
Abstract
The lack of effective drug therapies for motor neuron diseases (MND), and in general for all the neurodegenerative disorders, has increased the interest toward the potential use of stem cells. Among the cell therapy approaches so far tested in MND animal models, systemic injection of human cord blood mononuclear cells (HuCB-MNCs) has proven to reproducibly increase, although modestly, the life span of SOD1G93A mice, a model of familial amyotrophic lateral sclerosis (ALS), even if only few transplanted cells were found in the damaged areas. In attempt to improve the potential efficacy of these cells in the central nervous system, we examined the effect and distribution of Hoechst 33258-labeled HuCB-MNCs after a single bilateral intracerberoventricular injection in two models of motor neuron degeneration, the transgenic SOD1G93A and wobbler mice. HuCB-MNCs significantly ameliorated symptoms progression in both mouse models and prolonged survival in SOD1G93A mice. They were localized in the lateral ventricles, even 4 months after administration. However, HuCB-MNCs were not found in the spinal cord ventral horns. This evidence strengthens the hypothesis that the beneficial role of transplanted cells is not due to cell replacement but is rather associated with the production and release of circulating protective factors that may act both at the central and/or peripheral levels. In particular, we show that HuCB-MNCs release a series of cytokines and chemokines with antiinflammatory properties that could be responsible of the functional improvement of mouse models of motor neuron degenerative disorders.
Introduction
The initial hypothesis governing these studies was that the migration of stem cells to the affected areas and their transdifferentiation toward the neuronal lineage could replace degenerated neurons and therefore restore the functionality of the motor system. However, it is now clear that the effect produced by these cells is likely related to an indirect action (e.g., induction of neurogenesis, growth factors release, antiinflammatory activity) rather than an actual substitution of degenerating motor neurons. 14,15
Therefore, to verify whether an increased availability of HuCB-MNCs in the CNS could result in a further improvement of the therapeutic effect in transgenic SOD1 mutant mice, in the present study, we propose for the first time to transplant HuCB-MNCs into the brain lateral ventricles of two motor neuron disease (MND) mouse models, the transgenic SOD1G93A and the wobbler mice. The wobbler mouse carries a homozygous missense mutation (L967Q) in the Vps54 gene, coding for a protein involved in retrograde transport from late endosomes to the Golgi apparatus. 16 This mutation leads to a rapid and massive loss of motor neurons in the cervical spinal cord region and to a severe atrophy of foreleg muscles and motor impairment.
The effect on the disease progression and on the distribution of HuCB-MNCs in the CNS and their differentiation was investigated at different times in both mouse models. In addition, a detailed analysis of the factors produced and secreted by these cells in vitro was performed to identify the potential beneficial factors in delaying disease progression.
Materials and Methods
In vitro preparation and characterization of HuCB-MNCs
Collection of cord blood
HuCB was collected after informed consent of the mother from full-term newborns. After delivery of the baby, the HuCB was clamped and disinfected, CB was recovered with the placenta in utero into sterile CB collection bags containing 29 mL of citrate-phosphate dextrose (CPD) as anticoagulant (Macopharma, France). The use of human-derived cells in laboratory animals was approved by the Italian Ministry of Health and by an internal ethical committee.
HuCB-MNC purification
HuCB-MNCs were isolated by density gradient (Lympholyte-H 1.077 g/mL; Cedarlane, Ontario, Canada) at 1,600 rpm for 30 min at room temperature. The low-density cell fraction was collected and washed twice at 1,400 rpm for 10 min. The isolated HuCB-MNCs were cryopreserved using a controlled-rate machine in cryovials with 10% final dimethyl sulfoxide (Edwards Lifesciences, CA) and stored in liquid nitrogen. Before the transplantation, the vials were thawed at 37°. RPMI with 20% fetal bovine serum (FBS; Biochrom AG, Germany) was added, and the cells washed in cold RPMI with 20% FBS. The viability was evaluated by Trypan Blue assay (Fluka, Sigma-Aldrich, St. Louis, MO).
Reverse transcriptase polymerase chain reaction and PCR analysis
For reverse transcription polymerase chain reaction (RT-PCR) analysis, total RNA was extracted from HuCB-MNCs by TriZol (GIBCO-Invitrogen, CA) following the manufacturer's instructions. The RNA quality and integrity were evaluated by optical density (A260/280 ratio >1.9). The reverse transcription reaction was performed using the High-Capacity cDNA Archive Kit (Applied Biosystems, CA), while the PCR was performed using the GoTaq DNA Polymerase (Promega, WI) and the primers reported in Table 1. Positive controls were obtained from corresponding fetal tissues.
Two different groups of genes were investigated: embryonic stem cell genes (ES genes) and neuronal lineage−specific genes (neural genes). The table shows the primers used, the amplicon length, and the reference sequence. The presence and absence of the gene expression in our HuCB−MNC samples are reported as+and−.
RT−PCR, Reverse transcriptase polymerase chain reaction; ES, embryonic stem cell; F, forward; R, reverse; NCBI, National Center for Biotechnology information; HuCB−MNC, human cord blood mononuclear cells; OCT4, octamer−binding transcription factor 4; Runx1, runt−related transcription factor 2; Rex1, reduced expression gene 1; Sox−2, sex determining region Y box 2; hTert, human telomerase reverse transcriptase; FGF4, fibroblast growth factor−4; Pax7, Paired box protein; GFAP, glial fibrillary acidic protein; GAL C, galactocerebroside; NRCAM, neuronal cell adhesion molecule; NSE, neuron−specific enolase; TRKA, neurotrophic tyrosine kinase receptor A; MAP2, microtubule−associated protein 2; MORF, monocytic leukemia zinc finger protein−related factor; GAP43, growth−associated protein 43; GAPDH, glyceraldehyde 3−phosphate dehydrogenase.
Immunofluorescence
Immunocytochemical analysis was done by evaluating the following markers: Polyclonal human specific nestin (1:200, Chemicon, CA), monoclonal human specific glial fibrillary acidic protein (GFAP, 1:100, Chemicon), human specific monoclonal CD11β (1:50, Becton Dickinson, CA), and monoclonal β-tubulin III (1:200, Chemicon). The nuclei were stained with 4′,6-diamidino-2-phenylindole, dilactate (DAPI, 100 ng/mL, Sigma-Aldrich). To avoid possible false-positive results, control experiments were done using the primary or the secondary antibodies alone. All experiments did not produce staining in each well examined.
Enzyme-linked immunosorbent assay analysis
To evaluate whether HuCB-MNCs expressed relevant molecules for the physiological survival of motor neurons, the measurement of brain-derived neurotrophic factor (BDNF) protein level was evaluated on 20 μg of cell lysates by enzyme-linked immunosorbent assay (ELISA; BDNF Emax ImmunoAssay System, Promega, CA). A volume of 100 μL of lysis buffer (150 mM NaCl, 20 mM Tris, 1% Triton X-100, containing the Complete Mini protease inhibitor cocktail (pH 8, Roche, Basel, Switzerland), was added to lyse HuCB-MNCs and to isolate proteins. After acid treatment, the samples were plated in triplicate in a 96-well plate (Nunc MaxiSorp™ plate, NY) coated with the specific anti-human antibody. The following procedure was employed to improve the neurotrophic detection. After overnight incubation, a tertiary anti-immunogloblulin Y (IgY) horseradish peroxidase (HRP) conjugate antibody was added and, finally, the BDNF amount was detected using 3,3′,5,5′-tetramethylbenzidine (TMB, A450). Pure neurotrophin proteins provided in the kit were used to establish standard curves for quantification.
