
Abstract
BALB/c mice were divided into young, middle-aged, and aged groups, and each group was given 3 weeks of oral treatments: (1) 1 mL of VBC1-99 (a mixture of 42 fruits and vegetables extracts) or (2) 1 mL of same amount of antioxidant vitamins as control. Steady-state hepatic adenosine triphosphate (ATP) was assessed by phosphorus-31 nuclear magnetic resonance (31P-NMR) spectroscopy as: β-ATP/reference peak, inorganic phosphorus (Pi)/peak and β-ATP/Pi. As compared to untreated control, VBC1-99 significantly enhanced β-ATP/peak and β-ATP/Pi ratios (p<0.01) in all age groups and throughout the observation period (p<0.05) together with a significant decrease of Pi/ref peak ratio (p<0.05). However, this value in middle-aged and aged mice was comparable to antioxidant control mice. These NMR data demonstrate that VBC1-99 has a beneficial effect on hepatic energy metabolism, irrespective of age.
Introduction
Ames et al. 1 have suggested that an optimum intake of micronutrients could tune up metabolism and give a marked increase in health, particularly for the elderly, obese. Under conditions of increased ATP demand, liver mitochondria are able to increase ATP synthesis up to 2.8 times by switching more mitochondria from the resting state 4 to active state 3. However, aged hepatocytes, with a 30% complex I activity reduction, increase ATP production only up to 1.9 times. 2 Nonetheless, higher demands in younger subjects together with possible concurrent subtle micronutrients deficiencies may represent a further unsuspected health risk. Therefore, we examined steady-state energy status change in mouse liver of different age during 3 weeks treatment with VBC1-99 (a mixture of 42 extracts of fruits and vegetables), as compared to antioxidant-treated and untreated controls. As high-energy triphosphate compounds, we tested ATP using phosphorus-31 nuclear magnetic resonance (31P-NMR) spectroscopy, which allows noninvasive direct measurements of metabolite concentrations and tracing of metabolic fluxes in situ in experimental animals and human livers. 3
Materials and Methods
Animals
A total of 120 healthy BALB/c mice raised in a controlled vivarium were divided into three separate age groups (40 mice each): Young (1 month), middle-aged (12 months), and aged (24 months). All mice were given water ad libitum and standard chow. Mice were given either VBC1-99 extract (1 mL containing 400 mg of carbohydrates, 0.04 mg of protein, 0.04 mg of fat, 8 mg of vitamin C, 1.2 mg of vitamin E, 0.5 mg of coenzyme Q10, 6 mg of royal jelly, 5 mg of
31P-NMR spectroscopy analysis protocol
Steady-state hepatic ATP was assessed by 31P-NMR spectroscopy before treatment and at weekly intervals. Mìce fasted for 6 hr prior to NMR examination; they were then anesthetized with pentobarbital (50 mg/kg intraperitoneal injection) and immobilized on a shielding belt, which was embedded with paramagnetic beads, to minimize interference from abdominal muscle mass. 31P-NMR spectroscopy was performed at controlled room temperature (23°±1° C, 40%–60% of humidity). Normal ventilation of fresh air (1.0 L/min) through a face mask was also assured during the test. The homogeneity of the magnetic field was optimized by adjusting the shim coils while observing the free induction decay of hepatic water protons. The skin covering the liver side of the mice was attached firmly to the Faraday shielding plate and faced the center of the surface coil. A capillary tube (0.2 mm diameter; 30 mm length) was placed on the reverse side of the coil, filled with 40 mM of methylediphosphonic acid (MDPA), dissolved in dimethylsulfoxide (DMSO), and sealed at both ends. This was also used as reference standard (Ref:Peak) when calibrating the various NMR peaks of metabolic phosphates. The following parameters were adopted: 180° pulse located at the surface, 0.7-sec sampling interval, 800 times accumulation, 8 K sampling point, and 58 ppm spectral width. 4 A 40-Hz line broadening factor was added to increase the ratio of signal to noise, and Fourier transformation was then carried out.
The 31P-NMR liver variables considered were: β-ATP/ref peak, inorganic phosphorus (Pi)/ref peak, and β-ATP/Pi among the wider spectrum, which showed several phosphate peaks, including phosphodiesters, phosphomonoesters, phosphocreatine, γ-ATP, and α-ATP, the latter easily overlapping with ADP. Liver pH was determined by using the following formula: cellular pH=6.75+log (Δδ−10.85)/(13.25−δδ) where δδ is associated with chemical-shift differences of Pi and α-ATP for each examination. 5
Statistical analysis
All data were obtained from three measurements and expressed as means±standard deviations. Statistical analysis was done using Stat View J-5.0 program on a Macintosh computer. Significance was determined by one-way analysis of variance (ANOVA) with Bonferroni correction as a post hoc analysis. A significance level of p<0.05 was used for all the comparisons.
