-Free Blocking Buffer, Thermo Scientific) for 60 min and then incubated with a primary antibody (1:200 dilution in PBS) against either β-tubulin III (Chemicon) or albumin (Bethyl Laboratories, Inc.) at room temperature for 60 min to examine the differentiation of EB cells to neural and hepatic lineage, respectively. After three washes in PBS+0.05% Tween 20 for 10 min each, the fixed cells were then treated with an either fluorescein isothiocyanate (FITC)-conjugated or Cy3-conjugated secondary antibody (1:200 dilution in PBS) at room temperature for 60 min against β-tubulin III and albumin primary antibodies, respectively. Finally, the secondary antibody was washed out, and labeled cells were imaged using an epifluorescence microscope (Zeiss Axioskop 2 plus) with relevant filter sets.
Statistical analysis
Experiments in each series were repeated at least three times using EBs from independent batches. Data reported are means±standard error of mean (SEM). In some figures, data are shown as box and whisker plots where proportional box plots specify the 25th, 50th (the median), and 75th percentiles while the ends of the whiskers represent the minimum and maximum. The results were analyzed by analysis of variance (ANOVA) and using GraphPad Prism (GraphPad Software, Inc., San Diego, CA). Before ANOVA, arcsine transformation was performed on proportional data. Differences between the groups were considered statistically significant when the p value was less than 0.05.
Results
Effect of different concentrations of a permeating CPA on postthaw survival of EBs
This set of experiments were designed to investigate the postthaw viability as a function of permeating CPA (i.e., DMSO) concentrations. The rationale was to find the lowest effective concentration of the permeating CPA, and thus to minimize its long-term toxic effect. To this end, three concentrations (2.5%, 5%, and 10%) of DMSO were tested with respect to unfrozen controls. The results of these experiments were summarized in Figs. 1 and 2. All EBs maintained their overall morphology after thawing (Fig. 1A,B). Indeed, the membrane integrity assay revealed that high proportions of EB cells were intact in all groups (Fig. 1C). However, the survival rates in 5% DMSO (89.7%) and 2.5% DMSO (84.2%) groups were significantly lower when compared to that of unfrozen controls (94.5%), while there was no significant