
Abstract
It was once suggested that adult or tissue-specific stem cells may be immortal; however, several recently published data suggest that their efficacy is limited by natural aging in common with most other somatic cell types. Decreased activity of stem cells in old age raises questions as to whether the age of the donor should be considered during stem cell transplantation and at what age the donor stem cells should be harvested to ensure the largest possible number of viable, functional, and non-altered stem cells. Although stem cells remain active into old age, changes in stem cells and their microenvironments inhibit their regenerative potential. The impact of aging on stem cell populations differs between tissues and depends on a number intrinsic and extrinsic factors, including systemic changes associated with immune system alterations. In this review, we describe key mechanisms of stem and progenitor cell aging and techniques that are currently used to identify signs of stem cells aging. Furthermore, we focus on the impact of aging on the capacity for proliferation, differentiation, and clinical use of stem cells. Finally, we detail the aging of embryonic, mesenchymal, and induced pluripotent stem cells, with particular emphasis on aging mechanisms and rejuvenation.
Introduction
I
Today, aging is still an unclear phenomenon in human biology. Moreover, there is sound scientific evidence that stem cells are aging. The latest findings raise new questions as to whether the age of the donor should be considered during the transplantation of stem cells and to define the best procedure for obtaining the largest possible number of stem cells in vitro. Because of the high expectations regarding the therapeutic application of stem cells, inevitably, in the context of normal aging, several important questions arise that we will try to answer in this review.
Mechanisms of Stem Cells Aging
There are several factors that contribute to stem cell aging—shortening of telomeres, de-repression of the gene locus INK4a/ARF and cell cycle regulators, accumulation of DNA damage, epigenetics, changes in mitochondrial structure, signaling pathways, as well as systemic factors.
Shortening of telomeres
Telomeres are nucleoprotein complexes at the chromosome ends. They consist of linear arrays of repeat sequences that are 5–15 kb in humans but considerably larger in mice. 4 In many human somatic tissues, a decline in cellular division capacity with age appears to be linked to the fact that the telomeres get progressively shorter as cells divide. The reason for this is the absence of the enzyme telomerase, which is normally expressed in certain adult stem cells and in germ cells (in testis and ovary). 5 Campisi suggested that in dividing somatic cells telomeres protect cells from uncontrolled cell division, as occurs in cancer. As time passes and the telomeres are shortened, tumor protection is reduced. 6 Moreover, it was found that stress, especially oxidative stress, has a great influence on telomerase shortening. 7
Defects in human telomerase function cause diseases with features of premature aging, including impaired regeneration of proliferative tissues. 8 The decline in tissue regeneration in response to telomere shortening has been related to impairments in stem cell function. Telomere dysfunction impairs stem cell function by activation of cell-intrinsic checkpoints and by the induction of alterations in the micro- and macro-environment of stem cells. 9 However, O'Connor et al. showed that telomerase activity in muscle stem cells is largely retained into old age, and that young and old muscle stem cells are likely to produce tissue with normal telomere maintenance when used in molecular and regenerative medicine approaches for tissue repair. 10 Accordingly, old muscle stem cells are likely to be suitable for regeneration in aged skeletal muscle. Nevertheless, telomere preservation is one of the key aspects of genomic integrity because their misfunction impairs regeneration and accelerates aging 11 (Fig. 1).

