
Abstract
Dehydroepiandrosterone (DHEA) and dehydroepiandrosterone sulfate (DHEAS) are the most abundant circulating adrenal steroid hormones. The plasma level of DHEAS correlates with longevity in primates and varies during human development, with a maximum in early adulthood and a marked decline during aging. DHEA promotes the expression of molecular chaperones that are housekeeping constitutive or stress response proteins essential for the processes of folding, translocation, maintenance, and disaggregation or degradation of misfolded polymers (proteins, RNA, and DNA), as well as for homeostasis, immune response, and cancer resistance. The level of chaperone expression correlates with longevity and shows a decline during aging. DHEA-induced promotion of chaperone expression could contribute to the epigenetic evolution of primate longevity.
Introduction
T
Dehydroepiandrosterone (DHEA) and its sulfate (DHEAS) are the most abundant adrenal steroid hormones in human blood. The adult adrenal secretes approximately 6–8 mg/day of DHEA and 8–16 mg/day of DHEAS. 7 In plasma, the half-life of DHEA is approximately 15–30 min, whereas that of DHEAS is approximately 10–20 hr, resulting in a plasma level of DHEAS approximately 500-fold higher than that of DHEA. Contributing to the relatively high plasma level of DHEAS are the effective conversion of DHEA to DHEAS in the liver and the enterohepatic recirculation of DHEAS. This makes the plasma level of DHEAS in humans a relatively stable representation of total adrenal DHEA+DHEAS production.
A more pronounced diurnal variation of plasma DHEAS, with a maximum in the morning, is observed in shorter-lived primates.
8
The biological actions of DHEA involve steroid receptors as well as multiple G-protein-coupled, membrane-associated receptors and DHEA-specific receptors.
9
The biological effects of DHEA also derive from its role as a precursor for testosterone and estradiol. In adults, the conversion of DHEA occurs mainly in the gonads, whereas during aging the sex hormones are to a large extent converted in peripheral tissues. This mechanism, called intracrinology, accounts for 30%–50% of androgen synthesis in aging males and 100% of estradiol synthesis in postmenopausal females. The generation of sex hormones by intracrinology is advantageous because the hormones are delivered directly to the target tissues without general systemic exposure.
10
Plasma DHEA and DHEAS levels show major variation during human development, with maxima occurring in late fetal life and early adulthood, followed by a marked decline to 10%–15% of peak values during senescence.
11
In addition to synthesis in the adrenals, DHEA and DHEAS are “neurohormones” produced in the brain by astrocytes as well as by neurons,
12
leading to concentrations of DHEA approximately 10-fold higher in brain tissue than in plasma. This difference was observed to be less pronounced in Alzheimer's disease patients than in age-matched controls.
13
Thus, a defect in DHEA-mediated neuroprotection may predispose an individual for the development of Alzheimer's disease. Accordingly, DHEA counteracts amyloid-β protein toxicity as well as oxidative stress, anoxia, and N-methyl-
In the central nervous system (CNS), the level of DHEA is relatively high in the hippocampus, prefrontal cortex, and amygdala, which are areas associated with cognition and behavior as well as with the intracrine conversion of DHEA to estradiol and testosterone. 15
The molecular chaperones are housekeeping constitutive or stress response proteins essential for the proofreading, folding, and translocation of other polymers (proteins, RNA, or DNA), as well as for the disaggregation or degradation of misfolded polymers and for the structure and function of steroid hormone receptors. The molecular chaperone families referred to are the protein chaperones (Hsp and Hsc), the DNA chaperones (RecQ proteins), and the RNA chaperones (DEAD-box proteins). Here, emphasis is placed on the possible significance of these chaperones for the mechanisms of aging and longevity. 16 A role for DNA chaperones in longevity is supported by studies of the WRN chaperone (RecQ-like helicase), which when mutated results in Werner's syndrome of premature aging. 2 Additionally, the importance of the molecular chaperones as common mediators of longevity can be supposed from the observation that the chaperone-generating Forkhead box O (FOXO) transcription factors serve as a convergence point for multiple steroid hormone-dependent longevity pathways. 17,18 Chaperone expression is relatively high in brain and nerve tissues 19 and varies in level during development. In the retina, for example, the level of the constitutive chaperone heat shock protein 70 (Hsc70) shows peaks of expression in early infancy and adulthood followed by a 50% decline during aging. 20
Additionally, the effectiveness of the maintenance and repair mechanisms tends to decline with age, in parallel with the progressive functional impairment of the chaperone defense. For example, the 30%–50% decline in the constitutive levels of the endoplasmatic reticulum chaperones is suggested to constitute one of the central defects in Alzheimer's disease and could lead to many of the physiological declines associated with aging. 21 A significant association was also observed between decreasing Hsp70 levels and the degree of dependency and cognitive decline in geriatric patients, 22 as well as between the age-related decline in constitutive Hsc70-dependent, chaperone-mediated lysosomal autophagy and certain neurodegenerative disorders. 23 Thus, chaperone deficits most likely contribute to the frailty of aging.