difference between the control and 10% DMSO groups regarding the survival rates (94.5% and 91.6%, respectively) (Fig. 2). On the other hand, the survival rates in 10% and 5% DMSO groups were statistically similar and significantly higher than that in 2.5% DMSO group. Overall, these results were encouraging and suggested that cryobanking of EBs can be achieved. On the basis of these results, we selected 5% DMSO for the subsequent experiments with the expectation to further improve the cryoprotection while limiting CPA toxicity.
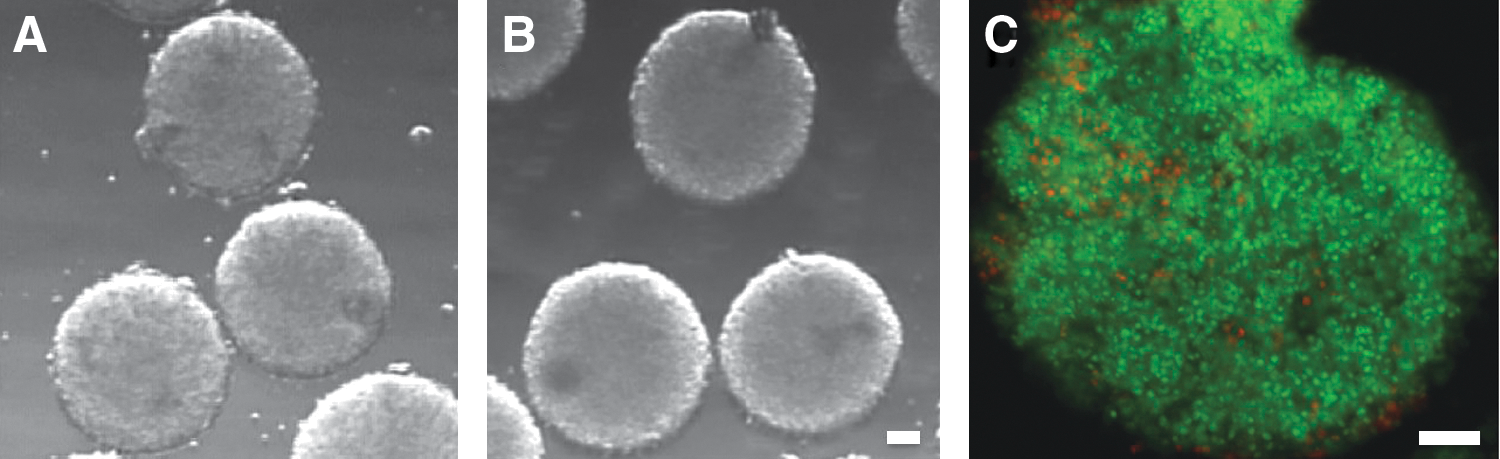
Morphologic appearance and membrane integrity of frozen–thawed embryoid bodies (EBs). Generally, EBs displayed similar morphology before (

Postthaw survival of embryoid bodies (EBs) as a function of dimethylsulfoxide (DMSO) concentrations. The membrane integrity of EBs either remained at 37°C (control) or cooled to −196°C (DMSO groups) was assessed by SYTO-13/EH staining as described in the text. Proportional box plots specify the 25th, 50th (the median), and 75th percentiles while the ends of the whiskers represent the minimum and maximum. Boxes with different letters are significantly different (p<0.05).
Postthaw survival of EBs as a function of cooling rates and trehalose concentrations
Because we and others have shown that extracellular trehalose in combination with a conventional permeating CPA improves the cryopreservation outcome of various cell types and tissues, 27 –30 this set of experiments was conducted to examine whether different cooling rates and extracellular trehalose concentrations can improve the cryosurvival of EBs. To this end, we investigated the postthaw survival of EBs that were cryopreserved in the presence of 0.0 M, 0.1 M, and 0.2 M trehalose using three different cooling rates (1°C/min, 5°C/min, and 10°C/min). On the basis of the first set of experiments, the DMSO concentration was fixed at 5%. The control group consisted of unfrozen EBs. The results of these experiments are shown as a box and whisker plot in Fig. 3. After thawing, all cryopreserved EBs displayed an intact EB morphology.
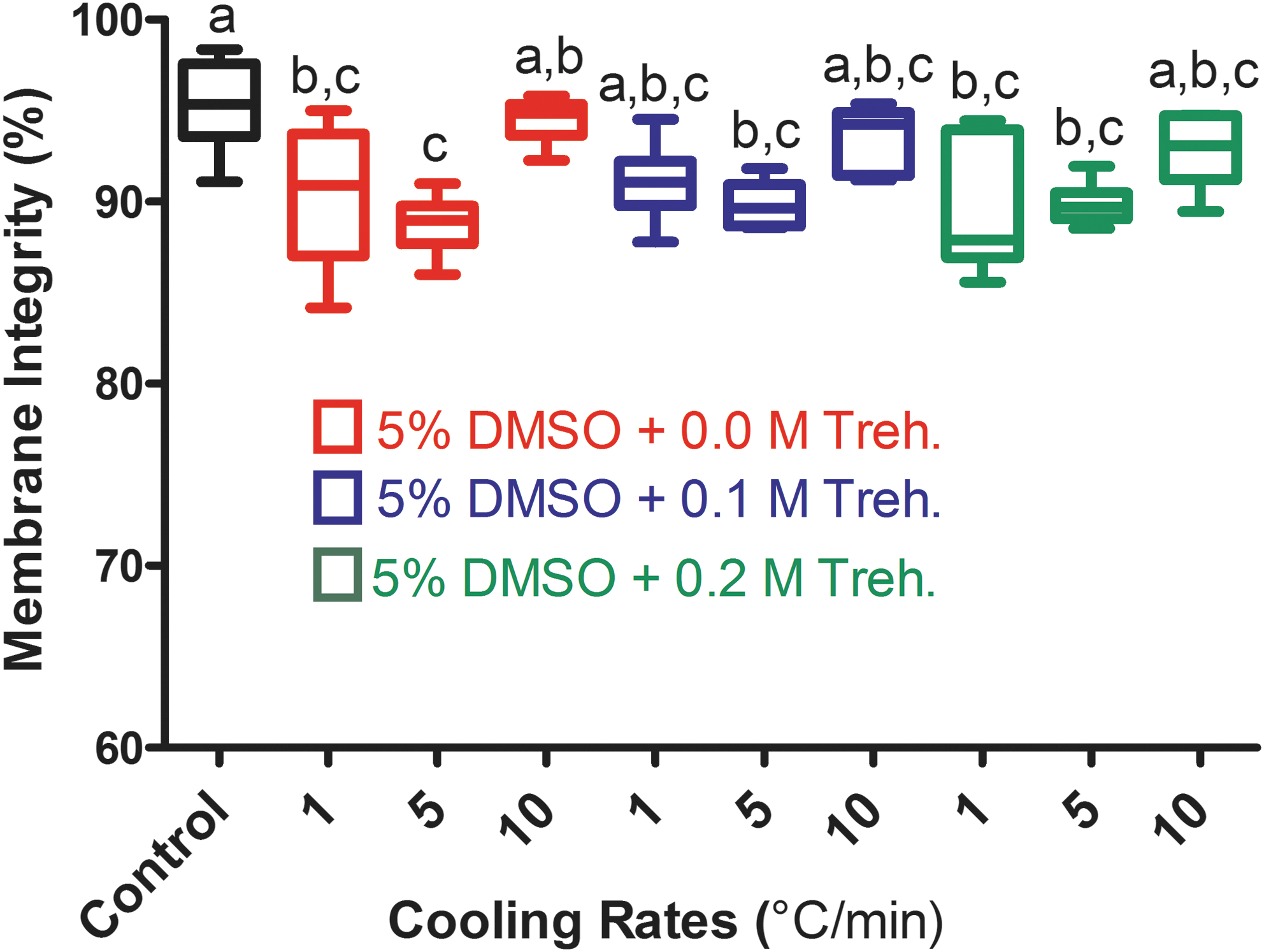
Postthaw survival of embryoid bodies (EBs) as a function of cooling rates and trehalose (Treh.) concentrations. Proportional box plots specify the 25th, 50th (the median), and 75th percentiles, whereas the ends of the whiskers represent the minimum and maximum. Boxes with different letters are significantly different (p<0.05). DMSO, Dimethylsulfoxide.
