
Abstract
We have investigated the effects of brief, non-specific deuteration of Drosophila melanogaster by including varying percentages of 2H (D) in the H2O used in the food mix consumed during initial development. Up to 22.5% deuterium oxide (D2O) in H2O was administered, with the result that a low percentage of D2O in the water increased mean life span, whereas the highest percentage used (22.5%) reduced life span. After the one-time treatment period, adult flies were maintained ad libitum with food of normal isotopic distribution. At low deuterium levels, where life span extension was observed, there was no observed change in fecundity. Dead flies were assayed for deuterium incorporation by complete hydrolysis in hot 12 N HCl solution followed by subsequent high-performance liquid chromatography/mass spectrometry (HPLC/MS). Isoleucine and leucine residues showed a small, linear dose-dependent incorporation of deuterium at non-exchangeable sites. Although high levels of D2O itself are toxic for other reasons, higher levels of deuterium incorporation, which can be achieved without toxicity by strategies that avoid direct use of D2O, are clearly worth exploring.
Introduction
Overexpression of superoxide dismutase can indeed extend life span, 10 lending support to the free radical theory of aging, although these findings were later questioned 11 and the issue is still an open question. 12,13 However, the strategy to combat oxidative damage that has widespread adoption, despite equivocal evidence of its efficacy, 14 is to consume “antioxidants” and suppose that these molecules are able to make their way to the sites of ROS production and then promptly quench the ROS before macromolecules are damaged. In this scenario, the oxidized form of the antioxidant itself is presumed to have no deleterious effects. An alternative approach would be to make the DNA and proteins less susceptible to oxidative attack, hopefully without altering their other essential functions. Such a goal may be achieved by isotopic substitution of hydrogen with deuterium, as proposed by Shchepinov. 15,16 A recent follow-on investigation has also demonstrated that genetically defective yeast that have a reduced life span can achieve that of the wild type when cultured in the presence of polyunsaturated fatty acids that are specifically deuterated at reactive sites. 17 Custom synthesis of such molecules is, however, expensive.
We took a simpler approach, using deuterium oxide (D2O) to partially deuterate Drosophila melanogaster larvae during the development period with different molar percentages of deuterium in the water used in the food mix (0%–22.5%). Although D2O, which has a “solvent isotope effect” 18 as well as a kinetic isotope effect, is known to be toxic to D. melanogaster at high levels of enrichment, we found that a one-time treatment followed by normal ad libitum feeding was able to extend mean life span of this wild-type organism, at intermediate levels of deuteration, by almost 9%. We note that previous studies, using 50% D2O on D. melanogaster throughout the life span, and in which temperature was the manipulated variable, found a decrease in life span. 19 Another study 20 found that 20% or 40% D2O slowed the heart rate of D. melanogaster late third-instar larvae, and an additional one found a slowdown in circadian rhythm. 21
Materials and Methods
Administration of deuterium
The short life span, ease of maintenance, and affordability of D. melanogaster made this organism ideal for our study. We used the “IV” population of D. melanogaster, 22 which is large and outbred, and has been maintained on strict 2-week life cycles at∼25°C and constant illumination since 1981. The IV population is adapted to a banana–molasses food medium, which was used for the control group in this study. A preliminary study using deuterium from 0% to 70% revealed appreciable toxicity above the 22.5% dose. Thus, the treatment groups received the same banana–molasses food, but D2O was used in place of H2O to create the following deuteration treatments: 7.5%, 15%, and 22.5%.
Twelve kilograms of sterile 70 mole% D water were obtained in sealed polyethylene containers (Cambridge Isotope Labs, Andover, MA) and assayed by nuclear magnetic resonance (NMR). This “70% D2O” was used to produce various “D” concentrations by diluting with distilled water (which is 0.015 mole% D). Every 0.5 L of food medium contained 17 grams of agar, 150 grams of banana, 18 mL of light karo syrup, 18 mL of dark Karo syrup, 28 mL of barley malt, 53 mL of ethanol, 40 grams of yeast, and 3 grams of p-hydroxybenzoic acid. The control group food was mixed in 1.24 L of water. The 7.5%, 15%, and 22.5% groups had food that was mixed in 1.11 L of water with 0.13 L of 70% D2O, 0.98 L of water with 0.27 L 70% D2O, and 0.84 L of water with 0.40 L 70% D2O, respectively. The percentages used to label the groups (i. e. “7.5%,” “15%,” etc.) were approximations for the mole percent deuterium in only the water used in the food cook. Other ingredients, of course, contain appreciable moisture.