The promotion of chaperone expression by DHEA may underlie the cytoprotective effects of DHEA, as well as its preventive effects in relation to a number of diseases, including infection, allergy, cancer, osteoporosis, cardiovascular diseases, immunosenescence, mental disorders, and Alzheimer's disease, as recently reviewed elsewhere. 24 –26
This review emphasizes the chaperones of DHEA synthesis and of DHEA receptors, as well as the evidence that DHEA promotes chaperone expression and that the plasma level of DHEAS correlates with the maximum life span of primates. It is suggested that the hormonal promotion of chaperone expression contributes to the epigenetic evolution of primate longevity.
DHEA/Chaperone interactions.
Chaperones of DHEA synthesis
The synthesis of the DHEA precursor pregnenolone depends on the chaperoning of cholesterol across the outer mitochondrial membrane by means of the translocator protein (TSPO, the former peripheral benzodiazepine receptor). This TSPO-mediated translocation of cholesterol is the rate-determining step of steroidogenesis. As with other chaperones, the expression of TSPO increases in response to the stress of inflammation or injury 27 and declines during aging. This age-related reduction in TSPO expression and neurohormone generation is counteracted by the chemical co-chaperone-like effects of recently discovered TSPO ligands. 28
Chaperones of DHEA receptors
The biological actions of DHEA involve steroid receptors as well as G-protein–coupled membrane-associated receptors. 9 In both cases, the molecular chaperones are important to receptor function. For example, the structure and activity of the steroid hormone receptors depend on the molecular chaperones heat shock protein 90 (Hsp90), Hsp70, Hsp40, and various co-chaperones. In absence of the ligand, this complex is present in the cytoplasm in a soluble form. Upon exposure to the ligand, the chaperone dissociates from the receptor, which in complex with the ligand associates with DNA and the transcriptional complexes. 20
Concerning the G-protein–coupled membrane receptors, DHEA is an endogenous ligand for and promoter of the expression of the cardio-protective Sigma-1 receptor, which was originally recognized as an opioid receptor. 30 The Sigma-1 receptor is an endoplasmatic reticulum chaperone that is highly enriched in the brain. This chaperone resides in lipid rafts at the endoplasmatic reticulum–mitochondria interface, a membrane micro-domain essential for signaling processes. 31 This is illustrated by the ability of DHEA to ameliorate cognitive impairment in mice by stimulating the activity of the Sigma-1 chaperone. 32 This chaperone has also been shown to promote cell survival, at least in part by up-regulating the anti-apoptotic Bcl-2 protein. 33 Evidence for the importance of the Sigma-1 chaperone for cell survival also comes from the observation that silencing of this chaperone induces cell death. 34 The chaperone ANKRD13C functions specifically to regulate the biogenesis and the exit of G-protein–coupled receptors from the endoplasmatic reticulum. 35
DHEA promotion of chaperone expression.