Overall, all tested conditions yielded high membrane integrity rates ranging from 88.7% to 94.5%. However, the mean membrane integrity rates after freezing at 5°C/min (88.7%, 89.8%, and 89.9% for 0.0 M, 0.1 M, and 0.2 M trehalose, respectively) were significantly lower compared to that (95.3%) of unfrozen controls. The mean survival rates after freezing at 1°C/min were also significantly lower for the 0.0 M (90.3%) and 0.2 M trehalose (90.0%) groups than that of controls, whereas the presence of 0.1 M extracellular trehalose yielded a mean postthaw survival rate (91.1%) statistically similar to controls. Interestingly, the fastest cooling rate (i.e., 10°C/min) resulted in membrane integrity rates for the 0.0 M (94.5%), 0.1 M (93.6%), and 0.2 M trehalose (93.0%) groups, which were comparable to that of unfrozen controls. Taken together, these results suggest that EBs are tolerant to a wide range of cooling rates and extracellular trehalose provides limited additional protection, possibly due to already present beneficial effect of the extracellular matrix as described next.
Comparison of tolerance of intact EBs and dissociated EB cells to cryopreservation-associated stresses
On the basis of the good survival of EBs in the first two sets of cryopreservation experiments, we hypothesized that the extracellular matrix packing the EB cells together may play a beneficial role in cryosurvival of intact EBs. To test our hypothesis, we subjected both intact EBs with abundant extracellular matrix and dissociated EB cells essentially free of extracellular matrix to extreme freezing, which involved freezing and thawing of cells in the absence of any added CPA. The extracellular matrix is mainly composed of sugar-rich proteoglycans and glycoproteins, thus we reasoned that sugars might be the principal cryoprotective components of the extracellular matrix. To examine this, we also performed rescue experiments that included cryopreservation of dissociated EB cells in the presence of 0.2 M trehalose, a natural disaccharide successfully used by many organisms to cope with extreme conditions such as freezing and almost complete drying. 31 Dissociated, but unfrozen, EB cells served as controls. The results of these experiments are summarized in Fig. 4.

