
Abstract
Decline of cognition and increasing risk of neurodegenerative diseases are major problems associated with aging in humans. Of particular importance is how the brain removes potentially toxic biomolecules that accumulate with normal neuronal function. Recently, a biomolecule clearance system using convective flow between the cerebrospinal fluid (CSF) and interstitial fluid (ISF) to remove toxic metabolites in the brain was described. Xie and colleagues now report that in mice the clearance activity of this so-called “glymphatic system” is strongly stimulated by sleep and is associated with an increase in interstitial volume, possibly by shrinkage of astroglial cells. Moreover, anesthesia and attenuation of adrenergic signaling can activate the glymphatic system to clear potentially toxic proteins known to contribute to the pathology of Alzheimer disease (AD) such as beta-amyloid (Abeta). Clearance during sleep is as much as two-fold faster than during waking hours. These results support a new hypothesis to answer the age-old question of why sleep is necessary. Glymphatic dysfunction may pay a hitherto unsuspected role in the pathogenesis of neurodegenerative diseases as well as maintenance of cognition. Furthermore, clinical studies suggest that quality and duration of sleep may be predictive of the onset of AD, and that quality sleep may significantly reduce the risk of AD for apolipoprotein E (ApoE) ɛ4 carriers, who have significantly greater chances of developing AD. Further characterization of the glymphatic system in humans may lead to new therapies and methods of prevention of neurodegenerative diseases. A public health initiative to ensure adequate sleep among middle-aged and older people may prove useful in preventing AD, especially in apolipoprotein E (ApoE) ɛ4 carriers.
Introduction
D
Of special interest is the metabolism of the beta amyloid (Abeta) protein. Abeta has numerous normal functions, e.g., as a transcription factor, activation of kinases, oxidative stress protection, cholesterol transport, and protection from microbes. Unfortunately, Abeta can damage neurons, and it is the key component of the amyloid plaques found in AD. Known mechanisms of Abeta clearance include uptake by microglial phagocytosis, receptor-mediated transport across the blood vessel walls, and degradation by enzymes, e.g., glutamate carboxypeptidase II, insulin-degrading enzyme, matrix metalloproteinase-9, neprilysin, and others. 