
Abstract
Scopolamine (Sco) can induce amyloid β (Aβ) deposition, oxidative stress, synaptic dysfunction, and learning/memory impairment as observed in Alzheimer's disease (AD), the most common form of dementia affecting more than 25 million elderly people worldwide. Herein we explored the protective effect of 7,8-dihydroxyflavone (7,8-DHF) on Sco-induced Aβ deposition, oxidative stress, synaptic dysfunction, and learning/memory defects. Rats were randomly divided into four groups (n=12 for each group). The control group received normal saline (NS); the Sco group received Sco (1 mg/kg per day) intraperitoneally (i.p.) for 2 weeks. Mice in the Sco+7,8-DHF group received 1 mg/kg per day 7,8-DHF i.p. for 2 weeks, followed by Sco (1 mg/kg per day)+1 mg/kg per day 7,8-DHF (i.p.) for another 2 weeks. The 7,8-DHF group received 1 mg/kg per day 7,8-DHF (i.p.) for 4 weeks. Results showed that the supplement of 7,8-DHF significantly reversed Aβ deposition, oxidative stress, synaptic dysfunction, and cognitive defects. Our data suggest that 7,8-DHF might serve as a promising therapeutic candidate for attenuating Sco-induced AD-like pathological dysfuntion.
Introduction
A
Scopolamine (Sco) is a non-selective muscarinic receptor antagonist that impairs learning and memory by blocking cholinergic signaling and by some indirect mechanisms. 7 Sco-treated rats display impaired spatial learning and memory abilities, increased Aβ deposition, 8,9 activation of oxidative stress, 10 and synaptic dysfunction. 11 It has been widely employed as a pharmacological drug to model AD-related cognitive decline and to screen potential therapeutic target molecules for AD treatment. 8,11 A variety of drugs used for dementia therapy have been screened via this model. 12,13
7,8-Dihydroxyflavone (7,8-DHF) was identified as a novel, high-affinity TrkB agonist that mimics the effects of brain-derived neurotrophic factor (BDNF) on TrkB binding and receptor dimerization, auto-phosphorylation, and activation of neuroprotective molecular pathways. 14 Recent studies found that 7,8-DHF conferred neuroprotective, cognitive-enhancing, and anti-oxidative stress effects in animal models. The present study was undertaken to determine whether 7,8-DHF could attenuate Sco-induced AD-like pathologic changes in rats.
Materials and methods
Animals and treatment
Male Sprague–Dawley rats (250±20 grams) were obtained from the Logistics University of Chinese People's Armed Police Forces. The rats were caged in a reversed 12-hr light/dark cycle at 25°C. The rats were divided into three groups (n=12 for each group)—control (Con), scopolamine (Sco), and scopolamine plus 7,8-DHF (S+D, 7,8-DHF; TCI Europe, Zwijndrecht, Belgium). The Con group was treated with saline, the Sco group was given Sco (1 mg/kg per per day, intraperitoneally [i.p.]), the S+D group was given the Sco (1 mg/kg per per day, i.p) and 7,8-DHF (1 mg/kg per per day, i.p). For the S+D group, the rats were received 1 mg/kg per day 7,8-DHF i.p. for 2 weeks, followed by Sco (1 mg/kg per day)+1 mg/kg per day 7,8-DHF (i.p.) for another 2 weeks. For 7,8-DHF group, the rats were pretreated with 7,8-DHF for 4 weeks. The Sco was administered 0.5 hr before the behavioral test.
Antibodies and chemicals
Antibodies used for western blotting were purchased from Abcam (Cambridge, UK). The kits of superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), glutathione (GSH), and malondialdehyde (MDA) were from Jiancheng Bioengineering Institute (Nanjing, China).
Water maze test
The water maze test was performed as previously described. 15 Briefly, the rats were trained to find the hidden platform through patterns hanging on the wall. Water was colored using ink to provide contrast for the rats. Each rat received four trials per day for 6 continuous days (n=12 for each group). The latency and pathway to search the hidden platform were recorded and used to evaluate the ability of learning and memory. Latency to find the hidden platform during the training and test days and the number of crossings over the platform area on the 7th day when the hidden platform was removed were examined.