For each treatment, eggs collected from the IV population were placed at densities of∼65–70 eggs per vial into 80 vials containing the appropriately deuterated food. To test for development time, we used similar methods to Burke et al. 23 Emerged flies were removed daily from the vials and sexed under a microscope. On day 11 from egg, 720 flies (half male and half female) from each group were placed in 8-dram vials in groups of 8 (4 male and 4 female). All vials then received standard, un-deuterated, banana–molasses food. Daily mortality was recorded for all vials, and flies were transferred to fresh food every other day. Flies from vials of the same treatment were sometimes combined to maintain equal densities.
On day 12 from egg, 30 freshly emerged female flies were individually placed in vials along with male partners from the same treatment. These flies were given a yeasted-charcoal medium, which allows for eggs to be counted more easily and has been used in previous studies of female fecundity. 24,25 The female flies and their partners were transferred to fresh food daily, and the number of eggs laid by each female was carefully recorded. If the male partner died, another same-aged male from the same treatment group was used as a replacement.
Data analysis
Mean development times, mean longevities, and mean lifetime individual fecundities were compared via analysis of variance (ANOVA) (
Mass spectrometric analysis
After exposure to D2O, a portion of the newly hatched, live flies was collected and separated by dosage group and then stored in a −80°C freezer until analysis. Flies were hydrolyzed in 12 N HCl for 48 hr at 60°C. To determine that there was no significant difference between the deuterium incorporation in male and female flies, 10 groups of 5 flies per sex were segregated by dosage. Each group was placed into a separate vial and, after treatment, a majority of the contents were dissolved and the exoskeletons removed. The hydrochloric acid was passively allowed to evaporate over a 24-hr period, leaving the hydrolyzed residue. During the hydrolytic process, most of the biomolecules present in the flies were destroyed or denatured, except individual amino acids. Leucine and isoleucine occur at high concentrations in the flies and were therefore used as a representative marker for deuterium incorporation. The concentrations of deuterium represented the combination of both amino acids, as they are not separable with the liquid chromatography (LC) method used, nor in the mass spectrometer itself.
The hydrolyzed residue was dissolved in 92.5% acetonitrile/water and analyzed by reversed-phase liquid chromatography positive-ion mode electrospray mass spectrometry using a Micromass LCT mass spectrometer with a Gilson 231 XL autosampler, and LC. The solvent system was: A, 97.8% H2O, 2% acetonitrile, and 0.2% acetic acid, and B, acetonitrile with 0.2% acetic acid. A linear gradient of 98% A to 95% B over 20 min was used. The column was a Phenomenex Luna C18; the injection volume was 10 μL. Mass chromatograms of m/z=132 [M-H]+ for leucine and isoleucine were determined. For these amino acids, the retention time was 1.25 min, so that some non-specific response from the void volume was observed. This response was compensated for by using background subtraction. The extent of deuteration was determined from the excess contribution of the [M Deut +H]+ peak at m/z 133 over the 6.6% that is due to the [13C,M+H]+ peak of the proteo ion (also on m/z 133).
While it would have been informative to compare the extent of deuteration in other types of amino acids, much more extensive sample preparation, including derivatization, would have been required to improve separation and enhance mass spectrometric performance. 27 A check on the viability of using the non-derivatization approach was made using the more labor-intensive phenylisothiocyanate derivatization method for a small subset of hydrolyzed flies. This technique, outlined by Heinrikson and Meredith, 28 resulted in a longer retention time for the amino acids that allowed better separation and much lower non-specific background interference. Comparison of the deuterium incorporation for the derivatized and non-derivatized amino acids showed that the percent deuterium incorporation on leucine and isoleucine was, within experimental error, the same. Therefore, the simpler approach was used for the results presented.
A small subset of the dead flies in each treatment group was also assayed in the manner described above to compare deuterium incorporation at the end of life. The results were nearly identical to those obtained on the newly hatched live flies, indicating that the deuterium incorporated during development is present throughout this organism's life span (data not shown).
