
Abstract
Hyperhomocystinemia could induce tau protein hyperphosphorylation, β-amyloid (Aβ) accumulation, and memory deficits as seen in Alzheimer disease (AD), the most common cause of senile dementia with no effective cure currently. To search for possible treatment for AD, we produced a hyperhomocysteinemia model by vena caudalis injection of homocystine (Hcy) for 2 weeks and studied the effects of acetyl-
Introduction
Acetyl-
Homocysteine (Hcy) is an amino acid in the blood that plays an important role in cellular methylation for proteins, DNA, and lipids. 21,22 Elevation of plasma Hcy is a recognized risk factor for AD. 23 –25 In hyperhomocysteinemia mouse models, many of the AD-like neuropathological changes have been observed, such as cognitive dysfunction, 26 disruption of blood–brain barrier integrity, 27 glutamate toxicity, 28 and an increased level of Aβ. 29 Our recent studies also show that vena caudalis injection of Hcy causes tau hyperphosphorylation by inhibiting protein phosphatase 2A (PP2Ac), 30 a crucial phosphatase in AD-like tau hyperphosphorylation. 31 –36 Hcy also causes Aβ accumulation with the mechanisms involving an enhanced phosphorylation of amyloid precursor protein (APP) at threonine-668. 37 These studies suggest that hyperhomocysteinemia produced by vena caudalis injection of Hcy may serve as a promising rat model for AD studies.
In the present study, we studied whether supplement of exogenous ALC via drinking water for 2 weeks could attenuate the Hcy-induced AD-like histopathologies and behavioral impairments. We found that ALC could rescue the decline of memory retention ability, alleviate hippocampal tau hyperphosphorylation, and attenuate the cortical Aβ accumulation induced by Hcy.
Materials and Methods
Antibodies and chemicals
The primary antibodies employed in this study and their properties are listed in Table 1. ALC was purchased from Sigma-Tau Industrie Riunite (Pomezia, RM, Italy) and dissolved to a final concentration of 50 mg/day per rat in the drinking water before use. Hcy was from Sigma Chemical Co. (St. Louis, MO) and was dissolved to a final concentration of 400 μg/mL immediately before injection.
Mono-, Monoclonal; poly-, polyclonal; WB, western-blot; IC, immunohistochemistry; P, phosphorylated.
Animals and yreatments
Male Sprague–Dawley rats (3–4 months old, 280±20 grams) were purchased from the Experimental Animal Center of Tongji Medical College (Wuhan, China). All animal experiments were performed according to the Policies on the Use of Animals and Humans in Neuroscience Research revised and approved by the Society for Neuroscience in 1995. All rats were kept under standard laboratory conditions: 12 hr light and 12 hr dark, lights on from 6:00 a.m. to 6 p.m., and temperature at 22±2°C. Accessible water and food was provided ad libitum.
A total of 40 rats were selected from 60 rats after 7 days of Morris water maze training according to their swimming pathways and escape latencies on the last day. The rats were divided randomly into four groups (n=10 each). Rats in the four groups were injected by vena caudalis with Hcy (400 μg/kg per day) or saline in the same volume for 14 consecutive days, 30 with or without a simultaneous supplement of ALC (50 mg/day per rat) through drinking water. 38,39 The injection was carried out each day from 9:00 a.m. to 2:00 p.m., including the memory tasks days, and the animals were sacrificed 24 hr after the final injection.
Morris water maze test
The Morris water maze test was used to measure spatial memory retention ability of the rats. 40 –42 The temperature of the room and water was kept at ≈26±2°C. For spatial learning, the rats were trained to find a transparent platform that was located 1 cm below the water surface in the center of the southwest quadrant (target quadrant) for 6 consecutive days. On each trial, the rat started from one of the middle of the no-target quadrants (northeast, southeast, northwest) facing the wall of the pool and ended when the animal climbed on the platform. If a rat failed to find the platform within 60 sec, it was manually guided to the platform and allowed to remain there for 30 sec. The platform, with a scarred surface for helping the rats to climb on, was 40 cm high and 10 cm in diameter and provided the only escape from water. The water surface was 18 cm from the rim of the pool. Through these training sessions, rats acquired spatial memory about the location of the safe platform. A camera, 1.5 meters above the water surface, was fixed to the ceiling of the room and connected to a digital tracking device; it was used to record swimming pathways and latencies of rats. A computer with the water maze software then processed the tracking information recorded by this device. The rats' swimming pathways and escape latencies were recorded on the 7th day of the pretreatment and the same test was performed again 12 hr after the final injection. Another test with the platform removed was performed to record the percent time and path length spent in target quadrant and the number of platform quadrant crosses. During all these trainings and tests, the order of the rats remained unchanged.