Tolerance of intact embryoid bodies (EBs) and dissociated EB cells to cryopreservation-associated stresses. Intact EBs and enzymatically dissociated EB cells were subjected to freezing and thawing in the absence of any added cryoprotective agent (CPA). For the rescue experiments, 0.2 M trehalose was added to Dulbecco modified Eagle medium (DMEM)/F-12, which was used as a base cryopreservation medium. Proportional box plots specify the 25th, 50th (the median), and 75th percentiles, whereas the ends of the whiskers represent the minimum and maximum. Boxes with different letters are significantly different (p<0.05).
Remarkably, all EBs that were not dissociated before freezing still displayed intact EB morphology after thawing. The membrane integrity assay showed that on average, 60.4% of cells in nondissociated EBs were intact after the 2-hr postthaw incubation. This was significantly higher than the postthaw survival (11.5%) of dissociated EB cells while significantly lower compared to the membrane integrity (89.5%) of dissociated but unfrozen EB cells. Interestingly, the addition of trehalose alone to the medium was able to rescue a significant portion of dissociated and then frozen–thawed EB cells with a membrane integrity rate (66.0%) similar to that of intact EBs. These results suggest that the intact EBs are resistant against the stresses associated with freezing and thawing, and sugar residues in their extracellular matrix might be contributing to this.
Postthaw competence of cryopreserved EBs to attach, grow, and differentiate
To address their postthaw functionality, EBs were frozen using three different cooling rates (1°C/min, 5°C/min, and 10°C/min) and different extracellular trehalose concentrations (i.e., 0.0 M, 0.1 M, and 0.2 M) in the presence of 5% DMSO as tested in the second set of experiments. Unfrozen EBs served as controls. After thawing, the EBs were cultured for more than 2 weeks and evaluated for their attachment to culture dishes and subsequent growth, as well as for their differentiation capacity to various cell types. However, the growth rate of individual EBs and the number of differentiating cells per EB were not specifically quantified.
The results of these experiments are shown in Fig. 5. All control EBs were attached to culture dishes and continued to grow and differentiate into various cell types as expected. The majority of the frozen–thawed EBs were also attached and continued to further grow and differentiate (Fig. 6A,B). However, the fastest cooling rate (10°C/min) in the absence of trehalose resulted in a significantly lower attachment rate (51.9%) compared to that of unfrozen controls (100%), whereas the slower cooling rates (i.e., 1°C/min and 5°C/min) had postthaw attachment rates (88.5% and 80.0%, respectively) comparable to that of controls. In the presence of 0.1 M and 0.2 M extracellular trehalose, all three cooling rates yielded the postthaw attachment and growth rates ranging from 69.2% to 93.2%, which were statistically comparable to that of controls. Particularly, the presence of 0.1 M trehalose at a cooling rate of 1°C/min significantly improved the postthaw attachment and growth rate (93.2%) when compared to that of 10°C/min cooling rate in the absence of trehalose, indicating a beneficial effect of trehalose. There was no significant difference in the postthaw attachment and growth rate between the three tested cooling rates when 0.1 M or 0.2 M extracellular trehalose was present. It should also be noted that some of the nonattached EBs also continued to grow and differentiate, although their growth rate was not as good as of attached ones. When taking into account the total number of the growing EBs (attached+nonattached), the differences among the groups were not statistically significant.