Factors affecting aging of stem cells—shortening of telomeres, accumulation of DNA damage, epigenetics, changes in mitochondrial structure, de-repression of the gene locus INK4a/ARF and cell-cycle regulators, signaling pathways, and systemic factors. ROS, reactive oxygen species; IR, ionizing radiation; ChM, chemical mutagens; MLH1, DNA mismatch repair protein Mlh1, mutL homolog 1; H4-K20, histone H4 lysine 20; FACE1, prenyl protein-specific endoprotease 1; HDAC, histone deacetylase; SIRT 1 and 2, sirtuins, class of III family of HDACs. Color images available online at
De-repression of the gene locus INK4a/ARF and cell cycle regulators
Damaged self-regeneration capacity of stem cells in the aging process is the result of an increased repression of the cell cycle (i.e., de-repression the activity of cyclin-dependent kinase inhibitor p16INK4a). It is considered that another cyclin-dependent kinase inhibitor, p19Arf, is responsible for the induction of cellular aging of stem cells (Fig. 1). An increased activity of the Hmga2 gene, involved in the modulation of transcription by chromatin structure modification, is responsible for de-repression of p16INK4a and p19Arf in old hematopoietic stem cells (HSCs). In all dividing cells, the rate of cell division is regulated by the cell cycle progression. The most important regulatory proteins in the cell cycle progression from the G1 in S phase are retinoblastoma family proteins (Rb). In the upstream Rb family of proteins in this regulatory system is p16INK4a, a cyclin-dependent kinase inhibitor, which leads to reduction of stem cell self-renewal in the course of aging, including neural and HSCs. Although p16INK4a plays a minimal role in the regulation of the cell cycle in young people, it has been proven that there is a de-repression of its activity in stem cells in elderly persons. 12,13 A similar situation applies to the second cyclin-dependent kinase inhibitor p19Arf (p14Arf in humans), which promotes p53 protein stability by inhibiting Mdm2-mediated degradation, although at the moment there are not enough details to prove that p19Arf negatively regulates stem cell function in aging tissues. 11
DNA damage
The accumulation of DNA damage in cells that occurs during the aging process is primarily the result of action of free radicals (reactive oxygen species, ROS), ionizing radiation, and chemical mutagens (Fig. 1). Parrinello et al. showed that provoking various forms of DNA damage triggers senescence in in vitro–cultured cells. 14 Failure of some genes, particularly those involving defects in DNA maintenance and repair functions, such as WRN, Ku80, and ERCC1, might contribute to accelerate aging. 15 The DNA damage through DNA double-strand breaks (DSBs) results in genome instability and cell death, advancing tumorigenesis and premature aging. 16 DNA DSBs in aging of stem cells is, by all accounts, tissue-dependent, and the aged cells, such as aged satellite cells in muscle, are unable to initiate regeneration due to cell cycle arrest resulting from accumulation of DNA DSBs. 16
Using the alkaline comet assay, Beerman et al. measured DNA damage and inferred that DNA damage accumulates in quiescent aged HSCs and is repaired upon cell cycle re-entry. After cell cycle re-entry, DNA damage profiles by alkaline comet assay in aged HSCs resemble those of young HSCs, but the proliferative potential of stem cells is decreased in aged HSCs. 17 In continuous DNA repair, constitutive suppression of the somatotropic axis inhibits aging, at least under laboratory conditions. 18 These observations have motivated researchers to consider DNA damage and DNA damage signaling as possible factors contributing to cellular senescence and stem cell aging. The age-dependent accumulation of DNA damage could provoke senescence of stem cells, especially induced pluripotent stem cells (iPSCs), and therefore reduce their functionality. 19
Epigenetics
Epigenetic signaling has an important role in tumorigenesis, but it can also be a significant factor of cellular senescence. 20,21 Several specific regions of the genomic DNA become hyper-methylated during aging; for example, there is an increase of methylcytosine within the ribosomal DNA clusters in livers of senescent rats, which could be associated with the decrease of RNA levels during aging. 22,23 Methylation of promoter CpG sites in non-tumorigenic tissues has been shown for a few genes, including insulin-like growth factor II (IGF2), estrogen receptor (ER), myogenic differentiation antigen 1 (MYOD1), and tumor suppressor candidate 33 (N33). Moreover, MLH1 and p14Arf are examples of colon cancer promoter hyper-methylation, which was observed more commonly in aged tissues. 22 It is still unknown if these histone-modifying enzymes are altered with aging.
The increase of trimethylated H4-K20 in aged-like cells has been caused by defects in the nuclear lamina, but there are not enough data on the molecular mechanism linking the nuclear lamins and the histone-modifying complex. However, there is a connection between modifications of the nuclear lamins and altered nuclear morphology. 20 In support of this, a recent study in mice reported that silencing of FACE-1 and metalloprotease involved in prelamin A proteolytic maturation are linked with premature aging and disruption of the nuclear integrity. 24 In recent years, it has be shown that H4-K20, an important epigenetic regulator, is potentially involved in aging.