Step-down inhibitory avoidance task
For step-down inhibitory avoidance task, the experimental device was a 30-cm×30-cm×30-cm electronic avoidance–response chamber made of Plexiglas on its three sides and hard black plastic on the other. The bottom of chamber was made of a group of parallel stainless steel bars (0.5 cm diameter) spaced 1 cm apart. A rubber platform, 5 cm high and 5 cm in diameter, was placed at a corner on floor to provide the rats a shelter from the electronic attack. Before tests, rats were trained three times from 6:00 p.m. to 10:00 p.m. on the 11th day after the injection. In training sessions, the rats were placed on the platform and received a 1-Hz, 0.5-sec, 36-V DC electronic foot shock immediately after stepping down. The rats were then tested at the same time at 24 hr (1st test) and 48 hr (2nd test) after the training. In test sessions, no foot shock was administered and step-down latencies, errors (placing their all four paws onto the grid), trials (placing two front paws onto the grid), and total time on the platform within 3 min was used as a measure of retention. 43 During all these trainings and tests, the order of the rats also remained unchanged.
Western blotting
The phosphorylation of tau protein at various sites was determined by western blotting, as described previously. 44 The hippocampi, removed from rats' brains cooled on ice, and slice samples were homogenized in an ice-cold buffer consisting of 10 mM Tris-Cl, pH 7.6, 50 mM sodium fluoride (NaF), 1 mM sodium ortho-vanadate (Na3VO4), 1 mM edetic acid, 1 mM benzamidine, 1 mM phenylmethylsulfonyl fluoride (PMSF), and 10 μg/mL each of aprotinin, leupeptin, and pepstatin A. Three volumes of the homogenate was added to 1 volume of the extracting buffer, which consisted of 200 mM Tris-Cl, pH 7.6, 8% sodium dodecyl sulfate (SDS), and 40% glycerol, and the extracts were then boiled in a water bath for 10 min. After