A neuroprotective effect of DHEA results from its activation of the G-protein–coupled nerve growth factor (NGF) receptor, 36 which leads to an increase in the expression of the chaperones Hsp27 and Hsp70 with resultant improvement of stress resistance. 37 The DHEA-promoted increased expression of Hsp70 occurs in parallel with improvements in survival rates in rodent models of trauma (abdominal surgery), hemorrhage (hypotensive shock), and inflammation (femoral fractures). This suggests DHEA as a possible option in the treatment of critical illness. 38 Adjuvant effects of DHEAS and Hsp70 have also been observed in attempts to improve the anti-bacterial immune response. 39
Topical DHEA on aged skin gave rise to an improvement of pro-collagen biosynthesis and an increase in the expression of the chaperone Hsp47, suggesting the use of topical DHEA as an anti-aging skin agent. 40 A meta-analysis of genome-wide association data identified eight independent common human genetic variants associated with aging and serum DHEAS concentration. Here the strongest co-variation signal was given by a variant on chromosome 7 associated with the expression of zinc finger proteins. 41 In the cold shock RNA chaperones, the zinc finger domain determines the activity and nucleic acid–binding properties of the chaperone. 42 In addition, the zinc finger–like region of the Hsp40 is required for its co-operation with Hsp70 to chaperone non-native polypeptides. 43 DHEA stimulates the adenosine triphosphatase ATPase) and adenosine triphosphate (ATP) synthase activity of brain and liver mitochondria in rats. 44 Activity of the ATP synthase depends on the α-subunit, which is an early-response heat shock protein related to the chaperone Hsp60. 45
The intracrine conversion of DHEA to sex hormones in peripheral tissues is of significance for chaperone expression. Accordingly, both estradiol and testosterone promote the expression of the protein chaperone Hsp70. 46 In addition, estradiol but not testosterone up-regulate the expression of the DNA chaperone Werner syndrome protein (WRN). 47
Concerning the observed DHEA/chaperone interactions, it is interesting to note that chaperone mechanisms are involved in the generation of DHEA as well as in the formation and function of the steroid hormone receptors, which signals the DHEA-induced increase in chaperone expression, thus constituting a kind of positive chaperone feedback. This pattern of hormone–chaperone interactions resembles those observed for cortisol 5 and probably holds true also for the effects of the sex hormones on chaperone expression.
An increasing body of evidence supports the suggestion that epigenetic phenomena are involved in the mechanisms of aging and longevity. Thus, methylation of gene promoters might contribute to the progressive age-related silencing of housekeeping genes, 3 such as the molecular chaperones. Estradiol, but not testosterone, counteracts this type of gene silencing. 48 Additionally, DHEA counteracts gene methylation as induced by exposure to dimethyl nitrosamine. 49
DHEAS and Longevity
The data demonstrating the correlation of maximum life span and serum DHEAS levels for a broad range of primates is given in Table 1 and illustrated in Fig. 1. This correlation of serum DHEAS level and primate longevity corroborates observations suggesting that increased DHEAS production, with high circulating levels of DHEAS, is one of the mechanisms underlying the evolution of human longevity. 55 An involvement of molecular chaperones in the evolution of primate longevity is suggested by the observation that the retinal level of the constitutive chaperone Hsc70 is twice as high in humans as in rhesus monkeys (Macaca mulatta). 20 Still, the causal structure of the DHEAS/longevity correlation remains unresolved. That plasma DHEAS is not the sole determinant of mammalian longevity appears from the fact that plasma DHEAS is relatively high in the short-lived hamster 56 and below the level of detection in cattle and horses. 53 In addition, the difference in longevity between genders correlates inversely with the difference in DHEAS level. 11 Probably the high level of estrogens in females poses a greater advantage to longevity than the small difference in DHEA levels between genders. For example, the estradiol-specific promotion of WRN expression 47 and counteraction of the age-related silencing of longevity genes 48 pose a specific advantage to female longevity. The correlation of DHEAS level with primate longevity resembles that observed for cortisol in relation to the longevity of mammals. In contrast to DHEAS, cortisol shows a tendency to increase with age, particularly in centenarians. 57 In spite of these deviations, the observed correlations of hormone level and species longevity suggest that the promotion of longevity by steroid hormones observed in model organisms 58 holds true in primates as well.