Enzymes with histone deacetylase (HDAC) activity also have a role in aging. Among them, the most important are sirtuins. 25 –29 Histone acetylation controls chromatin structure and thus affects the regulation of gene expression (Fig. 1). Sirtuins, which belong to the class III family of HDACs, are nicotinamide adenine dinucleotide (NAD)-dependent and are involved in multiple cellular events, including control of cell cycle, mitosis, chromatin remodeling, and transcription. Sirtuins are important to the control of crucial metabolic pathways and the regulation of cell growth and cancer. The most explored sirtuins are SIRT1 and SIRT2, whose expression has been shown to be age dependent. 27 Furthermore the expression of these two sirtuins is changed in cancer cells, 28 and they are involved in the epigenetic silencing of telomeres, nucleolar ribosomal DNA (rDNA), and mating type loci and form clusters at the nuclear periphery. 27 The latest data indicate that the role of SIRT1 in growth and differentiation of mesenchymal stem cells decreases with age. 28,29
Mitochondrial dysfunction
Mitochondria were first implicated in the aging process for over 50 years ago by Denham Harman in his free radical theory of aging. 30 An important connection between accumulation of mitochondrial DNA (mtDNA) mutations and aging is suggested by many studies. 17,31 –36 The mitochondrion, or powerhouse of the cell, has its own genome, which encodes several key proteins required for the primary function of generating adenosine triphosphate (ATP) by oxidative phosphorylation, although a large number of additional proteins encoded by the nuclear genome are also required. An accumulation of mutations in mtDNA, which happens during aging, leading to respiratory chain dysfunction has been observed in several cell systems. Age-related increases in the frequency of cytochrome c oxidase (COX)-deficient cells, which are linked with mtDNA mutation, have been demonstrated in human muscle, 32 brain, 33 liver, 34 and gut. 35 Cells in which mtDNA mutations reach a high level are likely to suffer from impaired ATP production, resulting in a decline in tissue bioenergenesis.
Beside the primary mitochondrial lesions such as mtDNA mutation, secondary alterations in mitochondrial function driven by age-related cellular and metabolic changes may also contribute to the aging process. In this respect it is important to note several signaling pathways, such as liver kinase B1 (LKB1)-adenine monophosphate–activated protein kinase (AMPK) pathway and phosphoinositide 3-kinase (PI3K)-AKT pathway, and peroxisome proliferator activated receptor gamma co-activator 1 (PGC-1), a key regulator of mitochondrial energy metabolism. 36
Signaling pathways
Aging leads to decreased expression of epithelial growth factor receptors (EGFR) (Fig. 1) in neurogenic stem cells. Signaling via EGFR is important in maintaining the proliferative potential of progenitor cells in their neurogenic niche. Therefore, the decrease of neurogenesis during aging is a result of reduced number of cells, which express the components of specified signaling pathway. In neurogenic niches of stem cells in the aged brain, tumor necrosis factor-α (TNF-α), the endogenous ligand for EGFR, is expressed less. 37 Many receptor–ligand interactions that occur in stem cell niches can modify the functional activity. For example, the Notch-Delta pathway is important in maintaining the number of stem cells. In the case of muscle damage, increased expression of the Delta ligand causes increased proliferation and differentiation of satellite cells in muscle cells. However, upon injury, expression of the ligand Delta is reduced in aged satellite cells compared to young satellite cells. In contrast, parabiotic pairing shows that under the influence of factors from the blood of young animals, there is an increase in the expression of Delta ligand stem muscle cells of aged animals. In older animals increased activation of signal transduction via transforming growth factor-β (TGF-β)/pSAMD3 and the WNT pathway is observed, whereas the activity of inflammatory cells decreases in stem cell niches. 38,39
Systemic factors
In addition to the local signals, systemic factors also influence the aging of stem cells. 35 In old mice, increased expression of TGF-β, which accumulates in aged muscle, hinders the proliferation and satellite cell regeneration. 40 Chronic elevation of inflammatory mediators in aged tissue may also contribute to the aging process of tissue, as well as stem cells. 41 All effects of the nuclear factor-κB (NF-κB) signaling transduction pathway are not yet known, but it is known that this pathway mediates chronic inflammation in aged skin, skeletal muscle, bone, and nervous system. 42 Quite a new finding is that aging cells may contribute to chronic inflammation, because inflammatory factors, growth regulators, proteases, and other signaling molecules 43 affect neighboring cells in the local environment and promote senescence and inflammation.