that, the extracts were sonicated briefly and centrifuged at 12,500×g for 15 min. The supernatant was stored at −70°C for western blotting analysis. Protein concentration in the supernatants was measured using the bicinchoninic acid (BCA) Protein Assay Kit (Pierce, Rockford, IL) with bovine serum albumin (BSA) as the standard.
The samples were boiled again in a water bath for 10 min with a concentration of 10% β-mercaptoethanol, and 0.05% Bromophenol Blue was added before loading on the gel. Equal amounts of protein in the extracts were separated by 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene difluoride (PVDF) membrane (Millipore, Bedford, MA), which were blocked in 5% (wt/vol) nonfat milk in Tween Tris-buffered saline (T-TBS) containing 10 mM Tris–HCl, 150 mM NaCl, 0.02% (vol/vol) Tween-20, pH 7.5, and probed with various primary antibodies (listed in Table 1) for 48 hr at 4°C. The blots were incubated with anti-mouse or anti-rabbit immunoglobulin G (IgG) conjugated to horseradish peroxidase (1:5,000) for 1 hr at 37°C, visualized by using a chemiluminescent substrate kit (Pierce), and then exposed to CL-XPosure film. The blots were scanned, and the bands were analyzed quantitatively by Kodak Digital Science 1D software (Eastman Kodak Company, New Haven, CT).
Bielschowsky silver and immunohistochemistry staining
According to the modified Bielschowsky silver staining method, 45 sections were immersed in a phosphate-buffered (pH 7.4) 0.5% sodium sulfide solution (PBS) for 3×5 min, followed by 2–4% silver nitrate (AgNO3) solution. After being kept at 37°C and away from light for 30 min, the sections were rehydrated, fixed in formaldehyde for 5 min, and rehydrated again. Finally, an ammoniacal silver ethanol solution was added. For immunohistochemistry, the rats were deeply anesthetized by 6% chloral hydrate, fixed in situ, and perfused through the aorta with 100 mL of 0.9% filtered