2 Similar mechanisms exist for other toxic, potentially pathological brain proteins such as α-synuclein and tau. For example, tau is degraded by autophagic and proteasomal degradative systems and a variety of proteases, including aminopeptidases, HTRA1, calpain, caspases, and thrombin. 3
The recent discovery of the glymphatic clearance system provides fresh insight into how the brain clears potentially toxic metabolites. 4 –6 Unlike most of the other organs in the body, the brain lacks lymphatic vasculature for clearing excessive interstitial fluid, metabolic waste products, and unneeded extracellular biomolecules, such as Abeta (Fig. 1). However, the brain does have the glymphatic system, which accomplishes the same purpose and protects fragile neurons from the toxic side effects of metabolism.
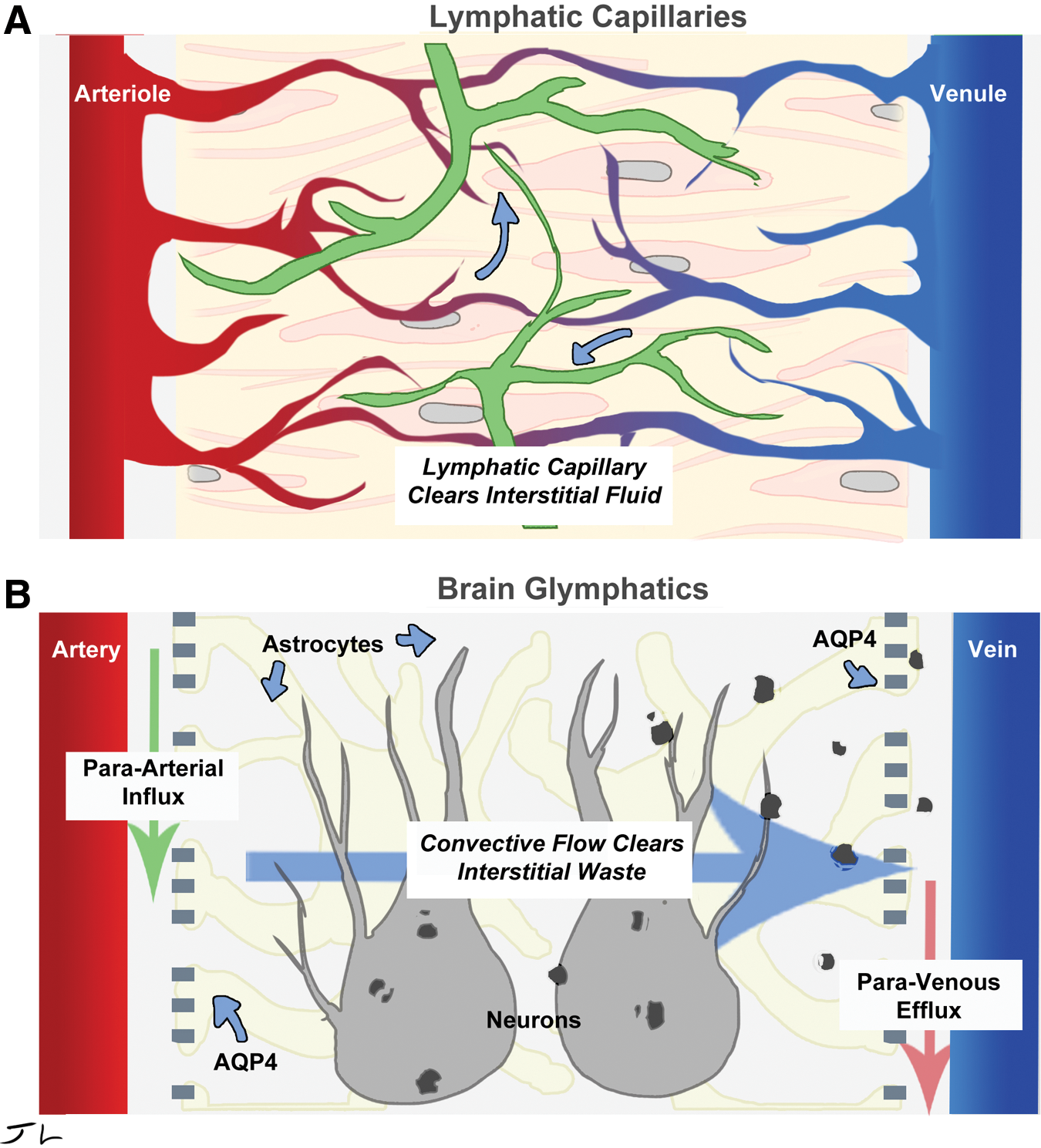
Glymphatic system works analogously to the lymphatic system. Lymphatic vessels clears interstitial fluid of waste (
The glymphatic system uses convective flow to clear biomolecules. Cerebrospinal fluid (CSF) enters the brain parenchyma (functional parts) along a para-arterial route and exchanges with the interstitial fluid (ISF). The ISF carries extracellular solutes from the interstitial (extracellular) space in the brain along para-venous drainage pathways. Aquaporin 4 (AQP4) water channels in the astrocytes, which encircle the brain's vasculature, are required for glymphatic function and facilitate clearance of soluble proteins, waste products, and excess extracellular fluid. This system was termed the “glymphatic” pathway due to its dependence upon glial cells and its performance of peripheral ‘lymphatic’ functions in the central nervous system (CNS) 4 (Fig. 1). From initial reports, it became clear that the glymphatic system was responsible for clearing 65% of Abeta, 5 and may be the major pathway by which metabolites are removed from the interstitial spaces of the brain.