Signals of the microenvironment are transferred between stem cells, and their niches made of the extracellular matrix, stem cells in direct contact with other cell types, and soluble factors that are secreted or concentrated locally. Conditions in the niche depend on the systemic milieu and dynamically changes to regulate stem cell function, especially during the aging process. Changes within the stem cell microenvironment that occur with aging are well documented and, unquestionably, these extrinsic changes affect stem and progenitor cell behavior. 36 Conboy et al. 44 demonstrated that the changes in the systemic environment reduce the regeneration potential of satellite cells in muscle with aging. Using parabiotic pairings, via common circulation, between young and old mice, they determined that factors in the young circulatory system were able to recover muscle tissue regeneration after injury in old mice.
How to Identify Signs of Aging Stem Cells?
To have an effectual therapeutic administration of stem cells, it is necessary first of all to standardize the methods of isolation and implement a cell quality control protocol. However, even in highly standardized culture conditions, the phenomena of aging should not be neglected during long-term cell culture. On the other hand, it is not completely clear if the same therapeutic effect can be obtained when using stem cells from old donors as compared to younger people. Therefore, we would like to describe the methods that have been developed to monitor stem cell aging.
The most important criteria to bear in mind are summarized in Fig. 2: (1) Reducing the number of passages; (2) reduction in cumulative doubling cell population; (3) accumulation of senescence-associated lysosomal β-galactosidase (SA-β-gal); (4) minimize changes in karyotype; (5) the shortening of telomere length; and (6) changes in gene expression. 45 It should be noted that there are some differences in aging between embryonic (ESCs) and mesenchymal stem cells (MSCs). Signs of aging ESCs cannot be monitored through age of the embryos. 46
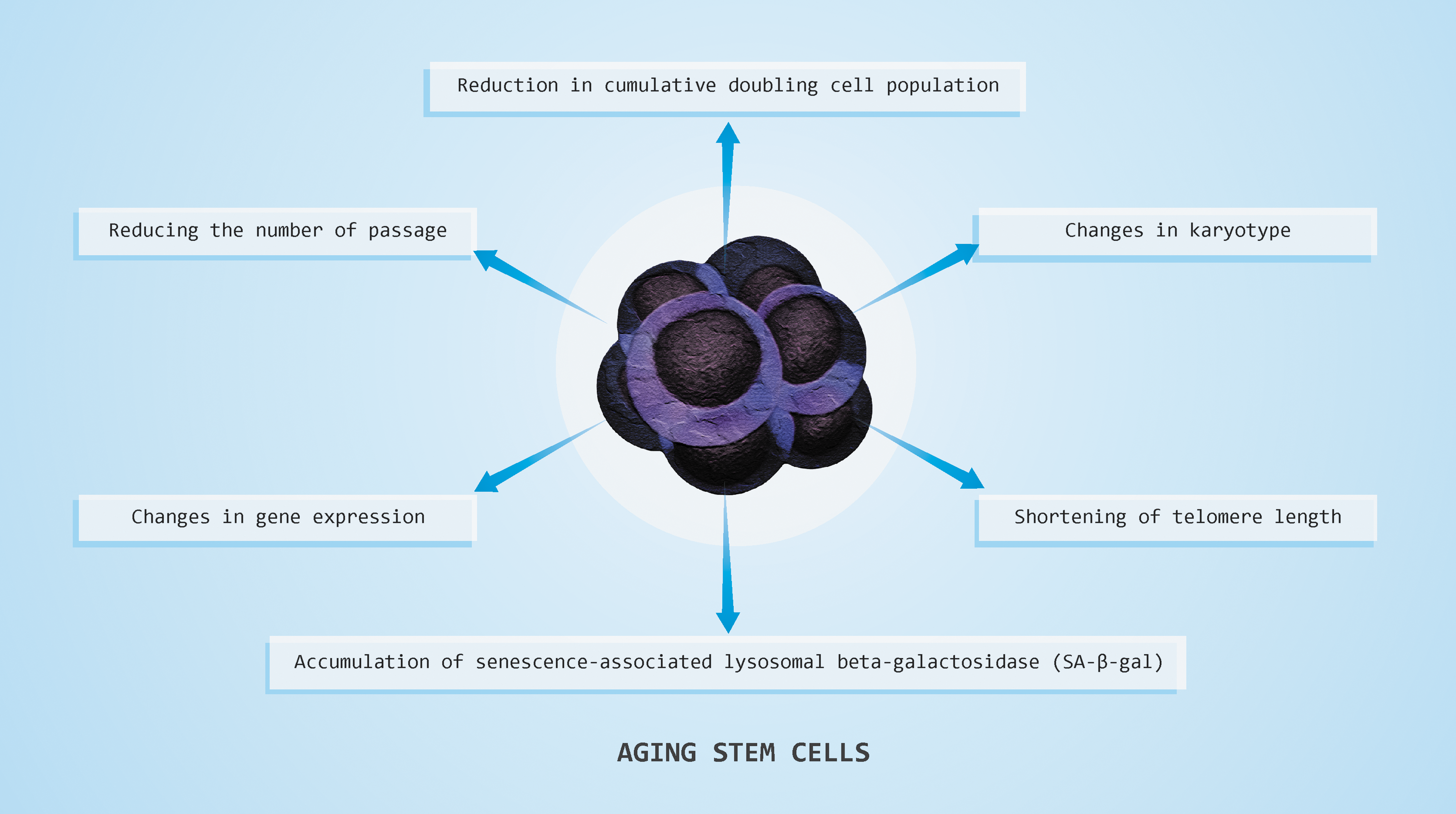
Consequences of stem cell aging—reducing the number of passages, reduction in cumulative doubling cell population, changes in karyotype, shortening of telomere length, accumulation of senescence-associated lysosomal β-galactosidase, and changes in gene expression. Color images available online at
Required methods in monitoring aging stem cells are assessment by the number of passages and accurately calculation of n the number of cumulative population doublings. MSCs are relatively stable with regard to karyotype aberrations, and these changes are not seen in normal MSC aging, as opposed to ESCs. Measuring the activity of SA-β-gal