saline, followed by 400 mL 4% paraformaldehyde in phosphate buffer. Their brains were dissected out and postfixed in the same 4% paraformaldehyde solution at 4°C before being sliced coronally for immunohistochemistry. The fixed brains were sliced coronally through the area of the hippocampus and cortex into 20-μm sections using Vibratome (VT1000s, Leica, Germany). The sections were permeated with 0.3% hydrogen peroxide (H2O2) (in absolute methanol) for 10 min to inhibit endogenous peroxidase, and then blocked with 3% BSA for 30 min. After incubation for 48 hr at 4°C with various primary antibodies (see Table 1 and figure legends), the sections were subsequently incubated with biotin-labeled secondary antibodies for 1 hr at 37°C, and finally developed with Histostain-SP kits (Zymed, South San Francisco, CA) and visualized with 3,3′-diaminobenzidine (DAB; Sigma) as brown color. 30,46 The Bielschowsky silver staining and immunohistochemistry were quantitatively analyzed by Image Pro-plus software (Mediacybernetics; Carlsbad, CA). 47,48
Cell and brain slice culture
For primary culture,
49,50
the rat embryonic (18 days) hippocampi were dissected out swiftly and the cells were dispersed by using collagenase. The separated cells were then cultured in six-well plates that had been precoated with poly-
For brain slice culture, the healthy adult male SD rats were decapitated when deeply anesthetized. 52 Their brains were removed rapidly and put into oxygenated (95% O2, 5% CO2) artificial cerebrospinal fluid (aCSF) consisting of 126 mM NaCl, 3.5 mM KCl, 1.2 mM sodium dihydrogen phosphate (NaH2PO4), 1.3 mM MgCl2, 2.0 mM CaCl2, 11 mM D(+)-glucose, 25 mM sodium bicarbonate (NaHCO3), pH 7.4, for 7–8 min at 4°C. The hippocampi were immediately isolated on ice and sliced coronally into 300-mm-thick pieces with a McIIwain Tissue Chopper (The Mickle Laboratory Engineering Co. Ltd, Gomshall, Surrey, UK). The slices were