Sleep Facilitates Clearance of Metabolites from the Brain
Xie et al. 7 now report that sleep plays a critical role in maintaining homeostasis in the brain. Natural sleep or anesthesia is associated with a 60% increase in the interstitial space, which in turn significantly increases convective exchange of CSF with ISF via the glymphatic system, removing potential neurotoxic waste products such as Abeta that accumulate during waking periods. This mechanism contributing to the restorative nature of sleep was unknown in spite of the numerous negative consequences of sleep deprivation: Prolonged reaction times, reduced cognitive performance, 8 increased risk of seizures or hallucinations, 9,10 and even death in a matter of days for flies 11 or weeks for rodents. 12 In humans, the syndrome known as progressive familial insomnia leads to dementia and death. 13
The recent discovery of the glymphatic system prompted the authors to test the hypothesis that sleep might prevent the negative consequences of insomnia by altering glymphatic function. Using sophisticated neuroscience and cell biology techniques, Xie et al. discovered that infusion of a small (3 kD) green fluorescent tracer molecule (fluorescein isothiocyanate–dextran [FITC–dextran]) via the CSF into brains during the day, when the mice were asleep, rapidly penetrated the brain (the specific regions noted were the peri-arterial spaces, the sub-pial regions, and the brain parenchyma). The tracer was visualized by two-photon imaging, in which two low-energy infrared photons need to be absorbed simultaneously by the tracer to stimulate emission, providing very high spatial resolution. Electrocortigraphy (EcoG), in which electrical signals from electrodes physically localized within the brain are analyzed, was used to determine that the mice were indeed asleep by the telltale appearance of slow wave patterns. After waking the mice by gently touching their tails, CSF loaded with a second fluorescent tracer of a different color, Texas Red–dextran (3 kD) was injected. This time, tracer influx was significantly reduced. EcoG showed decreased slow wave activity, consistent with awake animals. To confirm these results, the experiments were repeated in the evening when the animals are awake. This time the first tracer, FITC–dextran in CSF, was barely detectable within the brain. After 30 min, the mice were anesthetized with ketamine/xylene, and the second tracer Texas Red–dextran was injected. This tracer immediately entered the brain, finding its way to the same regions previously observed in the earlier experiment when the mice were sleeping. The authors concluded that the brain is more permeable to CSF when the animals are asleep. 7 How does this occur?
There are at least two possible mechanisms to explain the restriction of CSF flow within the awake brain. Perhaps arterial pulse waves that force the CSF inward are greater during wakefulness, or the interstitial space is reduced, increasing resistance to convective flow. Given that arterial blood pressure is higher during physical activity, and high pressure would presumably drive the CSF out into the interstitial spaces, the first hypothesis was deemed unlikely. So the authors investigated the second hypothesis. To measure the brain interstitial space they employed the widely used real-time iontophoretic tetramethylammonium (TMA) method in head-fixed mice. In the TMA method, an electrode with two components is embedded into the mouse brain. The first component uses an electric pulse to release a small amount of positively charged TMA to diffuse into the IFS. The second component is positioned a defined distance away from the first and specifically detects the presence of TMA. The amount of TMA detected in a defined time is proportional to the volume of the IFS.