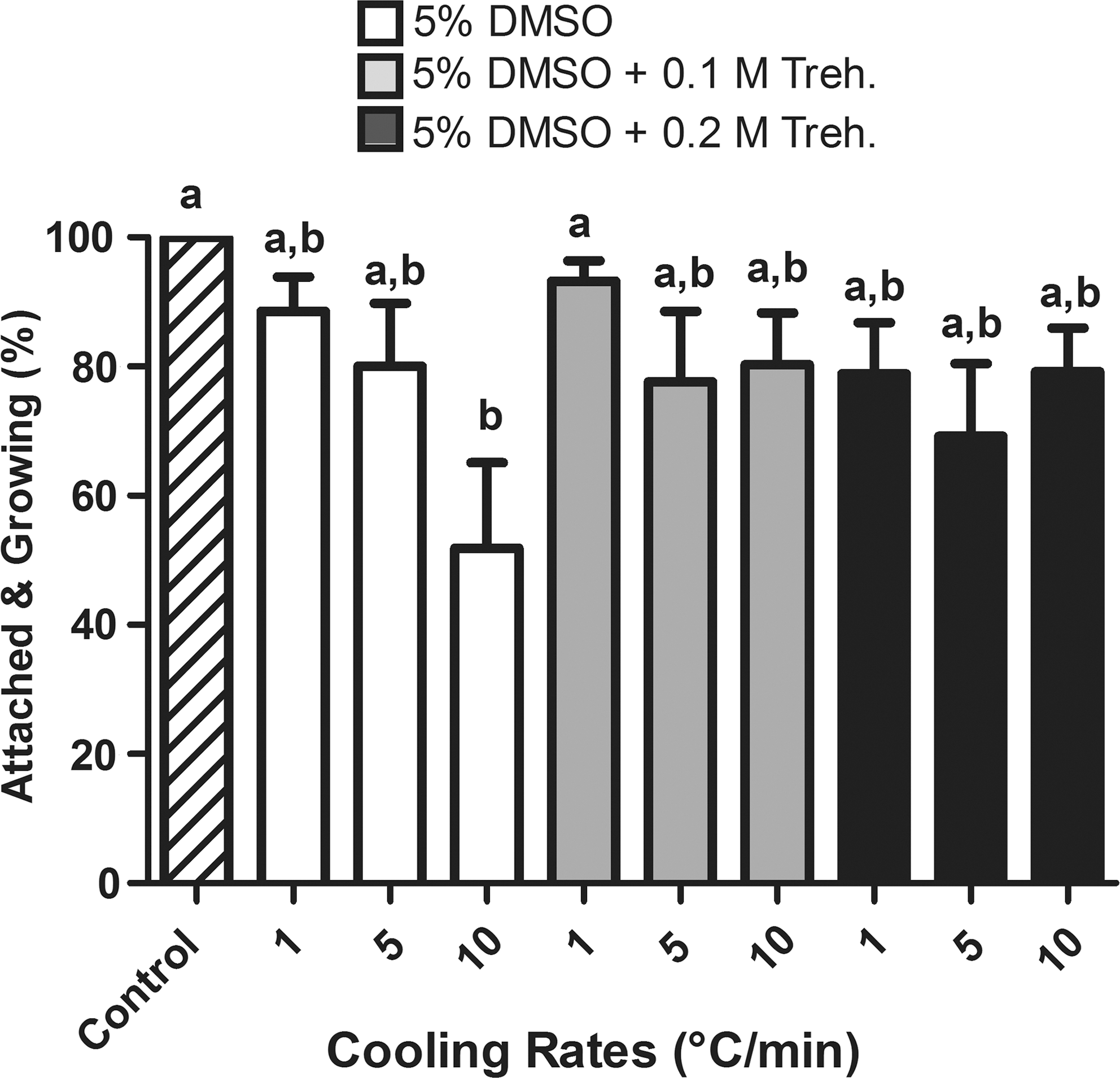
Postthaw attachment and growth of cryopreserved embryoid bodies (EBs) compared to unfrozen controls. Data represent mean±standard error of the mean (SEM). Bars with different letters are significantly different (p<0.05). DMSO, Dimethylsulfoxide; Treh., trehalose.