divided randomly into four groups (Con, Hcy, ALC+Hcy, ALC) and individually incubated at 37°C for 1 hr in aCSF with oxygen supplementation in four bottles. After the slices recovered from stress, 10 mM ALC was added to groups ALC+Hcy and ALC, followed by 0.2 mM Hcy to groups Hcy and ALC+Hcy 1 hr after supplement of ALC. Then 2 hr after final administration, all of the slices were homogenized for western blotting. 53,54
Statistical analysis
Data were expressed as mean±standard deviation (SD) and analyzed by using SPSS 17.0 statistical software (SPSS Inc. Chicago, IL). The one-way analysis of variance (ANOVA) procedure followed by the Student t-test was used to determine the differences of the means between four groups.
Results
ALC rescues the Hcy-induced spatial and fear-motivated memory impairments in rats
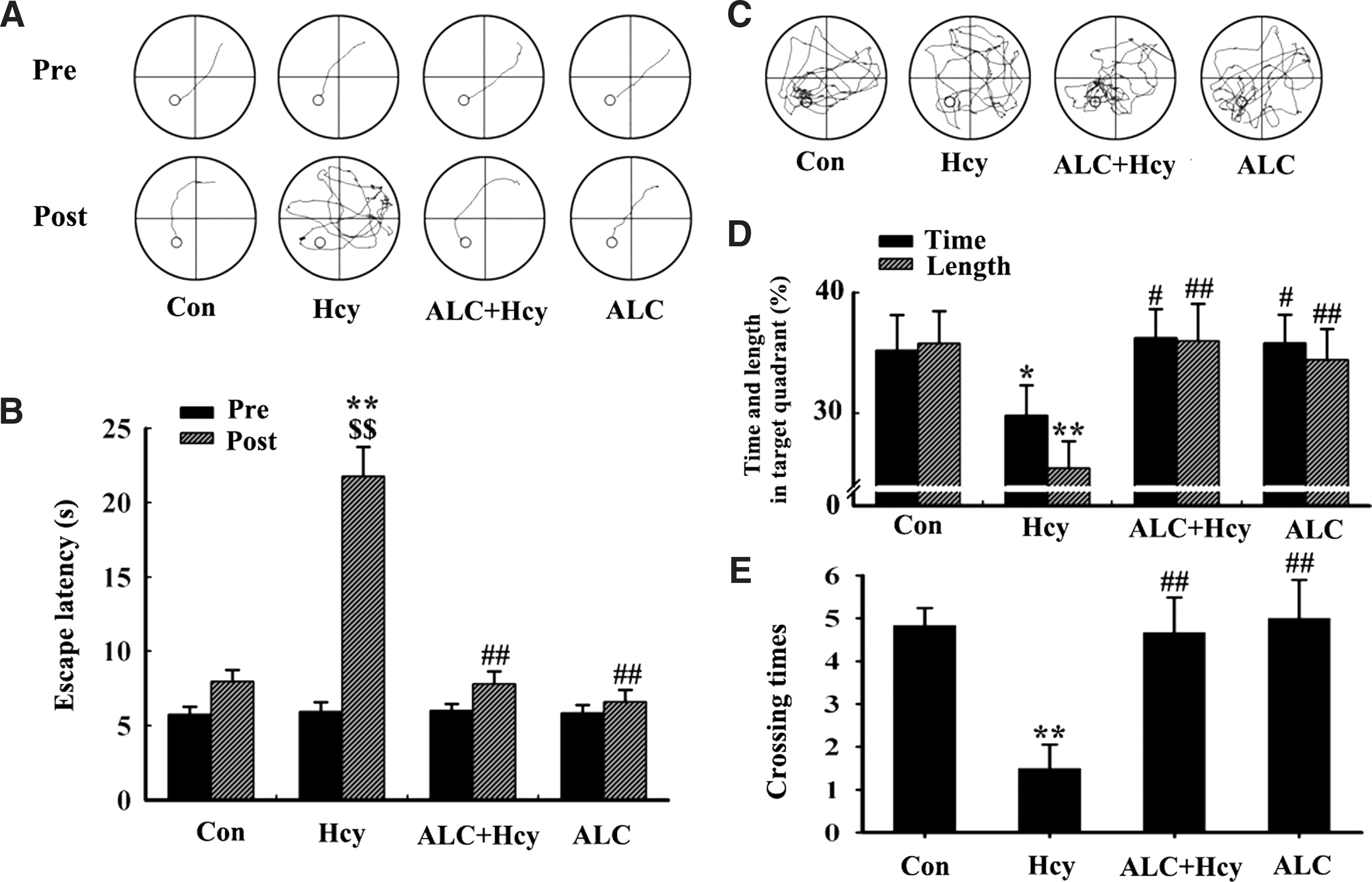
To explore the protective effect of ALC, we first used the Morris water maze test. 55 The rats were chosen as described above (Fig. 1A,B, pre-) and then were injected with 400 μg/kg per day of Hcy or normal saline (NS) for 14 days with or without simultaneous supplement of ALC, and the memory retention was tested at day 15 after the injection (day 22). Compared with the saline-injected controls, the rats injected with Hcy spent much longer time (latency) to find the hidden platform (Fig. 1 A,B, post-). After the platform was removed, less crossed times and trace length were found in the Hcy-treated rats (Fig. 1C–E), which confirmed the spatial memory deficits induced by high Hcy as reported previously. 37 Simultaneous supplement of ALC could efficiently reverse the Hcy-induced spatial memory deficits (Fig. 1A–E).
Acetyl-