TMA recordings when the mice were asleep indicated that the IFS volume was 23.4±1.9%, compared to 14.1%±1.8% when the mice were awake in the evening. EcoG readings confirmed the state of consciousness of the animals was as expected. IFS volume was increased when the animals were asleep, confirming the hypothesis. Furthermore, in experiments in which the waking mice were anesthetized, IFS volume increased consistently, >60% from 13.6±1.6% to 22.7±1.3%. Cortical IFS varied from 13% to 15% when mice were awake to 22% to 24% when they were asleep. Similar results were found when measurements were made 150 μm or 300 μm below the brain's surface. In addition the tortuosity (twistedness) of the IFS was unchanged. 7 The increased IFS volume during sleep decreases tissue resistance to IFS flux and inward movement of CSF (Fig. 2).
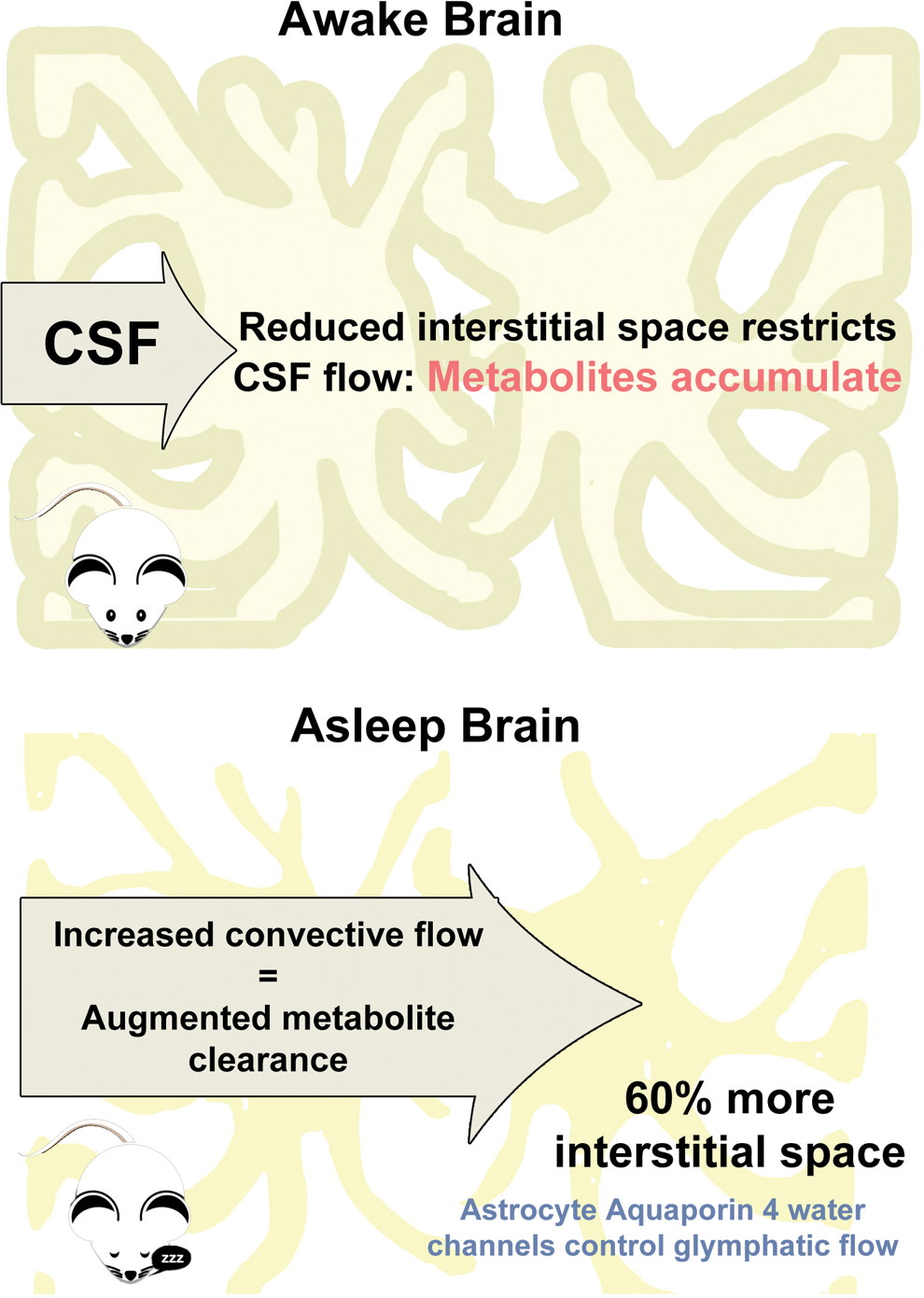
Sleep increases glymphatic flow by increasing interstitial space. (Top) In the awake brain, reduced interstitial space restricts cerebrospinal fluid (CSF) flow, allowing metabolites to accumulate. In the sleeping brain, aquaporin 4–mediated changes in astrocyte cell volume increase interstitial space to increase CSF access. (Bottom) Resulting increased convective flow augments metabolite clearance. Color images available online at
The next question is whether the increased convection during sleep results in increased clearance of metabolites or potentially neurotoxic biomolecules. Because the glymphatic system had been previously reported to clear 65% of Abeta, Abeta clearance was measured during sleep, wakefulness, and under anesthesia in mice using 125I-labeled Abeta. Brains were isolated 10–240 min later, and Abeta retention was measured. Consistent with the important role of the glymphatic system in clearance, Abeta was cleared twice as fast in sleeping and anesthetized mice as in awake mice. To completely rule out other transport effects (e.g., Abeta is also removed by receptor-mediated transport across the blood–brain barrier [BBB]), radiolabeled inulin was also observed to be cleared twice as fast in sleeping and anesthetized mice. 7
What triggers the sleep-associated increase in IFS and biomolecular clearance? Circadian rhythms are not a likely explanation, because anesthesia can be administered at any time to reduce interstitial space and activate clearance. Instead, the authors hypothesized that state of arousal itself was responsible. In particular, locus coeruleus noradrenergic signaling, which is associated with alertness/wakefulness, may be the direct cause. In other words, adrenergic signaling in the awake state reduces interstitial volume and decreases clearance. To test the hypothesis, awake mice were treated with a cocktail of adrenergic antagonists 15 min before CSF loaded with tracer was administered. As expected, tracer influx was increased in the presence of the antagonists. Next Xie et al. measured the effects of adrenergic inhibition on interstitial space using the TMA method after directly applying the inhibitors to the exposed cortical surface of the brains of awake mice. Interstitial volume increased from 14.3±5.2% to 22.6±1.2%, similar to that seen with sleep or anesthesia. Consistent with the known effects blocking adrenergic signaling, more slow wave patterns were observed with EcoG measurements indicating a more relaxed, sleep-like state was induced. 7 Therefore, the adrenergic system not only regulates wakefulness but also interstitial volume and glymphatic clearance. Which cells are involved in mediating interstitial clearance? That remains unknown, although sleep-mediated shrinkage of the astroglial and other neuronal support cells would be the leading hypothesis.