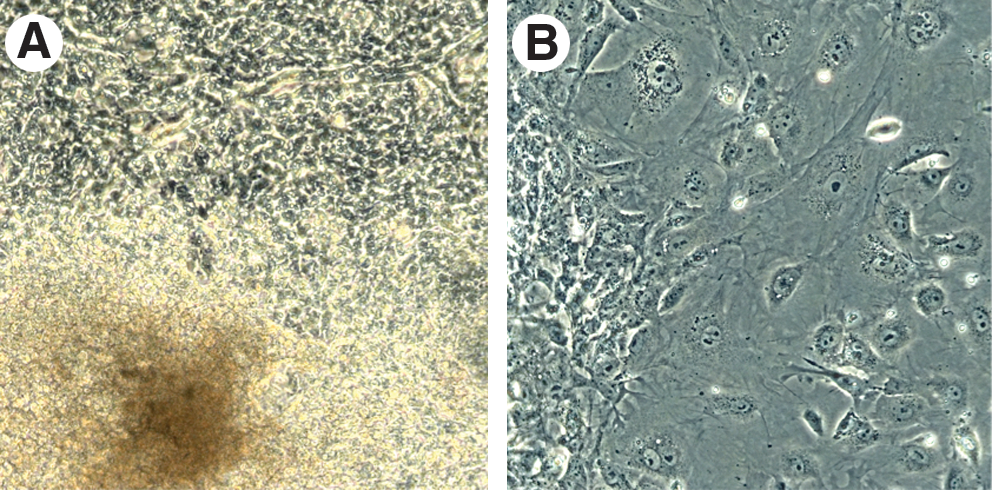
Representative images of cryopreserved embryoid bodies (EBs) showing their postthaw attachment and growth. (
The ability of control and frozen–thawed EBs to differentiate into cell types of three germ layers (i.e., ectoderm, mesoderm and endoderm) was evaluated by bright-field microscopy, as well as by immunocytochemistry. The spontaneous differentiation of both control and frozen–thawed EBs into cardiomyocytes and thus into the mesoderm layer was easily recognizable based on beating activity of cardiomyocytes (see Supplementary Videos). The directed differentiation of EBs into neural (ectoderm) and hepatic (endoderm) lineages was determined by antibody-based fluorescence staining of their markers (i.e., β-tubulin III and albumin, respectively) as shown in Fig. 7. Taken together, spontaneous and induced differentiation of frozen–thawed EBs along with controls revealed that both types of EBs differentiate into cell lineages of three germ layers. Hence, these results indicate that cryopreserved EBs were functional after thawing and retain their pluripotency. However, it should be noted that we have not quantitatively compared the differentiation potential of control and cryopreserved EBs. Therefore, our experiments cannot rule out that the cryopreservation process has no effect on the differentiation potential of frozen–thawed EBs.

Representative images of cryopreserved embryoid bodies (EBs) showing their differentiation into neural and hepatic lineages. (
Discussion
This study demonstrates the feasibility of cryobanking of EBs to enhance basic research and cell-based regenerative therapies by showing high postthaw viability rates and retained pluripotential differentiation capacity of cryopreserved EBs. Furthermore, the present study provides evidence that the presence of an intact extracellular matrix may play a beneficial role in the cryosurvival of EBs.
In general, successful cryopreservation of multicellular structures and tissues is more challenging than that of single-cell suspensions. Yet, EBs, with their compact multicellular structure, survived freezing and thawing surprisingly well, even at low CPA concentrations such as 5% DMSO (≈0.75 M) and fast cooling rates. In fact, the postthaw membrane integrity was highest at the fastest cooling rate tested (i.e., 10°C/min). However, our subsequent studies on the postthaw functionality of EBs revealed that a slower cooling rate (1°C/min) in the presence of 5% DMSO and 0.1 M trehalose yields better postthaw attachment and growth. These results suggest that the membrane integrity assay is useful, but not sufficient to fully evaluate the postthaw viability/functionality of cryopreserved cells.
Unlike in the present study, slow cooling protocols typically require moderately high (≈1.5 M) concentrations of conventional penetrating CPAs, whereas vitrification protocols use very high CPA concentrations (4–6 M). However, penetrating CPAs are known to be toxic at such concentrations. 32 –35 Therefore, the use of lower concentrations of penetrating CPAs is preferred to minimize the potential toxicity of such CPAs whenever satisfactory postthaw survival rates are achievable. In this study, we were able to obtain good postthaw survival using half of the typical penetrating CPA concentration employed in slow cooling protocols. However, it is important to note that a higher CPA concentration (i.e., 10% DMSO) generally yielded consistently good results with less variability.