Results
Mice in all groups remained healthy throughout the whole study period, and no adverse effect was noted nor any growth rate abnormality in the young mice.
Effects of VBC1-99 on hepatic pH
No liver pH (calculated as chemical shift difference of Pi and α-ATP) changes were recorded irrespective of the age and treatment group (data not shown).
Time-course study of hepatic energy status
Effects of VBC1-99 on β-ATP
Hepatic β-ATP, expressed as a ratio of hepatic β-ATP:Ref, was similar in all age groups at baseline (3.2±0.3; Fig 1). After VBC1-99 treatment, hepatic β-ATP increased significantly by 19%–25% at 1 week of observation in young and middle-aged groups (p<0.01 vs. AC and UC groups). Aged mice showed a significant increase in this parameter, but starting from the second week of observation (p<0.001 vs. AC and UC groups). At 2 and 3 weeks of observation, all treated groups showed a comparable and significant increase of this parameter.
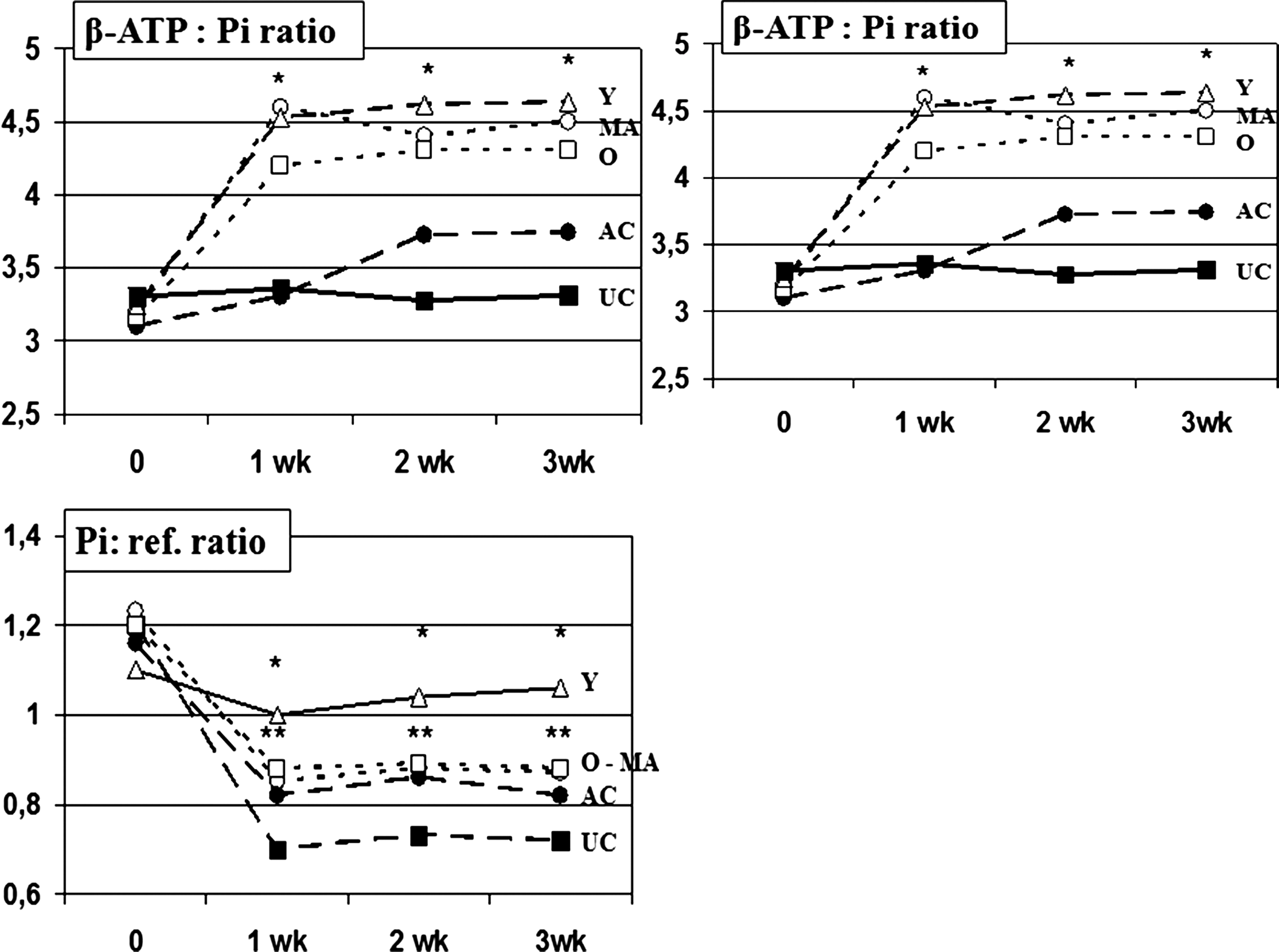
Time-course study of hepatic energy status: Effects of VBC1-99 on β-ATP, hepatic Pi, and β-ATP:Pi ratio. (*) p<0.001 vs. antioxidant control (AC) and untreated control (UC) groups; (**) p<0.05 vs. UC group. ATP, Adenosine triphosphate; Pi, inorganic phosphorus; ref., reference.