Results
Survival curves for control and treatment groups are shown in Figs. 1 (male) and 2 (female). A summary of life span statistics is laid out in Table 1. The data on mean longevity, mean development time, percent viability, and mean fecundity are displayed in Fig. 3. Figure 3 illustrates that the %D slowed the development time for the flies from∼12 days to∼14 days at the highest doses. In all of the pairwise comparisons between the treatment groups, the group with the higher D2O percentage had longer development time, although the difference between the females in the 7.5% and 15% groups was not statistically significant. Viability of eggs was also reduced by the presence of deuterium in the medium (Table 2, Fig. 3), decreasing as the %D increased. Viability was estimated in this case by noting the number of flies produced per vial, and assuming that roughly equal numbers of eggs (65–70) had been initially deposited into each vial. Unlike the fecundity analysis, individual eggs were not tallied.
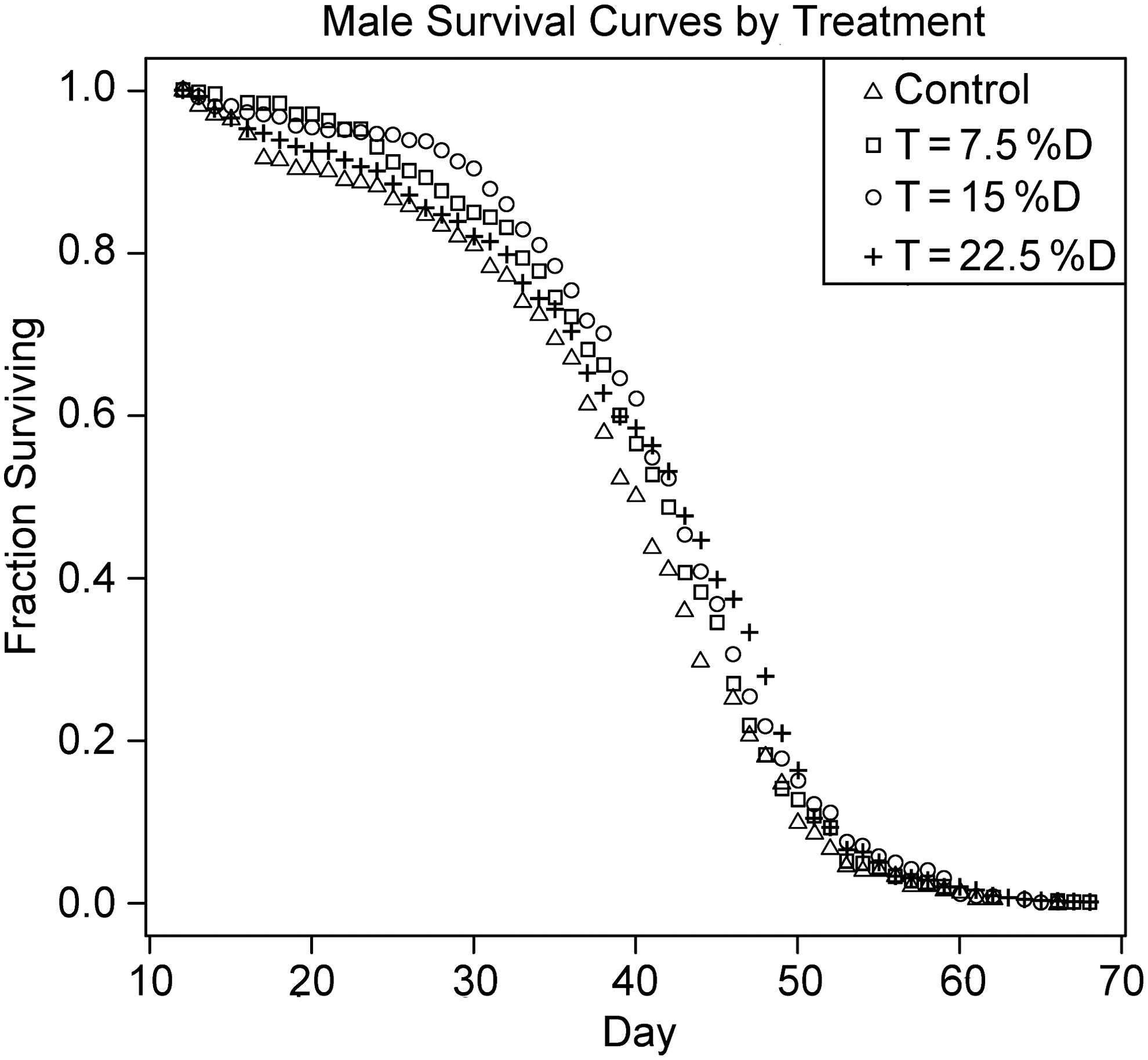
Male survival curves for the control and each of the three treatments.