Cytokine antibody array
The simultaneous release of 43 cytokines was detected on HuCB-MNCs conditioned medium (CM) obtained by culturing HuCB-MNCs for 2 days in standard culture conditions in the presence of Dulbecco modified Eagle medium (DMEM; Sigma) plus 10% FBS at 37°C. The same medium including FBS, but without cells, was incubated for 2 days under the same conditions and was used as control.
The RayBio Human Cytokine Antibody Array IV (RayBiotech Inc., GA) was performed following the manufacturer's instructions. Briefly, 1 mL of each sample was added to the membrane overnight, after a blocking step, and successive incubations with the specific antibodies were performed at room temperature. The chemiluminescent signals were revealed on Hyperfilms (Amersham Bioscience, UK) using the kit detection buffer, and the cytokine signal intensity was measured by spot densitometry using a specific program (Quantity One, BioRad, CA). The analysis was performed using Prism software (GraphPad Software, CA). Two separate analyses for each single condition were carried out. The ratio between the mean value of CM and basal CM was measured to evaluate the degree of cytokines expression in presence of HuCB-MNCs.
Flow cytometry
HuCB-MNCs were stained using a battery of selected and well-standardized antibodies for 20 min at room temperature in the dark, washed with phosphate-buffered saline (PBS) at 1,400 rpm for 10 min, resuspended in 0.1 M PBS, and analyzed with a Cytomics FC500 flow cytometer (Beckman Coulter, CA). The antibodies used were: CD3-PE (Beckman Coulter), CD14-FITC (Becton Dickinson, CA), CD34-FITC (Becton Dickinson), CD44-FITC (Becton Dickinson), CD45-PC7 (Beckman Coulter), CD73-PE (Becton Dickinson), CD90-APC (Becton Dickinson), CD133-PE (Miltenyi Biotec, Germany), CD146-FITC (BioCytex, France), SSEA4-PE (Becton Dickinson), PDGF-Rbeta PE (Becton Dickinson), NG2-PE (Beckman Coulter), and alpha-SMA FITC (Sigma-Aldrich,). The isotype-matched immunoglobulins IgG1 PE-FITC (Chemicon), IgG1-PC7 (Beckman Coulter), and IgG1-APC (Becton Dickinson) were used as negative controls under the same conditions. After staining, the cells were washed once with PBS containing 0.1% bovine serum albumin (BSA). At least 50,000 events were acquired with a Cytomics FC500 flow cytometer (Beckman Coulter), and plots were generated using CXP analysis software. Forward scatter (FSC) and side scatter (SSC) light characteristics were used to gate the 'live' cells and exclude debris and dead cells. Within this gate, the MNCs were analyzed for their marker expression.
Hoechst 33258 labeling
HuCB-MNCs were incubated in vitro with the fluorescent marker pentahydrate-bis-benzimide Hoechst 33258 (Sigma-Aldrich, MO). The cells were suspended in PBS buffer containing 0.2% human serum albumin (HSA) and the marker Hoechst 33258 (2 μg/mL) and subsequently incubated 1 hr at 37°C. After the incubation, the cells were washed. The cell viability was evaluated by the Trypan Blue assay (Sigma-Aldrich). Until the transplantation in lateral ventricles of mice, cells were maintained in RPMI with 20% FBS. Just before transplantation they were centrifuged and resuspended in sterile PBS.
Ex vivo expansion of Hoechst 33258 HuCB-MNC and viability evaluation
To evalute cell survival after fluorescent labeling, Hoechst 33258 HuCB-MNCs were cultured with the medium CellGro (Cell Genix, Germany) added with 10% inactivated AB plasma and expanded in vitro for 14 days by the addition of the following cytokines: Interleukin-6 (IL-6, 10 ng/mL, PrepoTech EC., UK), stem cell factor (SCF, 50 ng/mL, Endogen Pierce, IL), thrombopoietin (TPO, 10 ng/mL, PrepoTech), and FLT-3 ligand (50 ng/mL, Endogen Pierce). Afterward, the cells were collected and centrifuged at 1,400 rpm for 10 min. The cell viability was assessed by 7-aminoactinomycin (7-AAD; Beckman Coulter).
In vivo characterization
Animal models
Mice were maintained at a temperature of 21±1°C, relative humidity 55±10% and 12 hr of light. Food (standard pellets) and water were supplied ad libitum Procedures involving animals and their care were conducted in conformity with the institutional guidelines that are in compliance with national (D.L. No. 116, G.U. Suppl. 40, Feb. 18, 1992, Circolare No. 8, G.U., 14 Luglio 1994) and international laws and policies (EEC Council Directive 86/609, OJ L 358, 1 Dec.12, 1987; NIH Guide for the Care and Use of Laboratory Animals, U.S. National Research Council, 1996). All efforts were made to minimize the number of animals used and their suffering. When mice showed the first pathologic symptoms (reduced hind-limb adduction after tail suspension), food and water were supplied inside the cage. The protocol for the use of laboratory animals was approved by the Italian Ministry of Health and by an internal ethical committee.
Female transgenic SOD1G93A mice derived from the line originally obtained from Jackson Laboratories (B6SJL-TgNSOD-1-SOD1G93A-1Gur) expressing about 20 copies of mutant human SOD1 were bred (more than 30 generations) and maintained on a C57BL/6JOlaHsd strain at Harlan Italy S.R.L., Bresso (MI), Italy. As we previously reported, the mean survival of these C57BL6JOla SOD1G93A mice is much longer than the original 1Gur mice from Jackson Laboratories due to the different genetic background, as already reported. 17 Transgenic mice were identified by PCR performed on DNA from tail biopsies. 18 To maintain homogeneity in the number of transgene copies in our colony, we quantified the transgene hSOD1 compared to the endogenous murine SOD1 in each SOD1G93A male mouse breeder by quantitative RT-PCR analysis using the TaqMan assay. We calculated the delta Ct for each mouse and excluded those exhibiting a delta Ct that was not included in a range established in the laboratory on the basis of the life span of a group of well-characterized SOD1G93A male mice.