Enzyme-linked immunosorbent assay assay
A sandwich enzyme-linked immunosorbent assay (ELISA) was used for quantitative analysis of Aβ levels (n=4 for each group). Briefly, Aβ40 and Aβ42 in samples were captured with specific monoclonal antibodies and then probed with the antibody Biotin-WO2 (Abeta GmBH, Heidelberg, Germany), followed with HRP-NeutrAvidin (Pierce, Rockford, IL). The horseradish peroxidase (HRP) activity was measured using the TMP Microwell Peroxidase System (Kirkegaard and Perry Laboratories, Gaithersburg, MD).
Western blotting
Rats were sacrificed 2 hr after the behavior test. The hippocampus was removed and homogenized in western buffer (Beyotime, Shanghai, China) (n=4 for each group). The protein concentration was measured using the bicinchoninic acid (BCA) method. Equal amounts of protein were separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes (GE Healthcare, Pittsburgh, PA). The membranes were blocked by 3% bovine serum albumin (BSA) and incubated with primary antibodies at 4°C overnight. Then the membrane was incubated with HRP-conjugated secondary antibodies and detected with a chemiluminescent substrate kit (Pierce). The blots were scanned and analyzed using the Kodak Digital Science 1D software (Eastman Kodak Company, Rochester).
Electrophysiological recording
Rats were given urethane (n=4 for each group). The perforant pathway to dentate gyrus (PP-DG) pathway was measured by putting the stimulating electrodes in PP region (AP, 8.0 mm; L/R, 4.0 mm; DV, 3.5 mm) and the recording electrodes in DG region (AP, 4.0 mm; L/R, 2.0 mm; DV, 3.0 