Therefore, we should not forget that aging-induced stress signals via such mechanisms may be independent of the cell cycle and shortening of telomeres, so we could have aging cells with apparently normal telomeres. Regarding gene expression, today the greatest importance is attached to the increase in expression of PARG1 (ARHGAP29), CDKIN2b, and p16INK4a , and reduced gene expression of pleiotropin (PTN) and MCM3, which is necessary to determine the quantitative RT-PCR analysis. 42 With regard to the question of how to possibly stop cell aging, it is necessary to remind ourselves that excessive activation of certain cell signaling pathways exhaust stem cells pools. For example, the pathway of cell signaling that induce aging of epidermal cells is Wnt mediated by mTOR. In this way, among the pathways of cell signals that induce aging of epidermal stem cells should be counted time Wnt mediated by mTOR. Likewise mTORC1 hyper-activation leads to hyper-proliferation, and then very quickly to the reduction in the number of HSCs in their niche. On the contrary, stem cell aging can be prevented by using pharmaceuticals. For instance, activation of PI3K-AKT-FoxO delays aging of neurogenic progenitor cells. 49
The Impact of Aging on the Capacity for Proliferation and Differentiation of Stem Cells
The process of aging alters the immune system as a whole, as evidenced by decreased primary and secondary immune responses and increased incidence of inflammatory pathologies. 50 This concept of “immunosenescence” can modify overall immunity. Accordingly, the effect of immunosenescence on stem cells, progenitor cells, and especially primary MSCs may alter their proliferation, differentiation, and therapeutic potential, and, as many of their intrinsic beneficial functions deteriorate with age, other misbalanced regulatory events lead to alterations in their immunoregulatory potential. 51 Aging influences the secretion of numerous paracrine signals, thus disturbing the endogenous regeneration of solid organs. 52 Therefore, application of stem cells together with some factors, such as cytokines, biomatrices, or mural cells, could transiently create a healthy paracrine environment and enhance engraftment and differentiation of transplanted cells. 53 The ex vivo use of growth factors, microRNAs (miRNAs), and cytokines can modulate the function of stem cells, as has been shown for interferon-γ, which substantially enhances the immune suppressive function of mesenchymal stromal cells to suppress autoimmune and alloimmune diseases. 54 Similar treatment of pro-angiogenic cells, such as endothelial cells, with various miRNAs or antagomirs can also enhance their in vivo survival and homing and angiogenic activity or reverse their senescence phenotype. 55 Finally, it is important to note that the concept of “immunosenescence” is not relevant to ESCs.