Wobbler mice were originally obtained from NIH Genetics and then bred at the Charles River Italy, Calco (LC), Italy. Because heterozygous mice do not show any phenotypic difference compared to homozygous healthy littermates, heterozygous founders were designed by genotyping. 19
Cell transplantation
SOD1G93A mice
Female SOD1G93A mice (10 weeks old, n=32) were randomly assigned to different experimental groups. One experimental group (n=21) received 500,000 HuCB-MNCs resuspended in 10 μL of sterile PBS into the lateral cerebral ventricles by intracerebroventricular (ICV) administration. Another group (n=11) received the same volume of sterile PBS. Animals were anesthesized by single intraperitoneal (i.p) injection of Avertin (250 mg/kg body weight) dissolved in sterile saline solution before the surgical procedure. To avoid an excessive load in a single ventricle, 5 μL was injected for each side. The following stereotaxic coordinates were used (anterior posterior 0.0 mm from the bregma; lateral±1.0 mm from the bregma; deepness −3.0 mm from skull surface). One day before the transplantation and for the entire duration of the experiment, both groups received cyclosporine (10 mg/kg i.p.) once daily as immunosuppressant to avoid rejection of human cells.
Animals receiving HuCB-MNCs were randomly assigned to each experimental group as follows: 4 mice were sacrified at 24 hr and 5 mice at 4 weeks after transplantation (11 and 14 weeks of age, respectively) to examine the distribution and tracking of transplanted cells. Mice at 14 weeks of age exhibited only very mild signs of neuromuscular impairment, such as tremors of the hind limbs when they are raised by the tail. We consider them in a preonset stage. The other 23 SOD1G93A female mice receiving HuCB-MNCs (n=12) or PBS (n=11) were examined twice weekly for the clinical progression and motor dysfunction starting from the 15 weeks of age. Six HuCB-MNC- and 5 PBS-treated mice, taken randomly from each group, were sacrified at 22 weeks of age when PBS-treated mice displayed a decline in the stride length and in RotaRod test performance for two consecutive trials. The distribution and the intensity of labeled cells were examined in the brain and spinal cord. To track the fate of ICV-injected HuCB-MNCs in SOD1G93A, a distribution study of Hoechst 33258 HuCB-MNC–positive cells was performed on cryostatic sections (30 μm thickness). Spinal cords were also examined for the count of motor neurons in the lumbar region.
The other 6 mice for each group were monitored for disease progression until their death. One mouse transplanted with HuCB died early after transplantation for causes unrelated to MND and therefore was not included in the statistical analysis.
Wobbler mice
At the end of the third week of life, wobbler mice carrying a homozygous mutation (L967Q) for the Vps54 gene (59), were differentiated from the other littermates by the drastic reduction of growth (in length and body weight) and by the appearance of sustained tremors. These signs preceded motor deficits and enabled us to follow muscular impairment along the duration of the clinical progression. Thirty wobbler mice (4th week of life) were randomly assigned to two different experimental groups. One experimental group (n=18) received 300,000 HuCB-MNCs resuspended in 5 μL of sterile PBS into the lateral cerebral ventricles, 2.5 μL for each ventricle, under Avertin anesthesia (250 mg/kg i.p.). Another group (n=12) received the same volume of sterile PBS. Because of the small size of brains of 4-week-old wobbler mice, the sterotaxic coordinates were modified (anterior posterior 0.0 mm from the bregma; lateral±0.8 mm from the bregma; deepness −2.5 mm from skull surface). For each experimental group, the same number of males and females was recruited, because no sex-related difference in the evolution of the wobbler disease has been reported.
Six mice were sacrificed 1 week after HuCB-MNC transplantation for cell tracking. Twelve mice for each experimental group were examined weekly by standardized behavioral trials (see below) to monitor their clinical progression and motor dysfunction. Six animals for each group were sacrificed at the 10th week of age, which corresponds to the late symptomatic phase in PBS-treated mice. The remaining 6 animals for each group were monitored for disease progression until the 20th week of age.
Cyclosporine was given for the duration of the behavioral observation. Due to the difficulty of wobbler mice in sustaining a daily i.p. treatment, the cyclosporine (40 mg/kg per day) was administered at a concentration of 0.1 mg/mL dissolved in the drinking water containing 1% ethanol and 0.5% Tween-20.
Immunofluorescence of HuCB-MNCs in grafted tissues
SOD1G93A and wobbler mice receiving HuCB-MNCs (n=6 for each group) were analyzed at the intermediate stage of the disease, respectively, at 22 and 10 weeks old to test the possible lineage commitment of transplanted cells. Free-floating sections were treated with a solution of 0.1% Triton X-100, 10% normal goat serum (NGS, Vector, CA), and PBS for 1 hr followed by an incubation overnight at 4°C, with a primary antibody against nestin, β-tubulin III, or GFAP, as previously reported above. In addition, a primary monoclonal antibody directed against human leukocyte antigen (HLA) class I (Immunological Science, Italy, 1:500) was used to reveal the presence of human cells in the brains of both wobbler and SOD1G93A mice. The sections were mounted on slides with Fluorsave (Calbiochem, NJ) and analyzed. The images were taken by a microscope (BX81, Olympus, NY) equipped with a F-view II CDD camera (Olympus, NY), and the co-localization was analyzed by a unmixing fluorescence module (Olympus, NY) to exclude the possible crosstalk between the channels.
Disease progression assessment
For both SOD1G93A and wobbler mice, body weight measurements and behavioral trials were performed twice weekly (Monday and Thursday morning) by the same operator blinded to the treatment.
SOD1G93A mice
Transgenic SOD1G93A female mice were tested for quantitative assessment of the motor behavioral deficit by measuring the stride length and RotaRod until death. To measure stride length, mice were trained to walk a 75-cm-long ramp raised to a height of 13 cm at one end. A bright light was placed at the base of the ramp to provide an aversive stimulus. The mice's hind feet were painted with children's poster paints, and the tracks left as they ran up the ramp were recorded on paper tape lining the floor of the ramp. Stride length was defined as the distance between successive right-to-right and left-to-left footprints.
For the RotaRod test, SOD1G93A mice were trained to remain on the rod and habituated to the handling involved for at least 1 week before testing. On the day of testing, mice were transferred to the RotaRod room at least 15 min before the test. Performance was evaluated as the time (sec) spent on the rotating rod (12 rpm) without falling off (RotaRod treadmill for mice; Ugo Basile, Comerio, VA, Italy). The test was stopped after 3 min. Mice that fell off before the third minute were tested three times, with at least 5 min between tests. This schedule was used during the training period and after transplantation. SOD1G93A mice were sacrificed if they were unable to right themselves within 30 sec when laid down on their side. This time was used for calculating the survival curves. The body weight was recorded for all animals during treatment.