mm). A series of different stimulations ranging from 5 mV to 30 mV to elicit the maximum field excitatory postsynaptic potentials (fEPSPs) and 30% maximum stimulus intensity was used as the basic stimulation. High-frequency stimulations (HFS) were induced by four trains of 50 pulses delivered at 200 HZ with a 2-sec inter-train interval.
Statistical analysis
Data are presented as group means±standard error of the mean (SEM). Differences between the groups were analyzed by analysis of variance (ANOVA) followed by the least significant difference (LSD) post hoc test with SPSS (v. 17). p<0.05 was accepted as statistically significant.
Results
7,8-DHF attenuated Sco-induced learning and memory deficits
The Morris water maze was used to test the learning and memory abilities of rats. Rats treated with Sco showed significantly longer latency compared with the Con group; however, rats treated with Sco and 7,8-DHF showed similar latency with the Con group (Fig. 1A). On 7th day, when the platform was removed, Sco-treated rats exhibited longer latency and fewer crossing times compared with the Con and S+D groups (Fig. 1B, C). No defect was observed in the swimming speed of all groups (Fig. 1D). These results suggested that 7,8-DHF treatment effectively restored the learning and memory deficits induced by Sco.
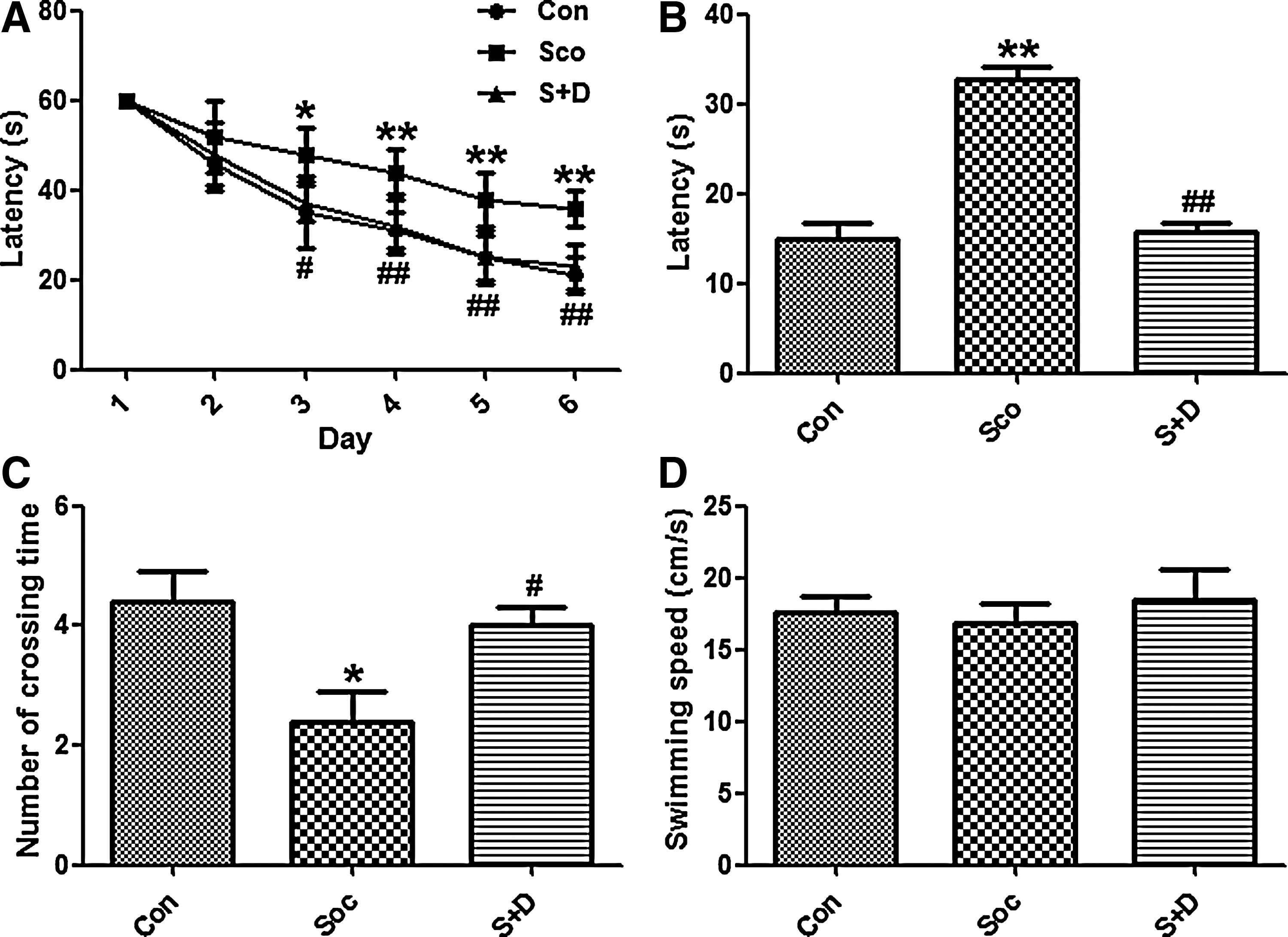
7,8-Dihydroxyflavone (7,8-DHF) attenuated scopolamine-induced learning and memory deficits. (
7,8-DHF reversed Sco-induced synaptic impairments