Female survival curves for the control and each of the three treatments.
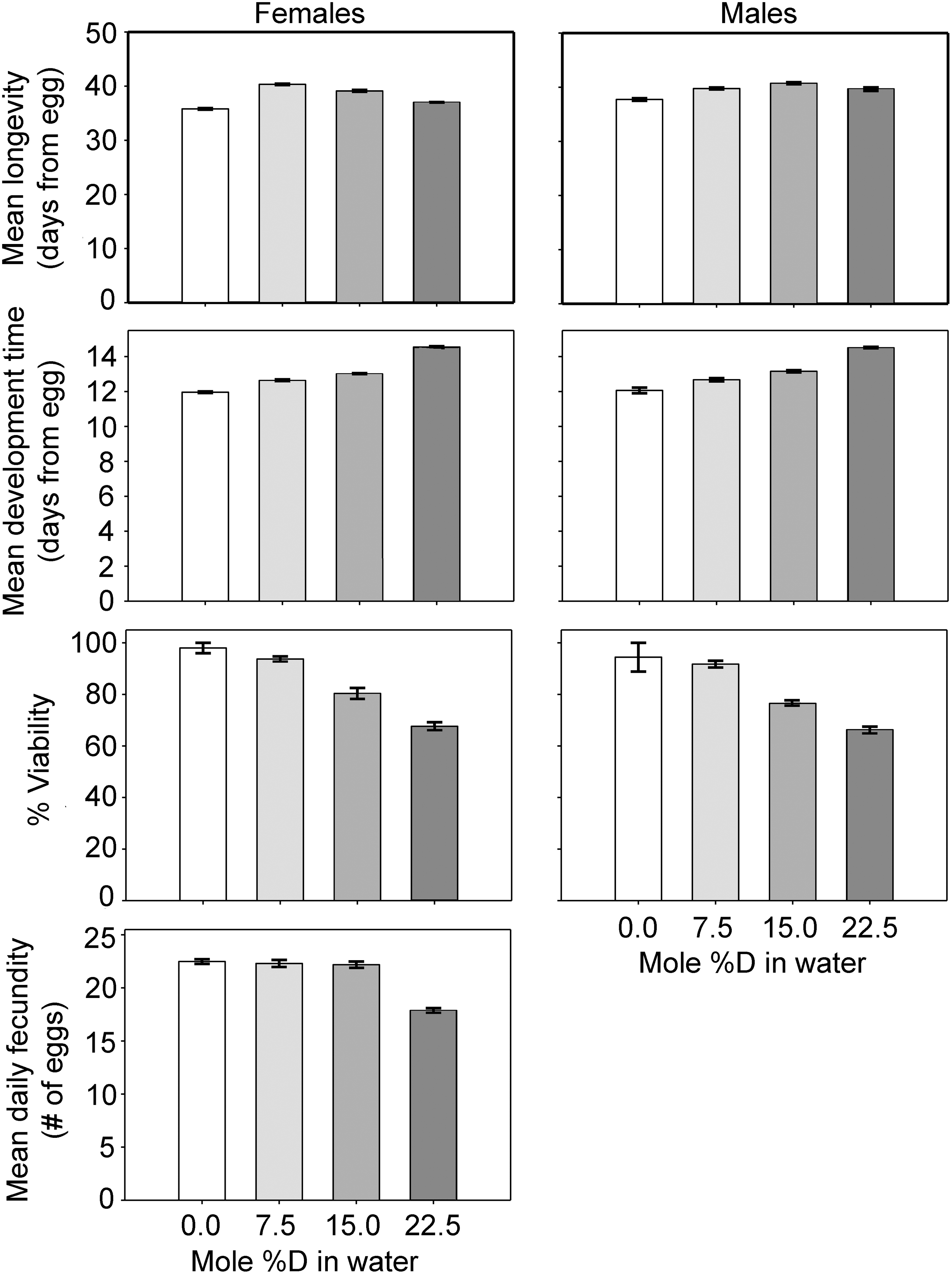
A graphical summary of mean life span, mean development time, percent viability, and fecundity for the control and each of the three treatments. Viability decreased monotonically across the range of treatments. Fecundity decreased monotonically across the range of treatments, but only the decrease at the highest %D was statistically significant. Mean life span increased, with the greatest effect at intermediate levels of %D.