We also performed the step-down inhibitory avoidance task to evaluate the effects of Hcy and ALC on the fear-motivated memory of the rats. We found that injection of Hcy could induce the fear-motivated memory deficits demonstrated by the increased step-down errors (placing their all four paws onto the grid) and trials (placing two front paws onto the grid) with a decreased step-down latency, whereas supplement of ALC improved the memory capacity of the rats (Fig. 2). These results demonstrated that ALC could ameliorate memory deficits induced by Hcy.

Acetyl-
ALC reduces Hcy-induced tau phosphorylation
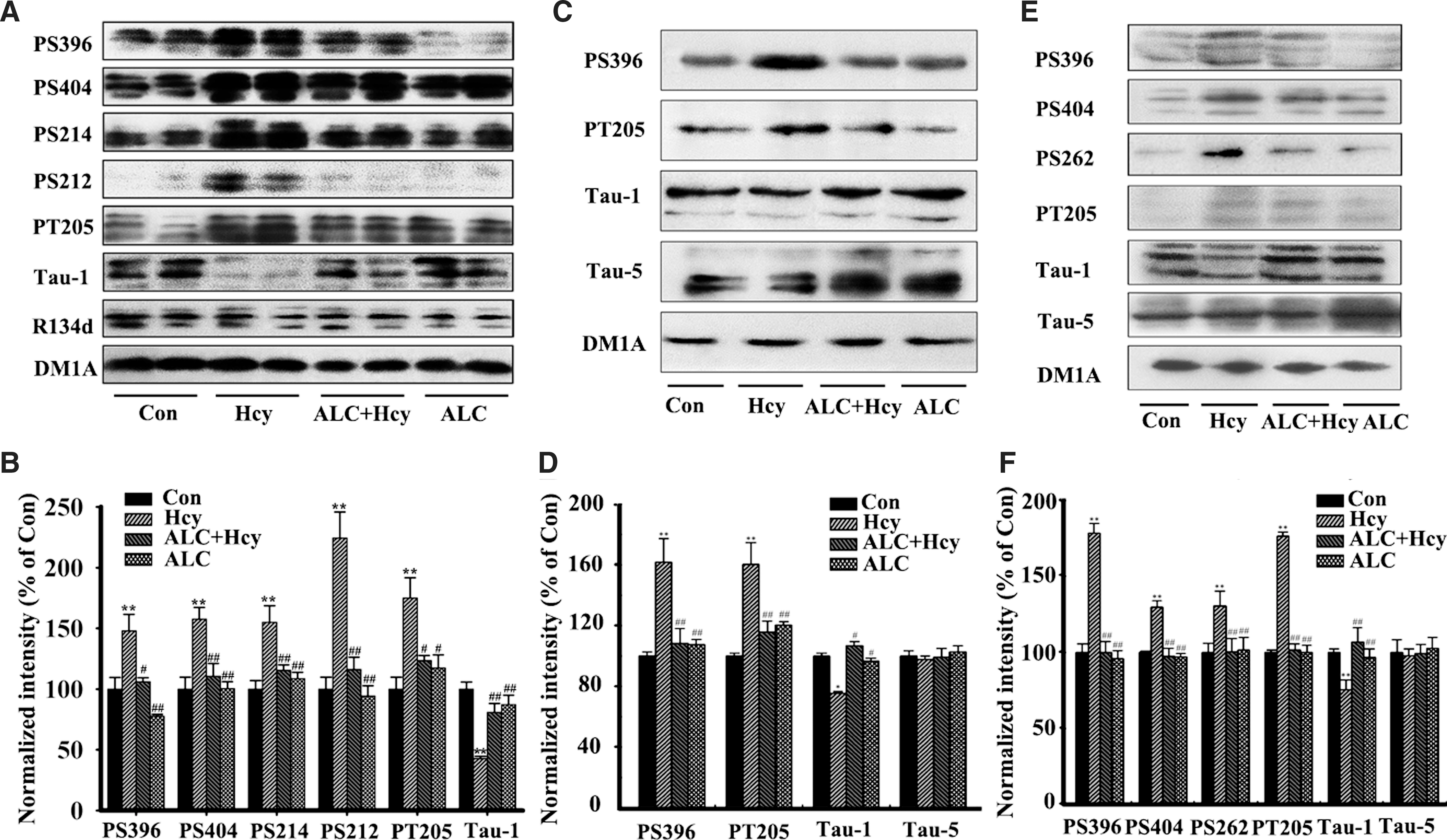
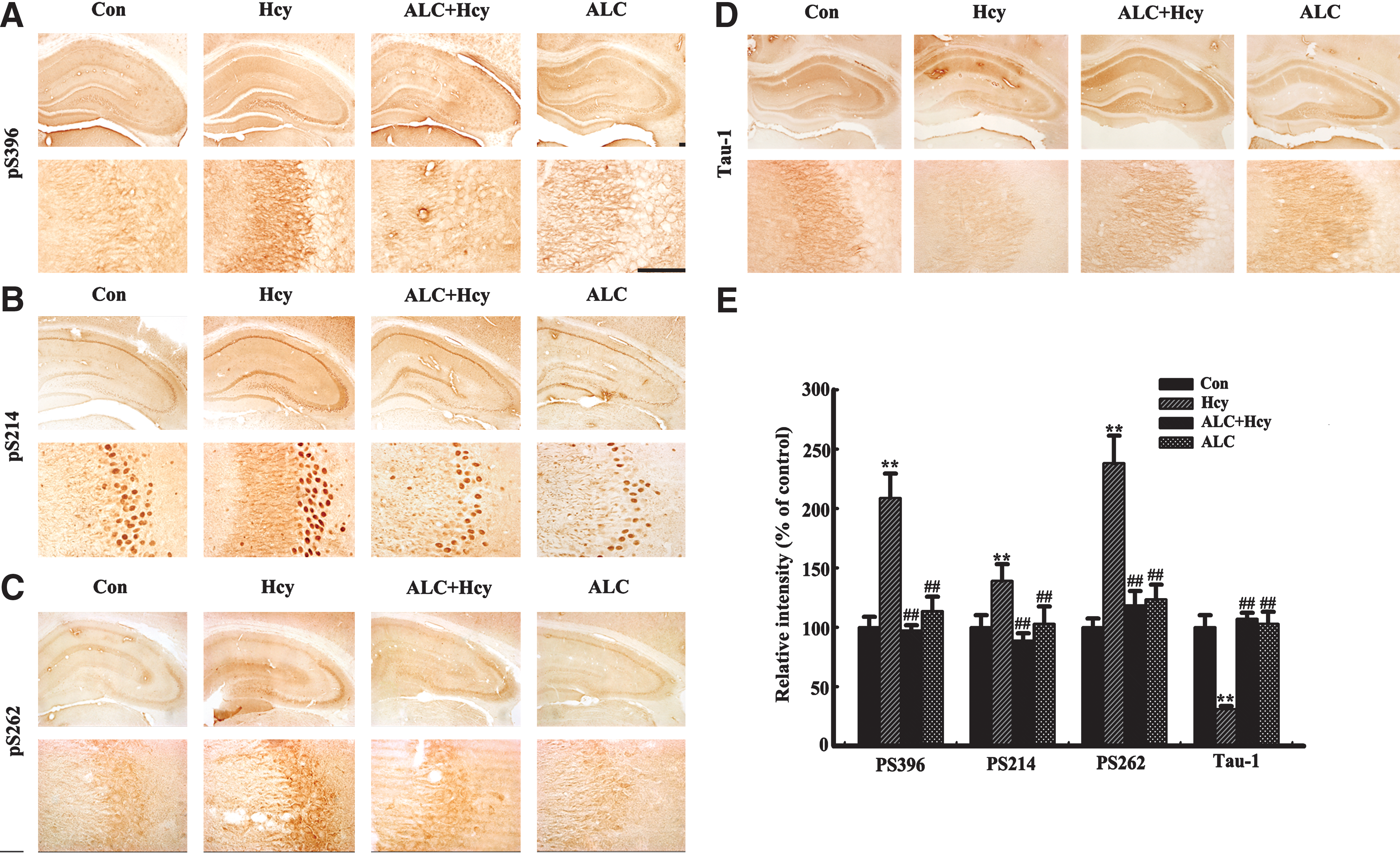
Hcy could induce hippocampal tau hyperphosphorylation at multiple AD-related sites by periphery injection of the rats (Fig. 3A,B), or addition to the primary hippocampal neuron cultures (Fig. 3C,D) and to the acute hippocampal slices (Fig. 3E,F), and simultaneous supplement of ALC could reduce tau protein phosphorylation (Fig. 3). By employing immunohistochemistry, we confirmed that supplement of ALC could attenuate tau protein hyperphosphorylation at Ser396 (PS396), Ser214 (PS214), Ser262 (PS262), and Ser198/199/202 (Tau-1) in the hippocampus, especially in CA3 area (Fig. 4A–D). Quantitative analysis data showed that Hcy increased tau protein phosphorylation to 1.3- to 3.3-fold of the normal levels at different sites, whereas ALC almost reversed the phosphorylation to the normal levels (Fig. 4E). These data strongly suggest that ALC may attenuate the Hcy-induced tau hyperphosphorylation.
Acetyl-
Acetyl-
ALC reduces Hcy-induced Aβ accumulation
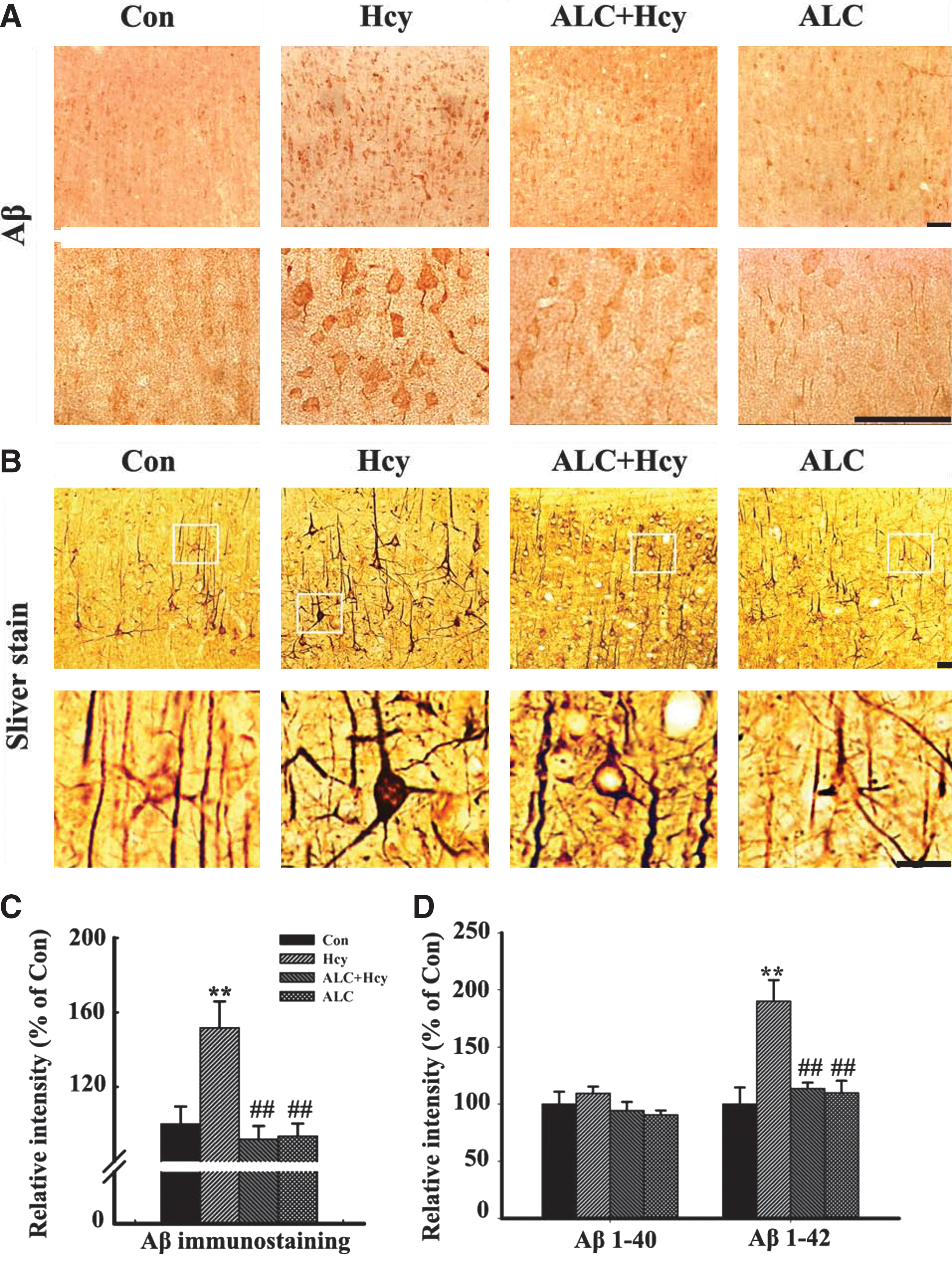
We also observed that ALC could attenuate the Hcy-induced Aβ accumulation in the brain cortex remarkably (Fig. 5A,C). By using the modified Bielschowsky silver staining, we detected an enhanced staining at the neural fibers and the cell bodies in rat cortex treated with Hcy, whereas the accumulation of the argentophilic substances dwindled prominently at the fibers and it almost vanished in the cell bodies after ALC treatment (Fig. 5B). Moreover, this pattern change was coincidently in agreement with the altered Aβ staining (Fig. 5A,C). The enzyme-linked immunosorbent assay (ELISA) analysis performed as described before 56 also showed that the increased Aβ1–42 but not Aβ1–40 level induced by Hcy was restored after ALC supplement (Fig. 5C).