Wobbler mice
The following tests were employed to evaluate the neuropathological progression affecting wobbler mice: 1. Paw and walking abnormality: Both the paw abnormality and the walking abnormality tests were observational. The operator assigned a score to these parameters, scaled from 0 to 4, on the basis of the severity of abnormalities. The paw position was graded as follows: 0, normal; 1, retracted digits; 2, curled digits; 3, curled wrists; 4, forelimb flexed to body. The walking pattern was graded as follows: 0, normal; 1, trembling (tremor without gait disturbance); 2, wobbling (gait disturbance); 3, curled-paw walking; 4, jaw walking (no use of front paw). 2. Running speed: Mice ran over an inclined platform (75-cm-long ramp inclined at one end to a height of 13 cm) stimulated with a gentle pressure on the tail (adverse stimulus). The running time was defined as the shortest time to reach the top of the platform from the bottom. Healthy mice rapidly improved their performance on the test until they reached the top of the platform in few seconds (1–3 sec). Wobbler mice needed a longer time to reach the top of the platform. Moreover, these animals showed a marked worsening of their performances due to the progressive muscular atrophy in the forelegs. 3. Grip strength: Mice were lifted by the tail and allowed to grasp with both forelegs to a horizontal bar, which is connected to a mechano-electric transducer (Basile). The grip strength of the front paws was measured at the point at which the mouse released the horizontal bar as a result of a gentle traction applied by the operator. Healthy mice can record values higher than 100 g, whereas values recorded by wobbler mice were very low (<20 g) and drastically reduced during symptom progression. When animals were no longer able to grip the bar, grip strength was recorded as 0 g. Values of grip strength were normalized by dividing each value by body weight to control for weight differences between wobbler and healthy mice.
Because wobbler mice develop early and severe atrophy of forelegs without a clear impairment of hindleg muscles, the classical RotaRod test cannot be considered a reliable tool to evaluate the clinical progression in these mice.
Histopathology of spinal cord
Animals were perfused transcardially with 4% paraformaldehyde (w/v) in 0.1 M PBS (Sigma-Aldrich), and the following steps were carried out according to the procedure previously described. 20 Immediately after perfusion, spinal cords and brains were rapidly removed and postfixed for 4 hr in the same fixative (4°C) and then dehydrated and cryoprotected with serial steps in 10%, 20%, and 30% sucrose in 0.1 M PBS at 4°C until they sank. Subsequently, brain was frozen in n-pentane at −45°C and spinal cord included in OCT (Tissueteck, Netherlands). All tissues were stored at −80°C until analysis. Due to the selective atrophy of forelegs in wobbler mice, biceps muscles were removed, dried by blotting paper, and immediately weighed.
To evaluate the neuroprotective effects of HuCB-MNCs on motor neurons in both animal models, the immunohistochemistry of choline acetyltransferase (ChAT) was applied to the most affected spinal region of SOD1G93A mice (L2–L4) and wobbler mice (C2–C6) . Serial 30-μm-thick sections were cut, and ChAT immunostaining (mouse monoclonal antibody, Immunological Science, Italy, 1:1,000) was carried out according to the protocol described by Houser et al. 21 in one every five sections. ChAT-positive neurons counting was carried out in SOD1G93A mice sacrificed at the 22nd week of age and in wobbler mice sacrificed at the 10th week of age according to the previously described method. 22 Olympus-DB software, coupled to the microscope camera, was used to count the ChAT-positive cells >250 μm2. The number of ChAT-immunopositive motor neurons was calculated for each single section, and the means of these determinations were used as individual data for statistical analysis. Nontransgenic littermates and healthy NFR mice (n=3–5 for each group ) were examined to assess the extent of motor neuron loss in SOD1G93A and wobbler mice.
Statistics
All data are expressed as mean±standard error of the mean (SEM), except for the survival of SOD1G93A expressed as mean±standard deviation. Analysis of survival length in SOD1G93A mice was performed using the log-rank (Mantel–Cox) test and the nonparametric analysis using a Gehan–Breslow–Wilcoxon test. Body weight and gait scores were analyzed by analysis of variance (ANOVA) for repeated measures (time) and different groups (treatments), followed by the post hoc Tukey test.
Paw abnormality, walking abnormality, and RotaRod values were compared by using the Wilcoxon matched-pairs test. Motor neuron counts for both wobbler and SOD1G93A mice, and biceps weight for wobbler, were analyzed with the unpaired Student t-test. p values <0.05 were considered significant. All statistical analyses were done using the GraphPad Prism version 4.00 for Windows (GraphPad Software, CA).
Results
Characterization and preparation of HuCB-MNCs
HuCB-MNC in vitro characterization
HuCB-MNCs were extensively characterized by RT-PCR analysis for the expression of markers of stamina and neural origin (Table 1) and by flow cytometry for the expression of hematopoietic and mesenchymal SC marker (Table 2). As shown in Table 2, transplanted cells contained cell subpopulations negative for the expression of the hematopoietic marker CD45, but positive for typical mesenchymal and perivascular SC antigens such as CD44, CD90, CD146, and PDGFRβ. Interestingly, a high percentage of HuCB-MNCs expressed the embryonic stem cell markers SSEA-4. RT-PCR analysis revealed that HuCB-MNCs were positive for the main embryonic stem cell markers, namely Oct-4, Runx-1, Rex1, and the catalytic subunit of telomerase (hTert). Moreover these cells clearly demonstrated a neuro-glial potential (RT-PCR analysis). As reported in Table 1, HuCB-MNCs expressed many lineage-specific markers, such as monocytic leukemia zinc finger protein-related factor (MORF), neuron-specific enolase (NSE), galactocerebroside (GalC), neural cell adhesion molecule (NRCAM), neurotrophic tyrosine kinase receptor A (TRKA), microtubule-associated protein 2 (MAP2), growth-associated protein 43 (GAP-43), and βtubulin III. We could not detect the developmental CNS transcription factor sex-determining region Y-box 2 (Sox-2) nor the neural stem cell marker Musashi-1 nor other neural typical transcripts such as nestin, GFAP, and neurogenin.
Moreover, to analyze whether Hoechst 33258 incorporation could impair cell survival, the viability of labeled and nonlabeled HuCB-MNCs was compared. No difference on the percentage of survival was observed in vitro between the two experimental groups. The median (and range) of the viable Hoechst 33258+ HuCB-MNCs was 99.0% (98.5–99.6) whereas in the Hoechst 33258-HuCB-MNCs it was 99.5% (99.1–99.9).
Neurotrophic factors and cytokines detection
In the HuCB-MNCs, the level of BDNF was 104.32 pg/mL. BDNF secretion was also found in the HuCB-MNC CM by the cytokine array (Fig. 1). Very interestingly, the array membrane identified 36 different growth factors/cytokines released by HuCB-MNCs. They included both angiogenic (fibroblast-growth-factor [FGF]-4 and -6, hepatocyte growth factor [HGF], insulin-like growth factor binding protein [IGFBP]-1 and -3) and neutrophic factors (neurotrophin [NT]-3 and -4, BDNF, neutrophil-activating protein-2 [NAP-2]). A detailed description of the main factor expressed and released by HuCB-MNCs is reported in Table 3.