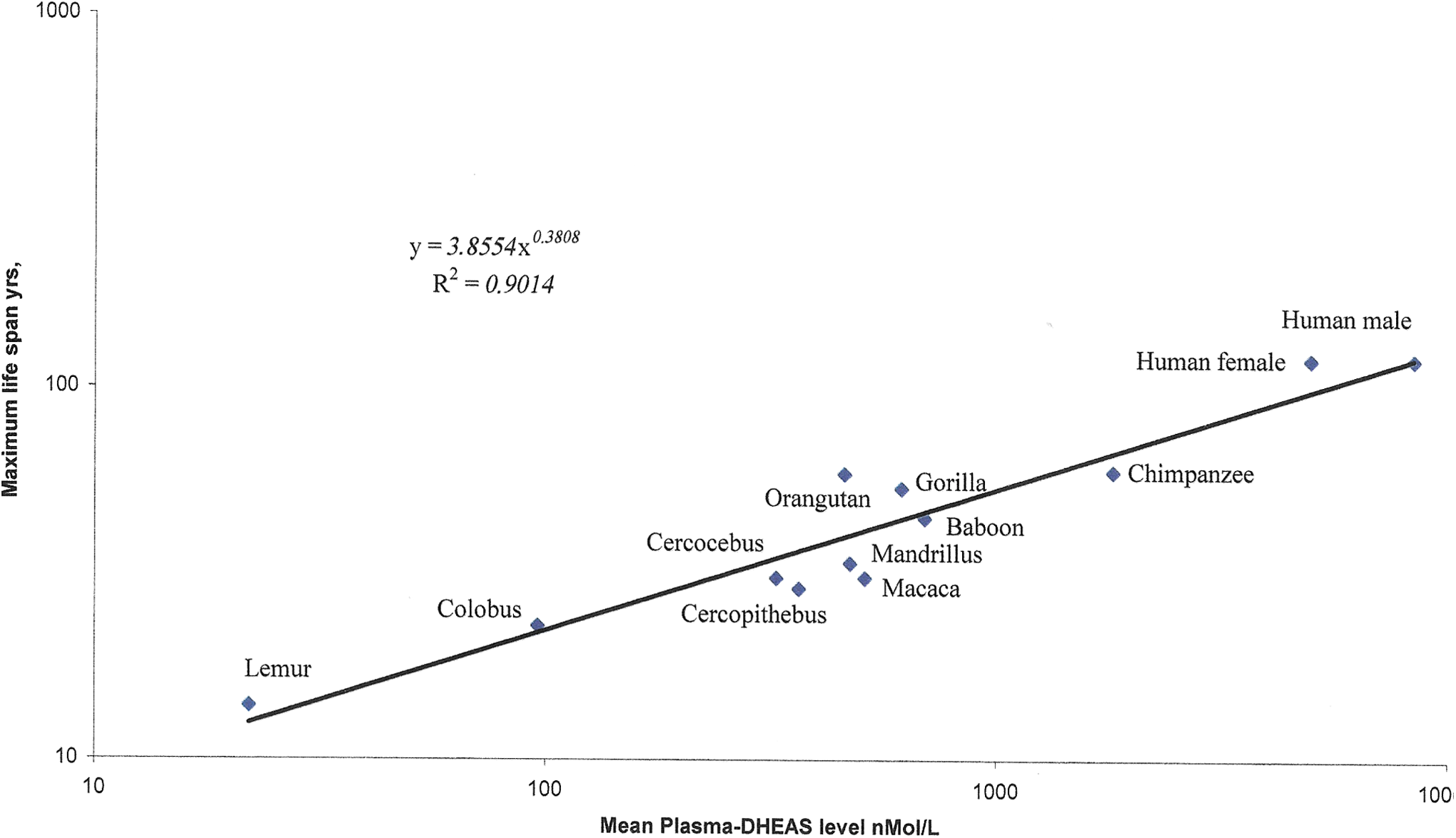
The correlation of the plasma dehydroepiandrosterone sulfate (DHEAS) level and maximum life span in primates. Data are from Table 1; scales are logarithmic. The regression equation defines a Pearson correlation coefficient of R
2=0.9014 with a two-tailed probability of 0.000240, indicating that the plasma level of DHEAS correlates with approximately 90% of the mechanisms contributing to the epigenetic evolution of primate longevity. Color images available online at
The maximum life spans of selected primates were derived from the AnAge database. 50 For non-human primates, the respective plasma-DHEAS levels provided are pooled values from equal numbers of healthy adult male and female animals reared at zoological institutions and obtained during the course of veterinary or routine medical examinations. 51 –53 The samples were likely taken during early working hours corresponding to the diurnal maximum of plasma-DHEAS in short-lived primates and are aimed at characterizing the evolution of adrenarche (young adults). Seasonal variations were not controlled for. The human data are reference values of normal young adult subjects (Tanner stage V). 11 DHEAS concentrations given in mass units (ng/mL) were converted to molar units (nmol/L) using the conversion factor 2.7. 54
p-DHEAS, plasma dehydroepiandrosterone sulfate.
It is interesting to note that chronic treatment with DHEA in the diet gave rise to an increase in the life span of mice by counteracting the development of specific diseases. Here, the effect of DHEA on the life span resembled that obtained by caloric restriction without any changes to food intake. 59 In contrast, treatment with DHEAS in the diet did not affect life span or cancer patterns in mice. 60,61 This difference in the biological effect of dietary supplementation with DHEA and DHEAS illustrates that DHEAS does not readily convert to DHEA. 62 Additionally, this difference suggests that the observed correlation of plasma DHEAS level and primate longevity reflects the hormonal effects of the DHEAS precursors rather than DHEAS itself, with an emphasis on DHEA and intracrine metabolites.