SD, Standard deviation; CI, confidence interval.
NA, Not applicable.
To obtain treatment effects for each experiment, a Cox proportional hazards model was fit separately to male and female mortality data for each (with a single treatment/control covariate). Models in 5 out of the 6 cases showed significant reduction in mortality hazard for treatment versus control at p<0.05 (both Wald and score tests) as shown in Table 3 and Fig. 4. The coefficient for treatment of males in the low-dose (7.5% D) condition was not significant, although the estimate was in line with those obtained in the other conditions. Models fit to males and females jointly in each experiment (with different baseline hazards but a single treatment effect), not shown, also showed significant reduction in mortality hazard in all three experiments (p<0.05, Wald and score tests). As implied by the standard errors in Table 3 and error bars in Fig. 4, we cannot reliably determine the relative magnitudes of effect sizes by sex or dose; however, it is clear that the overall effect is one of reducing mortality hazard for all treatments and groups examined by approximately 10%–30%, with the possible exception of the low-dose male condition. An overall view of the data indicated that the addition of D2O to the food increased the longevity of flies, with intermediate deuterium levels (7.5% and 15%) having more benefit. For females, we also compared fecundity among treatment groups. There were no appreciable fecundity differences among the 0%, 7.5%, and 15% groups, but all of these groups had significantly higher fecundity than the 22.5% group, as indicated in Fig. 3.