Acetyl-
To explore the possible mechanisms underlying the decreased Aβ accumulation by ALC, we measured the phosphorylation level of Thr668 at APP, which is known to facilitate the amyloidogenic cleavage of APP. 16 We found that the phosphorylation level of APP at Thr668 increased after injection of Hcy, whereas simultaneous application of ALC abated the APP phosphorylation with no obvious changes in total APP (tAPP) probed by APP369 (Fig. 6A,B). These results together suggest that ALC may attenuate Aβ accumulation.
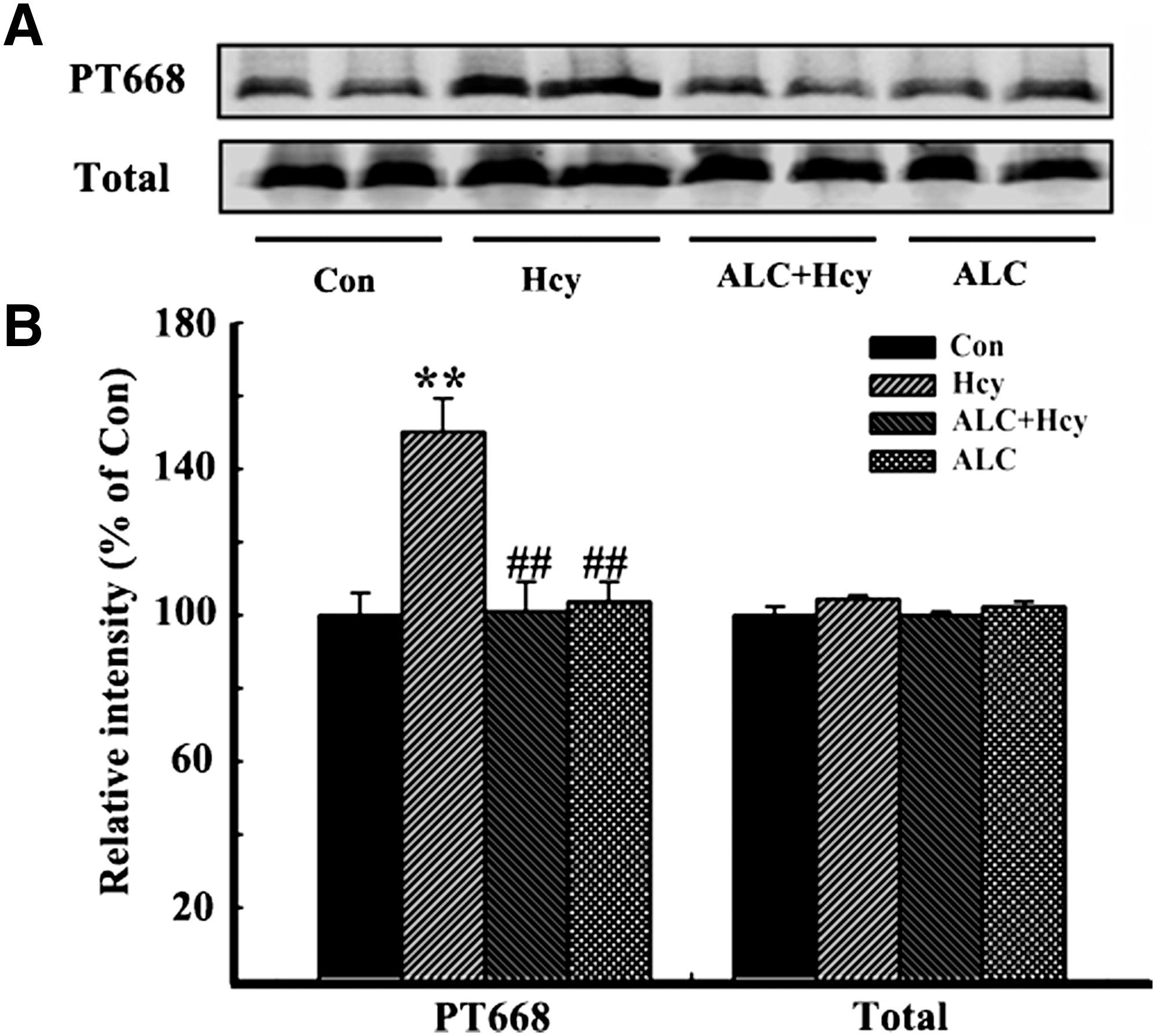
Acetyl-
Discussion
AD is the most common type of neurodegenerative disease, and there is currently no effective treatment for this disorder. In the present study, we investigated in a rat model whether ALC could attenuate AD-like histopathologies and behavioral impairments and the possible underlying molecular mechanisms. We found that supplement of ALC by drinking water for 2 weeks could effectively reverse the Hcy-induced tau protein hyperphosphorylation, Aβ accumulation, and memory deficits in rats. Our data suggest that ALC may serve as a promising candidate for AD therapy.
Protein hyperphosphorylation is caused by an imbalanced regulation of protein kinases and phosphatases. 29 Protein phosphatase 2A (PP2A) is the most crucial phosphatase for AD-like tau and APP dephosphorylation. 27,28 Our previous study demonstrated that Hcy could induce tau hyperphosphorylation at multiple AD sites, and Aβ overproduction by enhanced APP phosphorylation at the Thr668 site via downregulating PP2A 14 and ALC could attenuate the okadaic acid-induced tau hyperphosphorylation by abating PP2A inhibition. 