Aging of ESCs
Xie et al. 46 investigated changes in molecular profiles associated with young (<60 passages) and old (>120 passages) cells of the H9 human ESC (hESC) cell line as well as young (<85 passages) and old (>120 passages) cells of the PKU1 hESC line (Table 1). Their results show that the morphology, expression of stem cell markers, and telomerase activity do not differ significantly between young and old passage cells. Cells from both age groups were also shown to differentiate into derivatives of all three germ layers upon spontaneous differentiation in vitro. Interestingly, mitochondrial dysfunction was found to occur with prolonged culture. Old-passage cells of both the H9 and PKU1 lines were characterized by higher mitochondrial membrane potential, larger mitochondrial morphology, and higher ROS content than their younger counterparts. These findings suggest that prolonged culture of hESCs may negatively impact mitochondrial function and possibly affect long-term pluripotency. 46 These findings are related to in vitro growth conditions, whereas under in vivo conditions ESCs cells are generally considered to be resistant to replicative senescence. 46
Our work 56 showed for the first time an important role for telomerase reverse transcriptase (TERT) in differentiation capacity and maintenance pluripotency of human ESCs in vitro, as well as in cell cycle regulation. Up-regulation of TERT expression was associated with increases in cyclin D1 and CDC6 expression, as well as hyper-phosphorylation of Rb and enhanced proliferation and colony-forming ability of human ESCs. ESCs showed shortening of telomeric DNA as a result of loss of telomerase activity. Down-regulation of TERT caused loss of pluripotency and human ESC differentiation to extraembryonic and embryonic lineages. 56
Aging of MSCs
MSCs, with their multi-potent nature, have advantages over ESCs in terms of their availability and feasibility. However, their proliferative activity is destined to slow by replicative senescence, and the limited proliferative potential of MSCs not only hinders the preparation of sufficient cells for in vivo application, but also draws a limitation on their potential for differentiation. 57 The low frequency of MSCs in bone marrow 58,59 necessitates their in vitro expansion prior to clinical use. It is known that when the cells become senescent, they are unable to proliferate further. Ex vivo comparison of MSCs isolated from young and aged donors 60 and in vitro analysis of isolated MSCs over several population doublings 61,62 are the most utilized experimental aging models. A major flaw of these models is their incapacity to completely reproduce the events of natural aging observed in vivo. 63 –65 Age-related changes in MSCs include morphological changes, loss of proliferation and differentiation potential, and thus therapeutic ineffectiveness 66 –69 (Table 1). Geißler et al. 69 showed that long-term in vitro culture, but not in vivo aging, compromises the osteogenic and adipogenic differentiation capacity of MSCs and alters their morphology, susceptibility to senescence, and mitochondrial function. Accordingly, transcriptome analysis revealed that in vivo and in vitro aging results to a large extent in divergent changes at the molecular level. Thus, independent from donor animal age, in vitro aging of MSCs seems to result in complete loss of their progenitor characteristics. 69 Other experiments in mice also suggest that the aged milieu can suppress the functionality and potential of adult stem cells. 44,70 –72 Serum isolated from old compared to young mice can suppress the in vitro proliferation of stem cells. Unlike MSCs, ESCs have at least some capacity to antagonize this effect, suggesting that they produce some factors that can neutralize the anti-proliferative factors in aged serum. 70 –72 In recent years, many studies have demonstrated that MSCs isolated from older donors show many qualitative and quantitative changes compared to young donors. 73 –79
Huang et al. 73 investigated the biological characteristics of human bone marrow MSCs of different age donors. In their experiment, samples were divided into four groups: A (fetuses), B (0- to 20-year-old donors), C (20- to 40-year-old donors), and D (donors older than 40). The ability of expansion decreased after passage 15 for group A, passage 10 for group B, and passage 8 for groups C and D; all of the other results showed that cells were similar in morphology, antigenic phenotype, differentiation potential, and cell cycle.