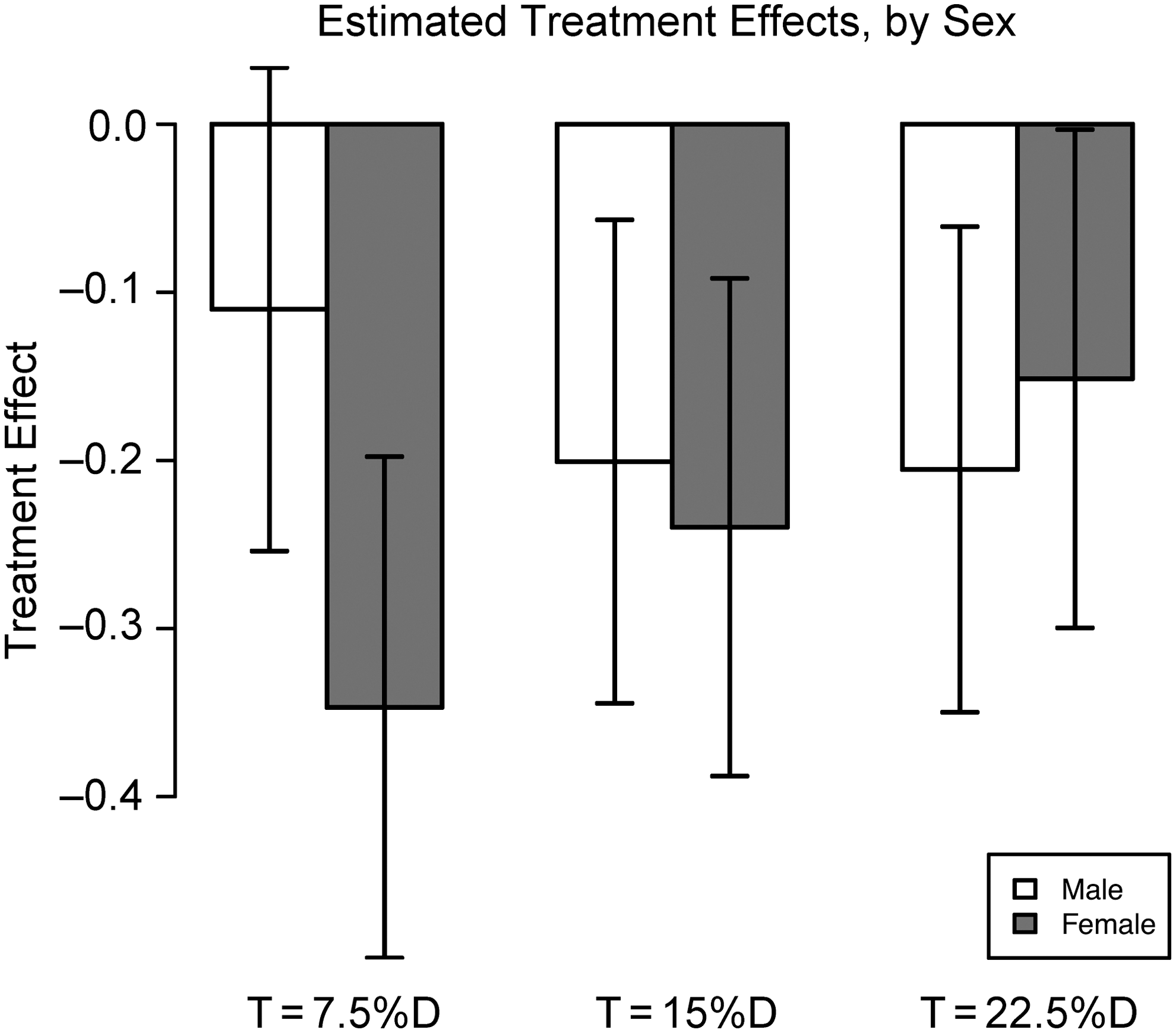
A graphical summary of the Cox proportional hazard analysis. Treatment groups showed a 10%–30% reduction in mortality hazard.
SE, Standard error.
Figures 5 and 6 show dose-dependent incorporation of deuterium atoms into leucine and isoleucine amino acids of female and male flies, as detected by HPLC/MS, plotted versus the %D2O in the water used for the aqueous portion of the food mix. It is clear that D2O administration during development results in some uptake of D into these amino acids, and only non-exchangeable C–D bonds would be seen in this assay. Any deuterated –OH and –NH2 groups, would back-exchange to the protonated form in the HCl(aq) solution that was used to hydrolyze the flies. It is known 29 that both isoleucine and leucine are essential amino acids for adult D. melanogaster, and so incorporation of D into these amino acids is not possible in the flies themselves, there being no pathway for such synthesis. However, yeast produce abundant deuterated amino acids when supplied with D2O, 30,31 can tolerate very high percentages of D2O, and are ubiquitous in the food itself. Thus, it is possible that the deuterium incorporation was via yeast or other bacteria. No effort was made to produce sterile conditions during the experiment. If essential amino acids contain non-exchangeable deuterium, it stands to reason that the other molecules (DNA, RNA, lipids, sugars, etc.) synthesized internally would also experience a similar, although not necessarily identical, ratio of deuterium incorporation.