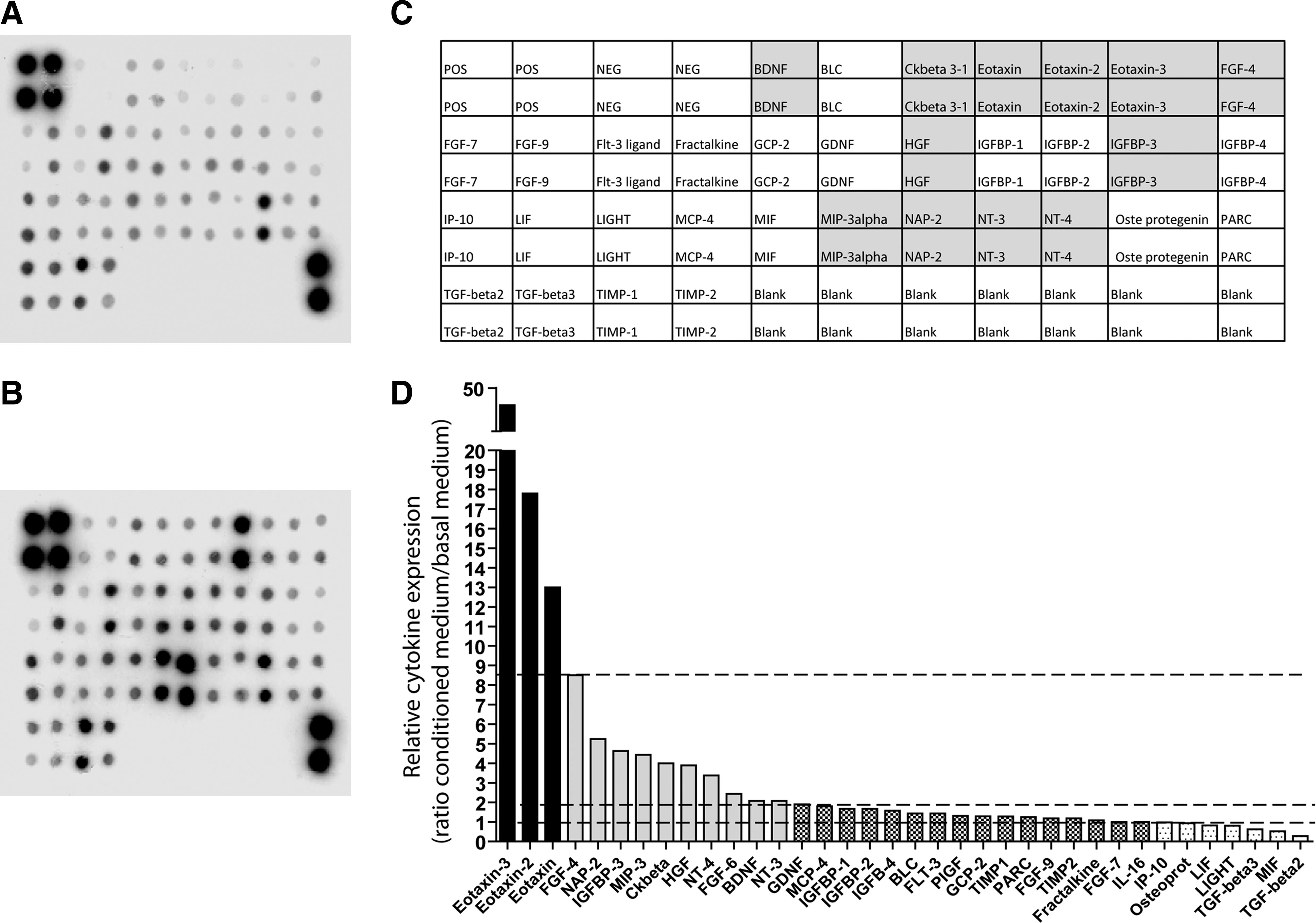
Representative blots showing the profile of expression of several cytokines in standard basal culture medium [DMEM+FBS 10%] (
CNS, Central nervous system; ALS, amyotrophic lateral sclerosis; MHC-II, major histocompatibility complex class II; CFS, cerebrospinal fluid; ND, not determined; FGF-4, fibroblast growth factor-4; NAP-2, neutrophil-activating peptide 2; IGFBP-2, insulin-like growth factor binding protein-2; IGF, insulin-like growth factor; MIP-2, macrophage inflammatory protein-2; NF-κB, nuclear factor κB; CREB, cAMP response element-binding; HGF, hepatocyte growth factor; SVZ, subventricular zone; NT-4, neurotrophin-4; FGF-6, fibroblast growth factor-6; BDNF, brain-derived neurotrophic factor; NT-3, neurotrophin-3.
In vivo characterization
HuCB-MNC tracking—SOD1G93A mice
One month after transplantation, Hoechst 33258-positive cells were mainly detected in the choroid plexus and in the wall of lateral ventricles of SOD1G93A mice (Fig. 2a). In only two cases were few cells found in the white matter; no Hoechst 33258-positive nuclei were observed in the grey matter of the spinal cord. Three months after transplantation, a relatively large amount of Hoechst 33258-positive nuclei were found in the walls of the lateral ventricles and in the choroid plexus (Fig. 2b) of all mice analyzed.

(
Although a progressive loss of fluorescence was observed in 22-week-old mice (12 weeks after transplantation) in comparison with the 14-week-old mice (4 weeks after transplantation), an appreciable number of Hoechst 33258-positive cells were still observed in the choroid plexus of mice at the end stage of the disease (Fig. 2c). Higher magnified pictures of Hoechst 33258-positive nuclei 4 weeks (Fig. 2a′) and 12 weeks (Fig. 2b′) after transplantation or corresponding to the end stage (Fig. 2c′) reveal an overall loss of fluorescence over the time.
HuCB-MNC tracking—wobbler mice
One week after HuCB-MNC transplantation, a large number of Hoechst 33258-positive cells were present in the ventricular system of the brain. These cells were almost exclusively confined in the inferior horn of the lateral ventricle and associated with the choroid plexus (Fig. 2d). Many Hoechst 33258-positive nuclei were also observed 6 weeks after HuCB-MNC transplantation with a similar pattern of distribution. They were mainly localized in the lateral ventricles layer close to the site of administration (Fig. 2e). An appreciable amount of fluorescent nuclei was also found in the posterior part of lateral ventricle, in the third ventricle, and in the brain parenchyma (Fig. 2f). Sixteen weeks after HuCB-MNC transplantation, the fluorescent nuclei were drastically reduced; few positive clusters were observed almost exclusively in the choroid plexus of lateral ventricles close the injection site (Fig. 2g).
Few Hoechst 33258-positive nuclei were observed along the needle track into the cerebral cortex (dotted lines) above the lateral ventricles 1 week after HuCB-MNC transplantation (Fig. 2h). The number and intensity of fluorescence was remarkably decreased 5 weeks later (Fig. 2i) and almost completely disappeared 16 weeks after cells transplantation (Fig. 2j).
Immunofluorescence
In both SOD1G93A and wobbler mice, only very few cells with Hoechst 33258-positive nuclei expressed human-nestin (Fig. 2k–m) and human-GFAP (Fig. 2l–m) in the brain of HuCB-MNC transplanted mice. No co-localization was found with human β-tubulin III. The co-localization experiment between Hoechst 33258 (Fig. 2n) and HLA-I (Fig. 2o–p) showed that 1 month after transplantation Hoechst-positive nuclei were still confined to the ventricular system and most of them were immunoreactive for the human endogenous marker HLA-I.
Effect of HuCB-MNCs on disease progression
SOD1G93A mice
The progressive decrease in body weight of SOD1G93A mice was not significantly modified (p>0.05) by HuCB-MNC treatment although the slope of the curve appeared less severe in treated mice (Fig. 3a).