In nature, a variety of multicellular organisms, including some frogs, nematodes, tardigrades, and brine shrimp can survive freezing and extreme drying. 31,36 The ability to survive such extreme conditions is closely associated with the accumulation of large amounts of sugars, such as trehalose, sucrose, and glucose. 31 Indeed, we and others demonstrated experimentally that sugars afford remarkable protection against cryopreservation- and drying-associated stresses. 37 –40 The experimental findings further indicated that sugars such as trehalose afford their optimal cryoprotection when present both intra- and extracellularly. 30,38,41,42 Nevertheless, when combined with a conventional permeating CPA, the extracellular presence of sugars also provides a considerable cryoprotection. 27 –30 Moreover, even in the absence of penetrating CPAs, extracellular presence of another disaccharide (i.e., 0.35 M sucrose) seems to afford significant protection during freezing and thawing of Chinese hamster cells and bone marrow cells. 43,44
In the present study, the beneficial effect of extracellular trehalose was less pronounced and became apparent in terms of improvement of the postthaw functionality. This might be due to the specific structure of EBs, in which cells are packed together with extracellular matrix comprised of sugar-rich proteoglycans and glycoproteins. Hence, it is possible that the sugar chains present in proteoglycans/glycoproteins afford cryoprotection somewhat similar to that of extracellular trehalose, and thus reduce the need for additional extracellular sugar. We tested this possibility by subjecting intact EBs and enzymatically dissociated EB cells to freezing and thawing in the absence of any penetrating CPA. Indeed, the majority of cells of intact EBs (60.4%) survive such extreme freezing, whereas the overwhelming portion (88.5%) of enzymatically dissociated EB cells were degenerated after freezing and thawing. We were able to rescue dissociated EB cells by adding small amounts (0.2 M) of extracellular trehalose to the freezing medium, supporting the notion that sugar-based properties of extracellular matrix might be contributing to the cryosurvival of intact EBs. It is well known that solutions of sugars and particularly sugar polymers rapidly become viscous upon cooling. Therefore, it is possible that upon cooling, sugar-rich extracellular matrix with its soluble and insoluble components may form a kind of viscous barrier around the cell against ice crystal growth. It is also an intriguing question whether sugar-rich extracellular matrix macromolecules such as proteoglycans and hyaluronan locally undergo glass transition earlier than rest of the freezing solution, resulting in a protective glassy coat around the cell. Clearly, further studies are required to understand the cryoprotective mechanism of the extracellular matrix molecules.
Cryopreservation of human ESCs has been studied extensively to improve unsatisfactory survival results. Until recently, the sensitivity of human ES cells to enzymatic dissociation has been a barrier to their effective passaging and cryopreservation. 6,45 –47 Therefore, human ES cell colonies were mostly cut into small pieces, and then were passaged or cryopreserved as small cell clumps, at least to partially preserve the extracellular matrix. More recently, the introduction of a specific ROCK inhibitor (i.e., Y-27632) permitted more effective passaging and cryopreservation of enzymatically dissociated human ES cell colonies. 46 –49 Unlike human ES cells, EBs have not been the focus of cryopreservation studies, despite potential applications of EB banking. To date, only two abstracts 50,51 and a brief communication 52 have been published on cryopreservation of mouse EBs. Overall, the results of these EB studies are consistent and indicate that EBs can be successfully cryopreserved.
In the present study, we used mouse EBs as a model to explore the feasibility for cryobanking of EBs to facilitate cell-based therapies and basic research. Although we have not tested the cryosurvival of human EBs, it has been shown that in the absence of any specific inhibitor such as Y-27632, nondissociated human ES cells survive freezing and thawing better than their enzymatically dissociated counterparts, 53,54 supporting the protective role of extracellular matrix. Therefore, it is likely that intact human EBs would show similar cryotolerance during their freezing and thawing.
For cryobanking purposes, EBs may need to be cryopreserved in large quantities. In this respect, cryopreservation of EBs in cryovials might be more practical. We also tested this option by cryopreserving EBs in cryovials and subsequently evaluating their postthaw survival. As shown in Supplementary Fig. 2, postthaw survival rates of EBs frozen in cryovials were also high and comparable to those frozen in plastic straws. These results suggest that EBs can also be effectively cryopreserved in cryovials, possibly in large quantities, without compromising their postthaw survival.
In conclusion, the results of the present study indicate that EBs can successfully be cryobanked and distributed, which would facilitate basic research and development of cell-based therapies by offering readily available standardized EBs from pluripotent ES and iPS cells.
Footnotes
Acknowledgments
The authors thank Dr. Kejing Zhang, Ms. Sarah E. Bailey and Ms. Barbara Kraj for their technical assistance. This study was supported by an intramural funding of Medical College of Georgia to A.E.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.