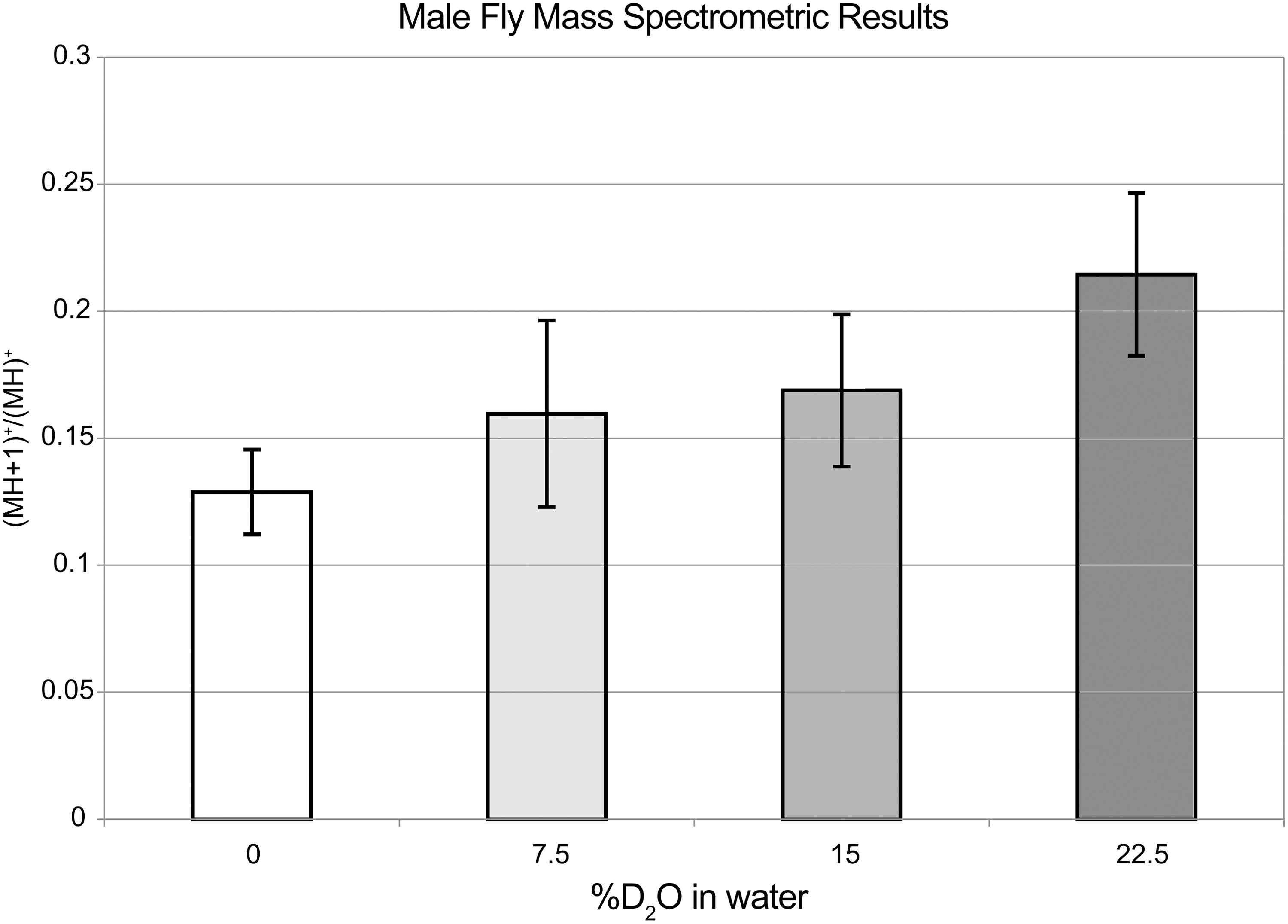
Mass spectrometric results for Leu and Ile amino acids obtained from hydrolyzed male flies, showing the good correlation between the (MH+1)+/MH + intensity ratio and the level of D2O in the aqueous portion of the food mix. A correction has been made for the naturally occurring carbon-13 isotopomers, which also occur on the same mass (see text).
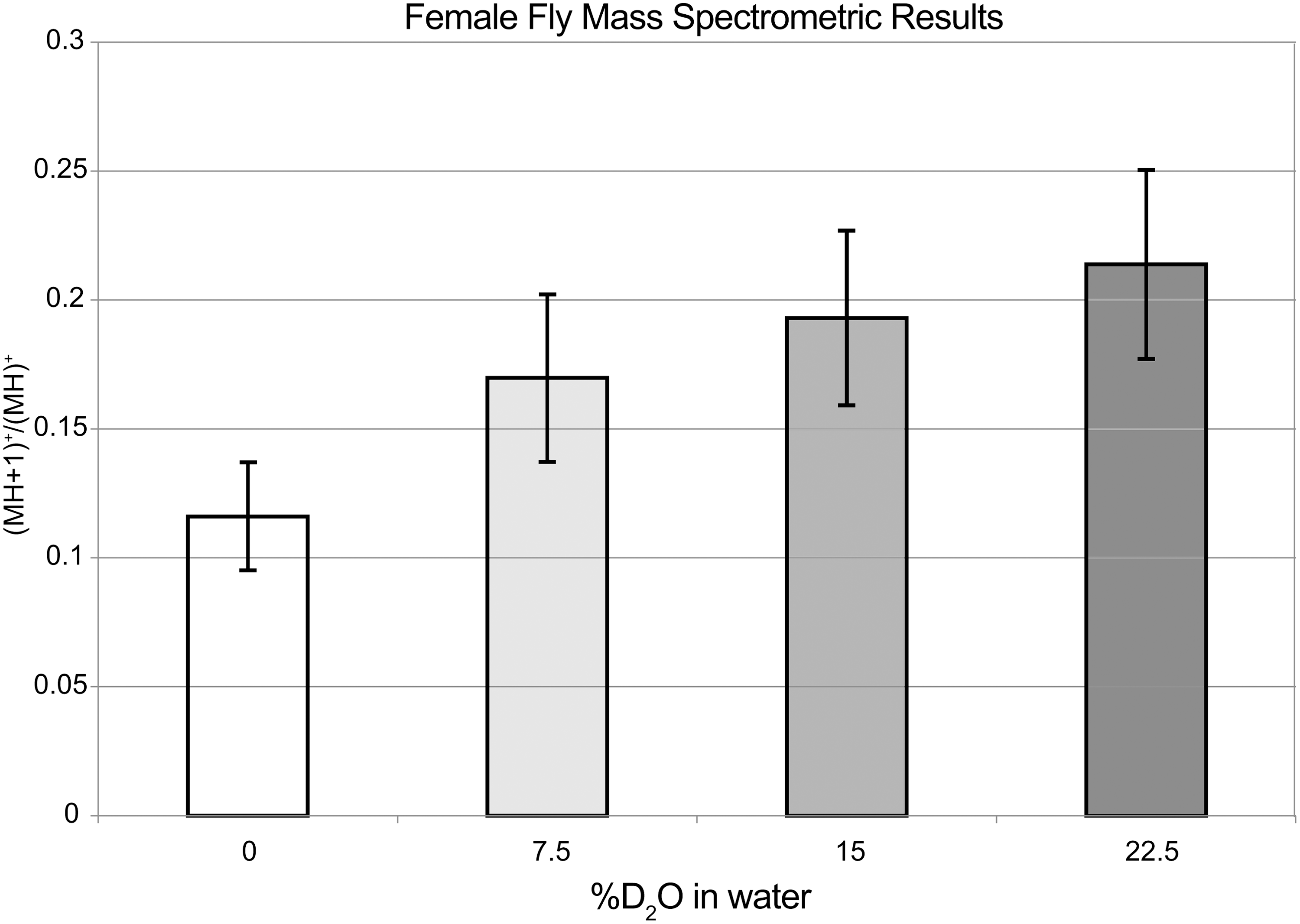
Mass spectrometric results for Leu and Ile amino acids obtained from hydrolyzed female flies, showing the good correlation between the (MH+1)+/MH + intensity ratio and the level of D2O in the aqueous portion of the food mix. A correction has been made for the naturally occurring carbon-13 isotopomers, which also occur on the same mass (see text).
Discussion
Low percentages of D2O added to the regular diet of D. melanogaster improved mean life span, as illustrated in Figs. 1 and 2, without an effect on fecundity. However, D2O had an adverse effect on development time and showed toxicity as the dose was increased. Thus, D2O itself is of limited utility to explore the possibility of greatly curtailing oxidative damage by a high-percentage substitution of D for H in D. melanogaster.
From the standpoint of theories of aging, the present results provide some support for the free-radical mechanism for aging, as the chemical stabilization afforded by deuteration appeared to increase mean longevity, by almost 9%. However, there are several important qualifications that should be borne in mind. First, higher levels of deuteration had adverse effects on development, viability, and