Synaptic transmission using long-term potentiation (LTP) recordings was described previously. 16 In Sco-treated rats, the amplitude and slope of fEPSP increased approximately 1.3-fold, much lower than the increase in Con group. Supplementation with 7,8-DHF restored the amplitude and slope of fEPSP (Fig. 2A, B). There was no significant difference between the Con and S+D groups. These data suggested that 7,8-DHF effectively reversed the synaptic transmission deficits induced by Sco.
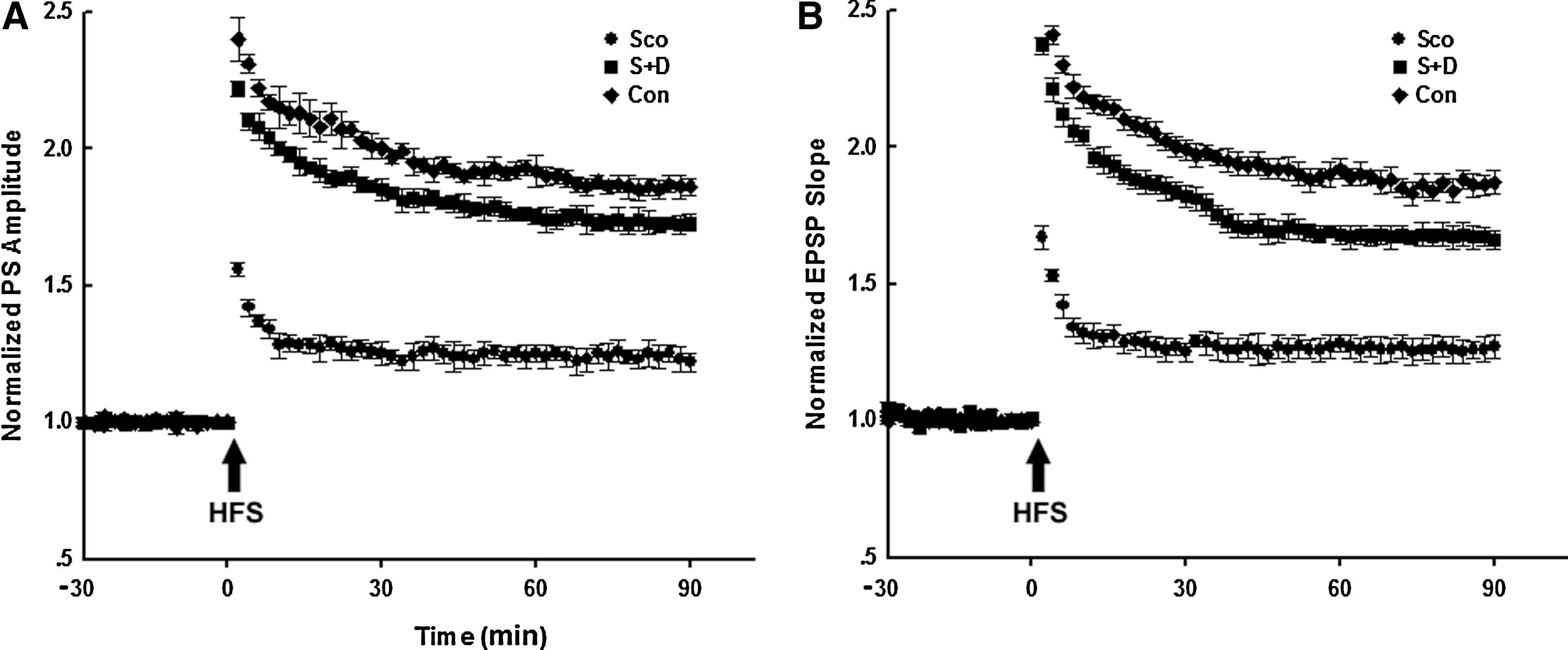
7,8-Dihydroxyflavone (7,8-DHF) reversed scopolamine (Sco)-induced synaptic impairments. (
7,8-DHF reversed Sco-induced Aβ accumulation
Sco administration was found to increase the hippocampal Aβ load remarkably as compared with the Con group. Supplementation with 7,8-DHF was found to decrease the elevated levels of Aβ40 and Aβ42 compared with Sco-treated group (Fig. 3A, B).
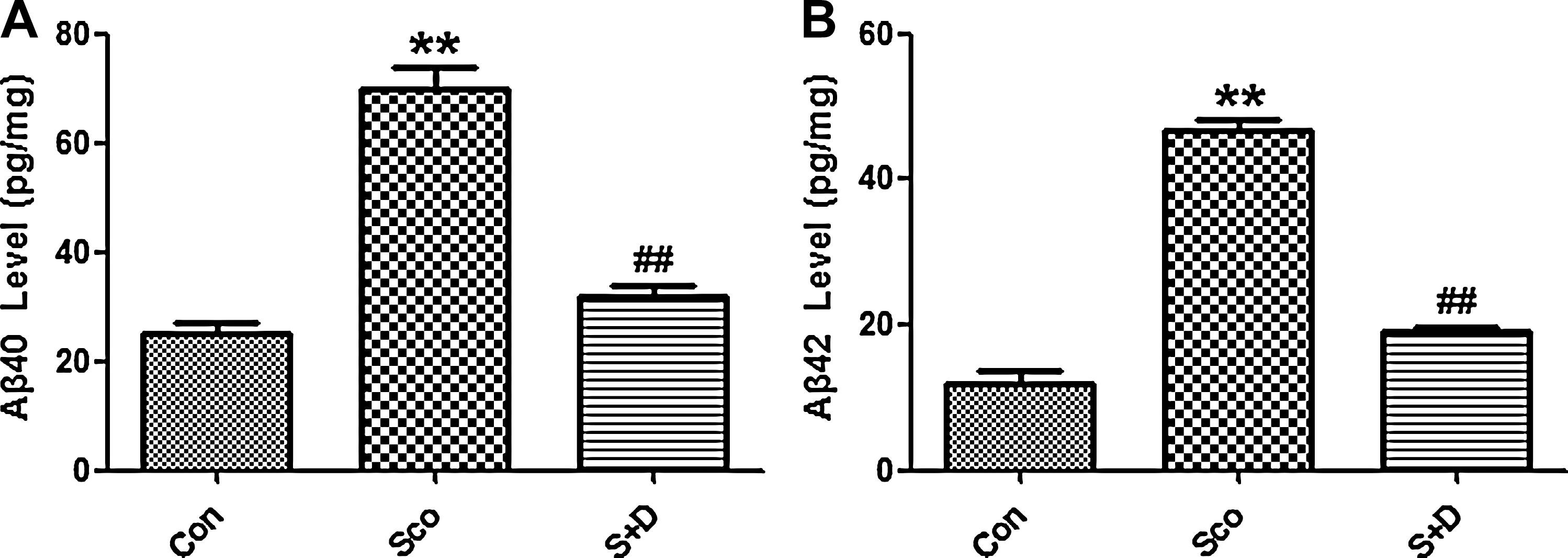
7,8-Dihydroxyflavone (7,8-DHF) reversed scopolamine (Sco)-induced amyloid β (Aβ) accumulation. (
7,8-DHF reversed Sco-induced oxidative stress
Oxidative stress was determined by measuring the levels of MDA and GSH, and the activity of SOD and GSH-Px in the hippocampus. Sco treatment increased the MDA level, while suppressing the GSH level and activity of SOD (Fig. 4A–C). However, there was no difference in the activity of GSH-Px (data not shown). Supplement of 7,8-DHF robustly restored these effects of Sco treatment (Fig. 4A–D). Because MDA represented lipid oxidation and GSH-Px activity indicated the ability of eliminating free radicals, 7,8-DHF could improve lipid oxidation without an effect on clearing free radicals induced by Sco.