57 Here, we also found that ALC can suppress the Hcy-induced phosphorylation of APP at Thr668. Therefore, we speculate that ALC could ameliorate the Hcy-induced AD pathological changes by rescue of the abated PP2A activity. Additionally, it is well known that ALC could act as an antioxidant and coenzyme for lipid oxidation, thus the underlying mechanisms for the ALC rescue of Hcy-induced AD-like changes deserve further study.
Clinical studies have demonstrated that the amount of neurofibrillary tangles composed of the abnormally hyperphosphorylated tau protein is positively correlated with the degree of dementia in AD patients. 58,59 Animal studies from different laboratories also show that Aβ overproduction is strongly related to the memory impairment. 60 –62 Thus, alleviation of tau hyperphosphorylation and reduction Aβ overproduction have been considered to improve the AD-like memory deficits. 37,63,64 Our data show that ALC could remarkably reduce Aβ accumulation and tau hyperphosphorylation. We believe that the alleviation of Aβ and tau histopathologies may at least partially underlie the improved spatial and fear-motivated memory of the rats. Additionally, ALC can increase the acetylation of histone, 65 which is also vital to memory formation. 66 –68 Further study may verify whether ALC increases the histone acetylation in AD-like model and its potential function.
Taken together, we found in the present study that simultaneous supplement of exogenous ALC significantly could rescue Hcy-induced Aβ accumulation and tau protein hyperphosphorylation with improved memory capacity in rats. We also found that ALC could decrease tau and APP phosphorylation and Aβ.
Footnotes
Acknowledgments
This work was supported in part by grants from the program for 2010 New Century Excellent Talents in University and from the Natural Science Foundation of China (30800342 and 30971478) to Dr. Ling-Qiang Zhu.