female reproduction. Although such effects were limited at the lower levels tested here, it is evident that at the highest level of deuteration we used (22.5%) there can be obvious toxic effects. Second, we have not shown directly that free-radical damage to macro-molecules in the assayed flies was in fact reduced. That is, the observed benefits of low-to-moderate levels of deuteration may have arisen from some other mechanism. Third, even granting that increased chemical stability is the mechanistic basis of the observed benefits of deuteration, our results support the hypothesis that free-radical biochemistry is a part of the mechanistic foundations of aging, rather than its singular cause. This is underscored by the rather modest increase in longevity that we have observed, particularly when compared with the longevity differences that can be produced by laboratory selection 32 starting with the same population of D. melanogaster that we studied in this work.
Footnotes
Acknowledgments
This research was facilitated by a faculty collaboration grant from the Council on Research, Computing and Library Resources (CORCLR) from UCI. S.H. and K.E. were partially supported by the UCI Undergraduate Research Opportunity Program (UROP) and the Summer Undergraduate Research Program (SURP). We thank Dr. John Greaves (UCI Mass Spectrometry Facility) for his expert assistance with mass spectrometry acquisition and data analysis, and Prof. Carter Butts (UCI Sociology Dept.) for expert assistance with the statistical analysis.
Author Disclosure Statement
No competing financial interests exist.