Overall, Xie et al. provide evidence that a key purpose of sleep as a physiological process is to clear potentially toxic waste products of metabolism, such as Abeta, which is known to trigger irreversible neuronal injury, interfere with synaptic transmission, and is the critical component of toxic amyloid deposits in AD. We can speculate that the accumulation of waste products somehow helps trigger sleep in conjunction with circadian rhythms and that increased interstitial volume associated with sleep may be involved in long-term memory consolidation, which occurs during sleep. These ideas might be the subject of subsequent work.
This study is a potential breakthrough in our understanding of sleep and brain physiology and has significant implications. However, to date direct evidence linking the glymphatic system to a critical role in sleep has been reported by primarily by one group. This work needs to be independently replicated, especially in other mammals, including humans. So far, evidence for the existence of the glymphatic system has been reported by the same group for rats. 6 Unfortunately, direct access to the human brain in more problematic than in mice, and such work may require modification of existing noninvasive interventions. In another report, the same group suggests lumbar intrathecal (spinal canal) injection of tracers visible by contrast-enhanced magnetic resonance imaging (MRI) for studies in humans. 6,14
There is potential benefit in retrospectively reviewing earlier results in light of this new report. For example, accumulation of adenosine in the brain was reported to drive induction of sleep. 15 Modulation of the glymphatic system provides a natural mechanism for adenosine accumulation and removal, 16 as well a potentially more general framework for how sleep may be regulated.
Correlations of Sleep with Neurodegeneration and Aging
If sleep really drives clearance of toxic biomolecules by altering the function of the glymphatic system in humans, then we would expect significant correlations of amount and quality of sleep with human neurodegenerative disease and longevity. And indeed there is some evidence that this is the case beyond the strong associations seen with lack of sleep and mortality/neurodegeneration in flies and rodents. 11,12 Most relevant is that IFS has previously been reported to decline from 19% to 13% in young versus old mice. 17
In humans there are consistent reports that both too little (generally less than 7 hr of sleep) and too much (generally greater than 9 hr of sleep) are associated with increased mortality and decreased quality of life. This correlation occurs in older adults, but earlier sleep disruptions may play a role in longevity. Cause and effect are notoriously difficult to establish in such studies, and differences in end points and methodology make such studies problematic for effective meta-analyses. Many of the studies were focused on connections to cardiovascular disease, which has been correlated with altered sleep. 18 Likely, death occurs before typical onset of neurodegenerative diseases. For direct support of sleep-associated glymphatic clearance, more studies focused on the role of sleep on neurodegeneration and cognition are needed, 19 although those that have been performed are consistent with sleep playing an important role. 20 The evidence suggests that organ systems other than the brain are more closely tied to sleep dysfunction and mortality; however, at a minimum, cognitive ability deteriorates with altered sleep. Regardless, the association of mortality with too much sleep is especially interesting, suggesting a “Goldilocks” effect where just the correct amount of sleep is needed to avoid problems, and the existence of trade-offs in sleep versus waking physiology. We speculate that brain function may suffer from too much time spent “house cleaning,” as well as too little.
What About More Direct Evidence for a Role of Sleep in Preventing Neurodegenerative Diseases Such as AD in Humans?
A small clinical study ∼70 people) showed that subjects with shorter sleep duration or shorter sleep quality have an associated greater Abeta burden, measured by a noninvasive probe for amyloid (Pittsburgh compound B) positron emission tomography. 21 This result is completely consistent with the potential role of sleep-associated glymphatic waste removal. Although this is the best example of a potentially direct connection of amyloid plaque in humans with sleep duration and quality, a significant set of data are consistent with this result (see the review in ref. 20] for more details).