(
Impairment of motor functions, revealed by stride length and RotaRod tests, in PBS-treated SOD1G93A transgenic mice started at about 21–22 weeks of age and progressed rapidly until death at about 25–26 weeks age. Treatment with HuCB-MNCs significantly slowed the reduction of stride length (p<0.05) (Fig. 3b) and remarkably delayed the deficit in the rotarod performance (p<0.05) (Fig. 3c) compared to PBS-treated mice. The life span of SOD1G93A mice treated with HuCB-MNCs (198±7, mean days±SD) was significantly longer in comparison with vehicle-treated mice (180±10, mean days±SD; p<0.01; Gehan–Breslow–Wilcoxon test; Fig. 3d
Wobbler mice
The rapid and progressive alteration of forepaw adduction observed in PBS-treated wobbler mice was greatly attenuated by HuCB-MNC transplantation (Fig. 4a). The positive effect of HuCB-MNC transplantation was confirmed also by the running speed and grip strength trials (Fig. 4b–c). The improvement was already observed at the second week after transplantation and was maintained until the end of behavioral observation (20 weeks). In addition, wobbler mice transplanted with HuCB-MNCs showed a significant increase of body weight from the 15- to the 20-week age (Fig. 4d). This difference was likely due to better well-being in wobbler mice receiving HuCB-MNCs rather than a direct effect of a reduced muscle atrophy.
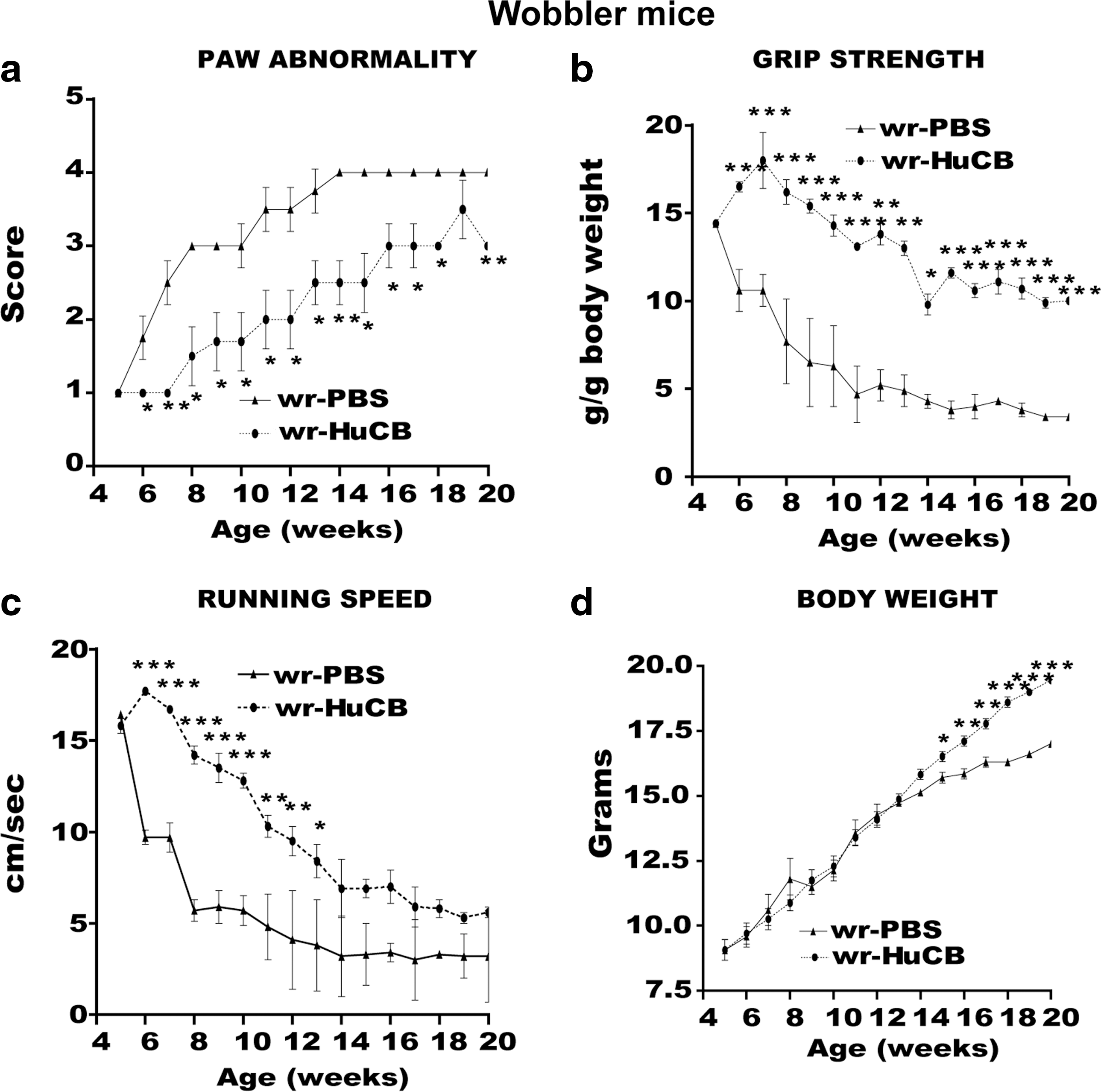
(
Histopathology
Figure 5, a–f, shows the immunohistochemistry of ChAT-positive motor neurons in lumbar spinal cord of 22-week-old SOD1G93A mice treated with PBS (Fig. 5b–e) or HuCB-MNCs (Fig. 5c–f) compared to nontransgenic mice (Fig. 5a–d). A reduction of about 70% or 66% was found in the lumbar spinal cord of PBS- (6.92±0.73) and HuCB-MCN- (7.82±2.69) treated mice, respectively, as compared to nontransgenic mice (23.20±0.73). We also examined the cervical region of spinal cord and found a decrease of about 50% of motor neurons in SOD1G93A PBS-treated mice (13.07±4.40), in comparison to nontransgenic mice (27.40±1.42). No differences were observed in the number of motor neurons of the cervical spinal cord from SOD1G93A mice treated with HuCB-MCNs (9.1±1.63).