Effects of VBC1-99 on hepatic Pi
This was derived from the NMR spectrum and expressed as a ratio to the reference peak and found to be comparable at baseline testing in all age groups. VBC1-99 treatment led to significant decrease of the Pi/ref peak ratio in all age groups (p<0.001 as compared to baseline and UC; Fig 1). However, the value of the Pi/ref peak ratio observed in middle-aged and aged mice was comparable to AC mice, whereas young mice showed a significant difference (p<0.05 vs. AC mice and other age-groups).
Effects of VBC1-99 on β-ATP:Pi ratio
The ratio of hepatic β-ATP:Pi represents the ratio of product-to-substrate for high-energy triphosphates, and the baseline value was similar in the three age groups. The ratio increased by 28%–36% (p<0.001) at 7 days of treatment with VBC1-99 (p<0.001 vs. control).
Discussion
With this study, we have shown that hepatic β-ATP increases in mice treated with 1 mL/day of VBC1-99, unlike with the antioxidant control. We also found that increases in steady-state hepatic β-ATP and decreases in Pi after VBC1-99 treatment enabled a significant improvement of the hepatic β-ATP:Pi ratio. On the other hand, VBC1-99 proved to be very safe for hepatocytes physiology given also that there was no change in hepatic tissue pH. Changes in Pi are likely to be greater than changes in β-ATP synthesis because Pi is used for synthesizing several high-energy triphosphate compounds, including α-, β-, and γ-ATP. Indeed, in our study, we observed that the decrement in Pi was not proportional to β-ATP increases. Because of the overlap of the peaks associated with α- and β-ATP with that of ADP in the 31P-NMR spectrum, actual total changes in ATP and other high-energy phosphate components may warrant further investigations that our study could not address. β-ATP was taken as representative of these compounds, although changes in β-ATP are thus likely to underestimate the actual response of these compounds to VBC1-99 treatment.
Nevertheless, our findings of increased β-ATP/decreased Pi phenomenon were quite significant, indicating a definite steady-state energy enhancement. These findings may be the result of enhanced synthesis of triphosphate compounds and/or reduced production of ATP. Indeed, a reduced oxidative stress may enhance energy metabolism in the mitochondria and facilitate the efficient use of an oxygen supply. On the contrary, it has been shown that patients with type 2 diabetes have markedly lower hepatic ATP production than nondiabetic individuals of comparable age and body mass, 6,7 and the reduction in flux through ATP synthesis depends primarily on the lower hepatic PI and ATP concentrations. 8 The role of each compound of VBC1-99 in enhancing energy metabolism awaits elucidation, but the lack of response with an antioxidant cocktail suggests an advantageous bioavailability of VBC1-99 bioactive moieties.
The VBC1-99-induced improvement in hepatic energy status may also be applicable to other tissue/organs, such as myocardium, skeletal muscles, and brain. It can be hypothesized that under the conditions of rapid oxygen consumption, such as during incremental exercise or high-altitude hypoxic environment, the higher production demands of high-energy triphosphates may be better met after VBC1-99 preconditioning. This holds some interest if we consider that moderate-to-long treadmill training is required to increase liver ATP. 9 Aging intervention may be a further interesting area when considering that the aging liver may be more susceptible to steady-state energy status decay, 9,10 and liver disease with its gradual loss of viable hepatocytes represents a clear pathological example. 11,12
These findings suggest that VBC1-99 is able to markedly enhance the hepatic production of high-energy triphosphates with higher steady-state energy status. These findings are likely to be extended to overall better aerobic capacity either in high-metabolism turnover conditions observed in youth and in decreased gas exchange anaerobic threshold situation taking place in the elderly. With due caution for the experimental setting, these data hold some interest when considering the recent report of the inefficacy of oral ATP administration (5 g/day for 28 days) in healthy subjects. 13
Footnotes
Author Disclosure Statement
The authors have no financial conflicts to disclose.