Changes in sleep pattern may predict AD. In a 9-year study, 214 dementia-free Swedish adults (≥75 years old, mean age 83.4 years old) were followed. In all, 40% of the subjects reported a change in sleep pattern. Between years 6 and 9, 28.5% of were diagnosed with dementia, 22.0% of whom had AD. Reduced sleep was associated with a 75% increased all-cause dementia risk (hazard ratio [HR], 1.75; 95% confidence interval [CI] 1.04–2.93; p=0.035) and double the risk of AD (HR, 2.01; 95% CI 1.12–3.61; p=0.019) after adjusting for age, gender, and education. Only adjusting for depression, a frequent correlate of sleeplessness, caused a loss of significance. Lifestyle, vascular status, and ApoE status did not alter the results. Sleeping pills proved ineffective. 22 In these types of correlative studies, cause and effect are always somewhat unclear, but it is interesting that sleeping pills, which presumably would induce an anesthesia-like state similar to that seen in the mouse studies, were ineffective. Because sleeping pills may not result in quality sleep, more careful investigation of sleeping stage (non-rapid eye movement [NREM] stages, rapid eye movement [REM], etc.) and glymphatic function may prove enlightening.
Even more intriguing is a recent report that sleep is a significant attenuator of risk for AD for people carrying the ApoE ɛ4 allele. The ApoE ɛ4 allele increases risk for patients to develop AD by approximately two-fold for people with one copy, and as much as 10-fold for those carrying two copies by age 79. Risk for those carrying two copies of the ApoE ɛ4 allele may be a high as 70% by age 79. In this study 698 community-dwelling older adults without dementia (mean age, 81.7 years; 77% women) were followed for 6 years. Subjects were tested for ApoE status, sleep quality/duration by 10 days of actigraphy (which uses a wrist watch–like device to measure movement and activity). During the follow-up period, 98 individuals developed AD. Better sleep quality attenuated the effect of the ApoE ɛ4 allele on the risk of incident AD (HR, 0.67; 95% CI 0.46–0.97; p=0.04 per allele per 1 standard deviation [SD] increase in sleep quality/duration) and on the rate of cognitive decline. From postmortem examination of the 201 people who died during the study, better sleep quality attenuated the effect of the ApoE ɛ4 allele on neurofibrillary tangle density, which accounted for the effect of sleep quality on the association between APOE genotype and cognition before death. These data support the possibility that supporting improved sleep may reduce the risk of AD and development of neurofibrillary tangles in APOE ɛ4+ individuals. 23
Medical Implications
The medical implications of these results are profound. If the glymphatic system plays as important role as the Xie et al. results suggest, then glymphatic dysfunction may play a hitherto unsuspected role in the pathogenesis of neurodegenerative diseases as well as the maintenance of cognition. Such mechanisms would be critical to unravel, and likely work is ongoing to achieve this goal.
If improved sleep can help prevent or delay neurodegenerative diseases, especially AD, then a public health effort should be made to achieve better sleep, especially among middle-aged and older people. A special effort should be made for people carrying the ApoE ɛ4 alleles, as it appears that good sleep reduces the risk of carriers by 33%. A Japanese study suggested that a short nap (30 min between 1300 and 1500 hr) in which the subject does not go into a deep sleep (which is actually counterproductive) and moderate exercise such as walking could positively impact sleep quality. 24 In fact, short naps may actually help prevent AD. 25 A 4-week trial of this intervention demonstrated greatly improved sleep efficiency, as well as physical and mental health. 24
Developing new drugs that stimulate removal of waste products in the brain is an exciting possibility. However, it is possible that some such drugs may already exist. Because adrenergic antagonists recapitulated the effects of sleep in awake animals, perhaps adrenergic inhibitors could prove useful. Perhaps β-adrenergic blockers presently used to control systolic high blood pressure should be evaluated. Intriguingly, a recent study subjects taking β-adrenergic blockers (BB) showed reduced risk of AD. 26 However, in this case all treatments that reduced blood pressure decreased the risk of AD including diuretics, angiotensin-1 receptor blockers (ARB), angiotensin-converting enzyme inhibitors (ACE-I), and calcium channel blockers (CCB). Interestingly, the beneficial effect was in addition to or independent of mean systolic blood pressure. However, in an earlier study of patients with incipient AD, BB, but not ACE inhibitors, were associated with a reduced rate of the progression of AD. 27
Conclusions
The existence of a sleep-driven convective system in the brain to remove metabolic waste products has profound consequences for maintenance of brain function during aging. Further characterization of the glymphatic system in humans may lead to new therapies and methods to prevent neurodegenerative diseases. A public health initiative to ensure enough proper sleep in middle-aged and older people may prove useful in preventing AD, especially among ApoE ɛ4 carriers. The benefits of sufficient sleep to cognition and quality of life have been evident since (and before) the times of Hippocrates, but these benefits may be greater than originally imagined!
Footnotes
Acknowledgments
The authors wish to acknowledge the graphic design work of Ms. Jasmine Larrick.
Author Disclosure Statement
No competing financial interests exist.