In any case, because DHEA stimulates chaperone expression and chaperones promote longevity, it seems reasonable to assume that the correlation of DHEAS with the life span of primates to some extent reflects the influence of DHEA on chaperone expression. Also, the decline in chaperone level and the increase in frailty during senescence probably to some extent reflect the concomitant decrease in DHEA promotion of chaperone expression.
DHEA supplementation during senescence,
A daily dose of 10–15 mg of DHEA sufficiently compensates for the age-related loss of DHEA secretion. 63,64 DHEA should preferably be administered using the percutaneous 65 or the trans-buccal route 66 or should be dissolved in sesame oil and given with meals, 64 all to avoid first-passage conversion to DHEAS in the liver. The long-term use of pharmacologic oral doses of DHEA (>50 mg/day) should be avoided due to the risk of developing liver tumors. 67 A future alternative to DHEA supplementation could be the use of TSPO ligands for the promotion of neurosteroid expression. 28,68
Conclusions
The level of DHEAS expression correlates with primate longevity. Contributing to this correlation may be the DHEA promotion of the expression of the molecular chaperones that are housekeeping constitutive or stress response proteins essential for the processes of proofreading, folding, and translocation of other polymers (proteins, RNA, and DNA), as well as for disaggregation or degradation of misfolded polymers. Also chaperone functions are of significance for homeostasis, immune response, and cancer resistance. Possibly the DHEA promotion of chaperone expression participates in the epigenetic evolution of primate longevity. DHEA supplementation during senescence has been suggested as a means for compensation of the age-related loss of DHEA secretion.
Footnotes
Author Disclosure Statement
No competing financial interests exist.