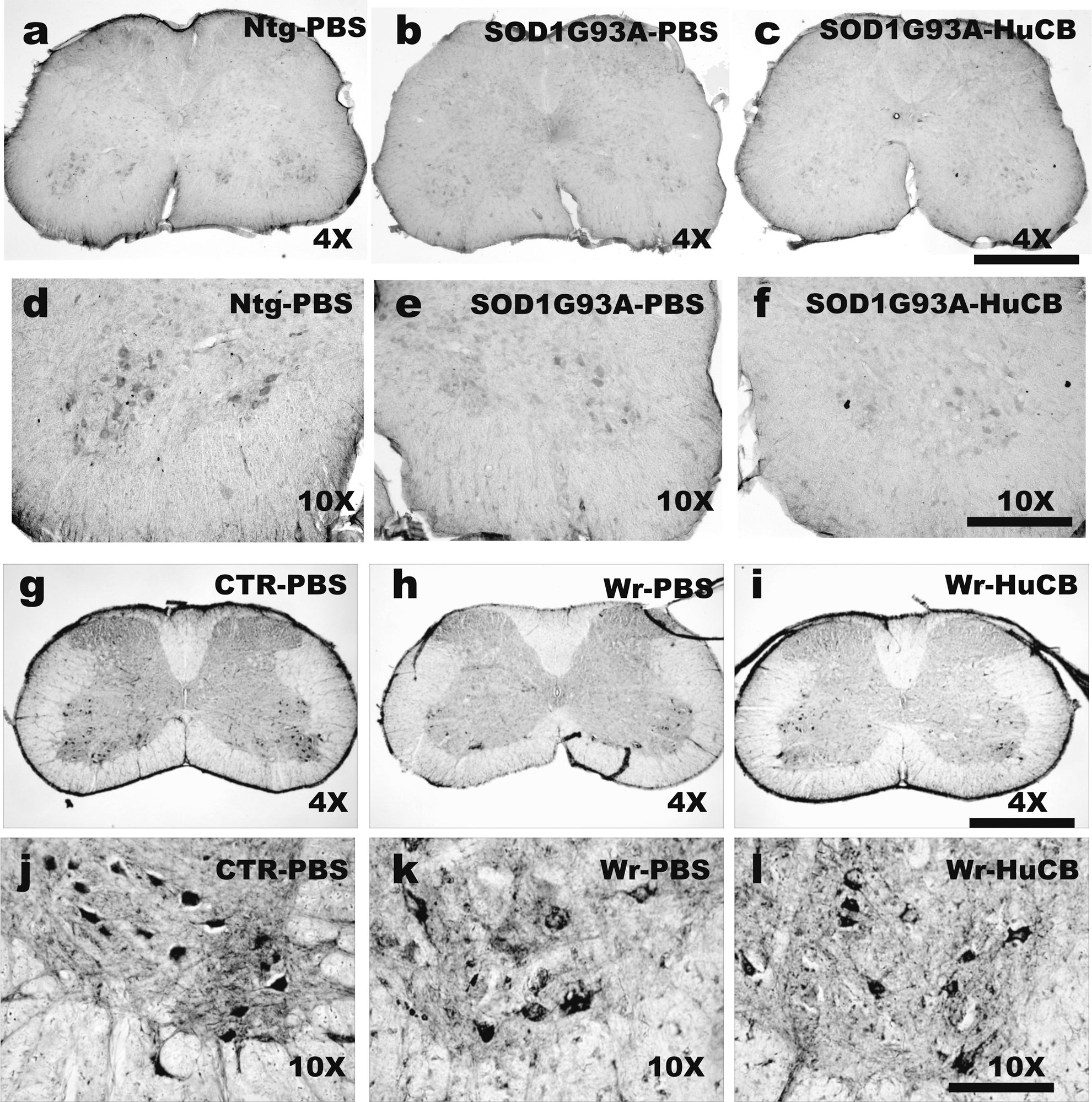
(
The mean number of ChAT-positive cervical motor neurons in 10-week-old PBS-treated wobbler mice (Fig. 5h–k) was about 66% lower compared to healthy littermates (Fig. 5g–j) (control [CTR] group 33.6±1.2; PBS-treated group 7.9±1.3, p<0.0001). The mean number of cervical motor neurons was significantly higher in 10-week-old wobbler mice receiving HuCB-MNCs (Fig. 5i–l) compared to PBS-treated mice (HuCB-MNC group 15.1±0.6; PBS group 7.9±1.3, p<0.001). However, the motor neurons in HuCB-MNC-treated wobbler mice still remained about 50% lower compared to healthy littermates. The biceps weight, an indirect measurement of the degree of atrophy, was significantly reduced in PBS-treated wobbler mice compared to healthy littermates (vehicle-treated wobbler mice, mg: 3.4±0.4; healthy littermates, 18.3±2.9, p<0.001). This effect was partially, but significantly, counteracted by HuCB-MNC treatment (HuCB-MNC-treated wobbler mice, mg: 6.1±0.6; p<0.01). However, their weight remained three-fold lower than that of the NFR mice. The present result suggests that HuCB-MNCs only marginally reduced the motor neuron loss and muscle atrophy in wobbler mice.
Discussion
Systemic injection of a very high concentration of HuCB stem cells into SOD1G93A mice exhibited a beneficial effect regarding the clinical progression and survival, even if only few HuCB-MNCs were found in the brain and no replacement of degenerating motor neurons was reported. 11,13 To enhance the effect of HuCB in the CNS by using a lower number of cells and avoiding surgical problems related to the direct graft into the spinal cord, ICV transplantation was carried out in two different models of motor neuron degeneration. This route of administration has been already successfully exploited for vascular endothelial growth factor (VEGF) treatment in rat models of ALS. 23
To allow the tracking of transplanted cells in the CNS, a vital nuclear intercalant (Hoechst 33258) was used to label the cells before the transplantation. This labeling strategy did not influence the survival of HuCB-MNCs in vitro. In both SOD1G93A and wobbler mouse models, a relevant amount of Hoechst 33258-positive cells remained confined to the ventricular system for the entire duration of the disease, and only very few cells were detected in the parenchyma. A selective localization of HuCB-MNCs was found exclusively associated with the choroid plexus of lateral ventricles, even 4 months after transplantation. It is possible that the adhesion of HuCB-MNCs to mesodermal tissues (like the endothelial layer of ventricles and choroid plexus) may favor their long survival. 24 Only very few labeled cells in the ventricles expressed the neural progenitor markers, such as nestin and GFAP, suggesting that CNS environmental milieu did not influence their differentiation toward neuro-glial–like phenotype. This is in line with observations from Garbuzova et al. using systemic injection of HuCB-MNCs. 11 Very small clusters of fluorescent nuclei were localized in the dorsal white matter of the spinal cord, close to the meningeal layer, in both SOD1G93A (lumbar and cervical region) and wobbler mice (cervical region). No labeled cells were found in the ventral horn of the spinal cord, thus indicating a lack of migration of the HuCB-MNCs toward the area of degeneration. Even in the wobbler mice, where a slight protection of motor neurons was found, no Hoechst 33258-positive cells were observed in the cervical spinal cord.