7,8-Dihydroxyflavone (7,8-DHF) reversed scopolamine (Sco)-induced oxidative stress. (
TrkB receptor–mediated the protective effect of 7,8-DHF
The BDNF/TrkB pathway has been thought to perform beneficial effects on synaptic and neuronal plasticity. 17 Recently, 7,8-DHF has been identified as a specific TrkB agonist and as playing neuroprotective roles. 18 The phosphorylation level of TrkB was reduced in the hippocampus of Sco-treated rats while it was restored following treatment with Sco plus 7,8-DHF. We further measured the activities of TrkB, AKT, and extracellular signal-regulated kinase (ERK), all of which are essential downstream molecules of BDNF/TrkB pathway. 19,20 These results showed that 7,8-DHF reversed the TrB signaling pathway that was suppressed by Sco (Fig. 5A, B).
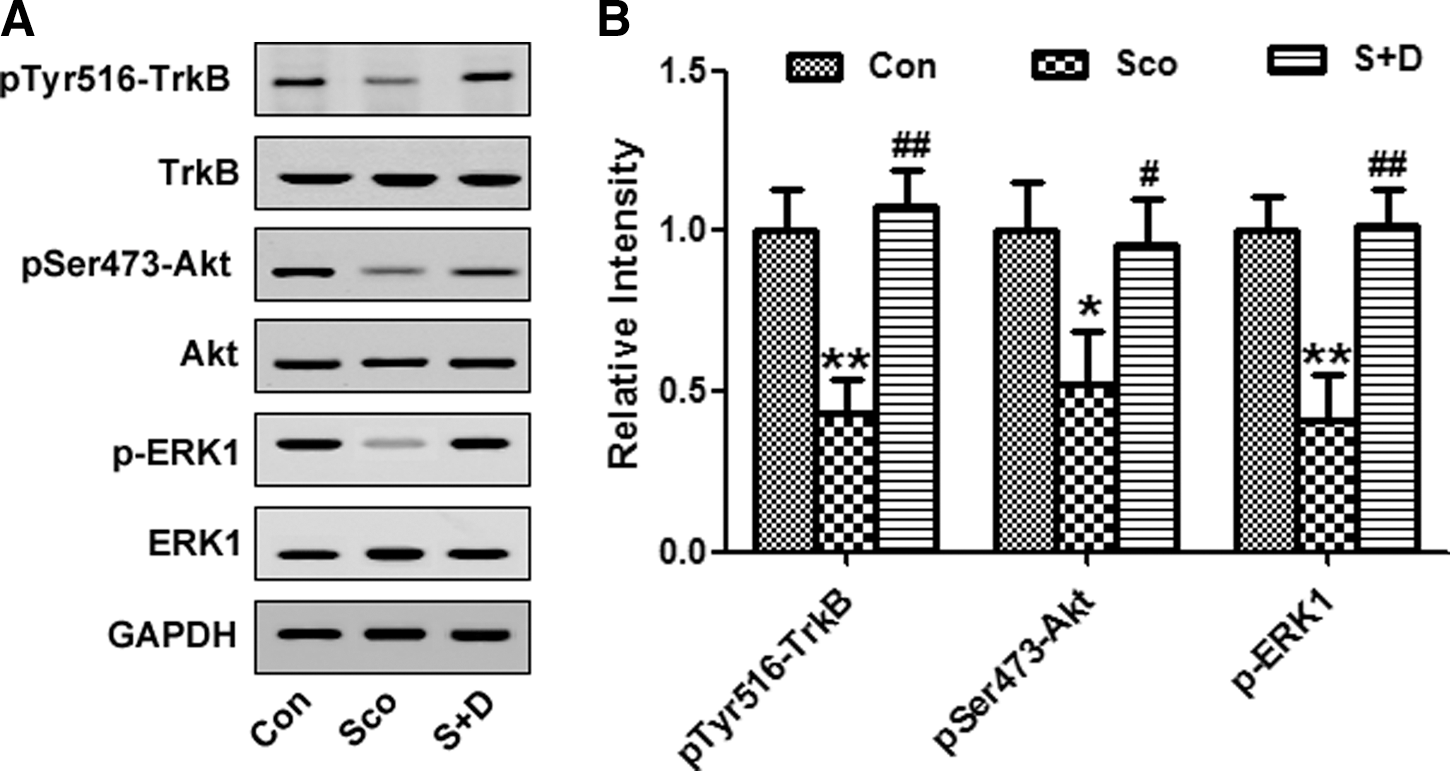
The TrkB receptor mediated the protective effect of 7,8-dihydroxyflavone (7,8-DHF). (
Discussion
The aim of the current study was to evaluate the protective properties of 7,8-DHF on Sco-induced AD-like pathologic dysfunction. The neuroprotective effects of 7,8-DHF have already been reported in previous studies. Most studies have focused on depressive-like or stress-related memory tests. 21 Recently, 7,8-DHF has been found to improve object memory formation in healthy rats. In APPswe/PS1dE9 mice, a transgenic mouse model of AD, administration of 7,8-DHF was found to improve spatial memory. 19 It was found that animals administered 1 mg/kg 7,8-DHF could perform better in an object recognition memory test of healthy rats 19 ; we adopted that concentration here. Similarly, in this study, 7,8-DHF played protective roles against Sco-induced AD-like pathologic dysfunction, including learning/memory impairment, Aβ accumulation, oxidative stress, and synaptic dysfunction, suggesting that 7,8-DHF might be a promising therapeutic target for AD.