Thus, while in wobbler mice the behavioral improvement might be partially associated with a modest rescue of cervical spinal motor neurons, the lack of motor neuron protection in SOD1G93A mice suggests that other mechanisms could contribute to the delayed disease progression and increased survival of these mice. It has been reported that peripheral mechanisms at the nerve and muscle level significantly contribute to the pathogenesis of the disease in SOD1G93A mice. 25,26 A modulation of these mechanisms by HuCB-MNCs might favor a delay in the disease progression, even in absence of motor neuron protection. In fact, a recent study has demonstrated that HuCB-MNCs significantly improved the probability of neuromuscular transmission in muscles from SOD1(G93A) mice. 27
Therefore, the beneficial effect emerging from this study may be explained by the effect of factors released by transplanted cells (bystander effect) such as a variety of growth factors, cytokines, and chemokines that could support the functionality of the remaining motor neurons and/or acting at the peripheral districts involved in the disease, like nerves and muscles. 28 HucB-MNCs are known to produce and release several neurotrophic factors, cytokines, and chemokines with potential protective effect in neurodegenerative disorders, when cultured from 10 to 28 days after plating. 29,30 Because the behavioral improvement already occurred 1 week after HuCB-MNC transplantation in wobbler mice, we decided to evaluate a panel of released cytokines and growth factors in vitro 2 days after plating. Among the several cytokines/chemokines detected in the medium, the highest levels were associated with the family of eotaxins (also known as CCL11, CCL13, and CCL24). Eotaxins belong to the intercrine beta (chemokine CC) family. Their main activity is the recruitment and the activation of eosinophils and basophils in response to the release of histamine and leukotriene and the modulation of the inflammatory state during the first phases of allergic response by playing a relevant role through a Th2-associated response. 31,32 Interestingly, high mRNA levels of CCL24 and CCL11 (9.5-fold, 3.9-fold, respectively) were found in laser-dissected facial motor nuclei at the early phase after axotomy in both wild-type and SOD1G93A mice in association with the overexpression of antiinflammatory cytokines. 33 On the contrary, they decreased when proinflammatory response was activated and motor neuron degeneration occurred. 33 In addition, a significant increase of eotaxin (CCL11) levels was reported in the serum of ALS patients compared to noninflammatory neurological patients. 34
Therefore, we speculate that the high levels of CCL11 secreted by HuCB-MNCs might induce a Th2 antiinflammatory response in the injured regions both in the CNS and in peripheral districts like nerves and muscles, supporting previous evidence of an association between the presence of Th2 cytokines and the increased life span of SOD1G3A mice after systemic injection of HuCB-MNCs. 11,35 This is in line with the hypothesis that the activation of an adaptive immune response both in the central and peripheral nervous system has beneficial effects in the mouse models of MND 36,37 and may provide opportunities for therapeutic intervention in ALS. 38
Another important HuCB-MNC-secreted cytokine with potential beneficial effect in ALS is HGF. HGF has been shown to delay ALS progression in both SOD1 mutant rats and mice models 39,40 by a bifunctional action, i.e., direct neurotrophic effect on motor neurons that express its specific receptor MET 41 and indirect antiinflammatory activity. 39 In particular, HGF has been reported to reduce gliosis and attenuate microglia activation in damaged regions through the inhibition of monocyte chemoattractant protein-1 (MCP-1). 39 MCP-1 plays a relevant role in the Th1-mediated proinflammatory response by the recruitment of monocytes, macrophages, and microglia to sites of injury and infection. An early increase of MCP-1 mRNA levels was observed in the spinal cord of presymptomatic SOD1G93A mice, 42 and this effect markedly increased with the progression of the disease. In addition, elevated expression of MCP1 has been found in both the plasma and cerebrospinal fluid (CSF) of ALS patients. Interestingly, high levels of MCP-1 in CSF correlated with a rapidly progressing disease, suggesting that a reduction of this chemokine could have an effect in the delay the disease progression. 42
Overall these data suggest that the release of eotaxins and HGF from transplanted HuCB-MNCs, in particular at the first phases of the disease progression, may favor the maintenance of a Th2 innate adaptive immune response delaying the switch to the Th1-mediated proinflammatory response and therefore attenuating the clinical progression in both SOD1G93A and wobbler mice. This view is in line with the hypothesis formulated in a recent review by Appel et al. 38
Another interesting factor released from HuCB-MNCs, which may contribute to the amelioration of the disease progression of MND mouse models, is FGF-6. FGF-6 administration has been shown to improve muscular function after sciatic nerve transection in rats by favoring the muscle reinnervation independent of the neuron source used for reinnervation. 43 This may account for the improvement of motor performance, observed in both HuCB-MNC-transplanted SOD1G93A and wobbler mice, despite of the lack of or the slight effect on motor neuron protection.
In addition, other neural factors released from HuCB-MNCs, such as NT-3, NT-4, and BDNF, that are able to enhance neuronal survival 44 and retard motor dysfunction as well motor axon loss in wobbler mice 45 may also contribute to the overall beneficial effect of HuCB-MNCs. In this respect the use of HuCB-MNCs may represent a reliable therapeutic strategy for a combination of protective factors. Nevertheless, it must be considered that these cells incubated over long period, i.e., 28 days, are also capable of producing proinflammatory cytokines, such as tumor necrosis factor-α (TNF-α, IL-1, IL-6, and MCP-1) that may conflict and/or reduce the beneficial effect according the study by Newman et al. 30
This might account for the modest effect in delaying the disease progression, even if transplanted HuCB-MNCs remained in ventricular layers during the whole duration of the experimental study. Hence, in agreement with the suggestion by Garbuzova et al., 11 a better approach could be the multiple administration of fresh HuCB-MNCs during the disease progression with the aim to maintain highest the secretion of antiinflammatory and protective factors in respect to those that are potentially toxic.
Conclusions
The present study confirms the protective role of HuCB-MNCs in attenuating symptom progression in SOD1G93A transgenic mice and provides the first evidence for the beneficial effect of this cell therapy in the wobbler mouse model of motor neuron degeneration associated with a mutation in a protein involved in vesicular transport and cell trafficking. The evidence that HuCB-MNCs have a protective role in murine models of motor neuron degeneration characterized by different gene mutations and functional expression of the mutated protein greatly extends the spectrum of the potential application of HuCB-MNCs toward SOD1 mutation–unrelated ALS forms. Another important aspect highlighted by the present study is that despite the cells' transplantation directly into the CNS the benefit obtained was similar to that of previous studies with the intravenous injection of HuCB-MNCs 11 (mean 10% increase of SOD1G93A life span in both studies). Importantly, in both MND models, HuCB-MNCs do not migrate toward affected spinal cord region(s), thus confirming the hypothesis that HuCB-MNCs act as source of protective factors that limit the propagation of damage rather than substituting injured cells, as also demonstrated in a murine model of myocardal infarction. 46
Finally, because many antiinflammatory chemokines/cytokines are potentially secreted by HuCB-MNCs either in the CSF or in the periphery, the present study further corroborates the hypothesis of a neuroprotective immunomodulation as a potential strategy for treating this devastating disease.
Footnotes
Acknowledgments
This study was supported by grants from the Italian Ministry of Health (Progetto Ricerca Finalizzata 2005; Progetto a concorso 2006 and 2007, Ex Art. 56), the Italian Ministry of Health (Malattie Neurodegenerative, ex Art.56, n.533F/N1), Foundation NovusSanguis and Foundation Jerome Lejeune, the 6FP EU Project–THERCORD, Thierry Latran Foundation (France). V.S., L.C., and C.C. were supported financially in part by the Italian Ministry of Health (Malattie Neurodegenerative, ex Art.56, n.533F/N1). We thank Dr. Mauro Tettamanti and collaborators of the Mario Negri Institute for their support in the statistical analysis of data.
Author Disclosure Statement
No competing financial interests exist.