Evidence suggests that disturbances in the BDNF-TrkB system induced by Sco affected hippocampal-dependent memory and cognitive dysfunctions. 22,23 BDNF was implicated in learning and memory. Phosphorylation of TrkB activated by BDNF-activated PI3K/Akt signaling in the hippocampus increased during spatial memory formation. 24 Akt was an essential kinase in the regulation of cyclic AMP response element (CRE)-binding protein (CREB) activation, and in turn modified the expression of molecules essential for learning and memory. 25 The activity of Akt was decreased in AD transgenic mice as indicated by decreased phosphorylation. 26 ERK was also an important kinase for Tau phosphorylation and cellular survival; it was also activated by TrkB activation via MEK/ERK signaling. Consistent with Moosavi et al., Sco remarkably suppressed the activity of Akt and ERK. 27 Herein, we have shown that in vivo activation of TrkB by supplementation with 7,8-DHF, a specific TrkB agonist that mimics the effect of BDNF, improved the spatial learning and memory in cognitively impaired rats induced by Sco.
Synaptic transmission represented the synaptic function, which was critical for learning and memory. Sco treatment inhibited synaptic function by impairing synaptic transmission as shown by impaired LTP. 11 7,8-DHF treatment counteracted the dysfunction of synaptic transmission in the hippocampus. In addition, 7,8-DHF reversed Sco-induced Aβ accumulation and oxidative stress. The neurotoxic effects of Aβ have been widely studied, including impairing synaptic plasticity, inducing apoptosis, and promoting Tau phosphorylation and oxidative stress. 28 Sco has been found to induce Aβ deposition and oxidative stress, and our work has identified the neuroprotective role of 7,8-DHF against Sco. On the basis of these findings, 7,8-DHF may protect memory deficits by activating the TrkB signaling pathway.
Footnotes
Acknowledgments
This work was supported by the National Nature Scientific Fund of China (81271392, 81301050) and the Fund of Logistics University of Chinese People's Armed Police Forces (WHTD201306-1).
Author Disclosure Statement
No competing financial interests exist.