Mareschi et al. 74 investigated MSCs from bone marrow (BM) of eight pediatric and 17 young adult donors to analyze the growth kinetic, immunophenotype, telomere length, and karyotype during ex vivo expansion. No morphological differences were observed in the MSCs isolated from BM, but their cell growth was strictly related to the donor's age. Cumulative population doubling of MSCs isolated from pediatric donors is almost twice better than MSCs isolated from young adult donors after 112 days. Expression of antigens in the MSCs isolated from two groups until 10th passage (77 days) showed no significant difference between young and old MSCs. Schipper et al. 75 compared MSCs isolated from subcutaneous adipose tissues in 12 female patients classified into three groups: I (25–30 years), II (40–45 years), and III (55–60 years old). They noted differences in cell proliferation related to age. Younger patients had increased peroxisome proliferator-activated receptors-γ (PPARγ) agonist expression, unlike older patients, who showed different and variable expression. Besides, age-related changes in function of MSCs, such as reduction of proliferation, are registered. Nayan et al. investigated the proliferative capability and therapeutic potential of young MSCs, which had significantly higher proliferation potential than old MSCs at every passage. 76 They proved that old MSCs exhibited decreased cell viability, proliferation, and differentiation potentials, and that a better outcome may be achieved if aged recipients are given MSCs obtained from young donors rather than using their own autologous MSCs.
It is assumed that organismal aging is linked to diminished organ repair due to reduced functional capacity of tissue resident stem cells and that such cells residing in the elderly are subjected to age-related changes and thus contribute less to tissue rejuvenation. Similarly, age-related diseases such as diabetes and heart failure also negatively impact the function of endogenous progenitor cells. 77 The recent study demonstrated the effect of donor age on the cell expansion and differentiation potential of adipose tissue-derived MSCs and represents one of the most comprehensive analyses providing insight into whether cell-based therapies will be negatively affected by donor age. 79 Analysis indicated that the overall yield of nucleated cells was significantly and negatively affected by donor age. Similar observations have been reported of circulating endothelial progenitor cells. 78
These results indicate that age-related changes in MSCs should be taken into account whenever these cells are considered for clinical applications in the elderly. Although MSCs from all age groups had the ability to form colonies, MSCs from younger donors produced more colonies containing larger numbers of cells. 79 Except for disagreement about morphological changes with Stolzing and her collaborators, 66 this study confirmed all previous serious findings in the literature on the impact of aging on the differentiation and therapeutic potential of MSCs.
Aging of iPSCs
The story of the aging of stem cells does not bypass iPSCs. The technique of obtaining iPSCs opens new and exciting possibilities in medical research, especially personalized cell therapies for the therapy of various diseases. 80 To treat damaged tissues or repair organs in elderly patients, it will be necessary to establish iPSCs from their tissues. However, the important question is whether iPSCs retain epigenetic memory characteristics of their tissue of origin that may affect their differentiation and therapeutic potential. iPSCs reprogrammed from senescent and centenarian human fibroblasts have reset the expression of genes associated with mitochondrial metabolism and oxidative stress. Mitochondrial morphology and distribution were also both comparable to ESCs. 81 In iPSC clones derived from human dermal fibroblasts, mitochondrial biogenesis and ROS stress defense mechanisms were analogous to those observed in ESCs. 82 Resetting of metabolism and restructuring of mitochondrial signature have also been observed 83,84 and suggest that human iPSCs exhibit a metabolic signature that, while not completely identical to ESCs, clusters more closely with ESCs than with differentiated cells.
Although different reports have been published regarding the extent of rejuvenation, these data clearly demonstrate that the reprogramming process resets mitochondria and related metabolic and stress mechanisms to a more youthful state. Our group has generated experimental data 85,86 that support a contribution of age-related epigenetic changes to the overall aged phenotype of adult, tissue-specific stem cells. In our previous research, we stated that epigenetic change is not the only causative factor in aging, for there can be little doubt that all mitotically active cell types, even relatively quiescent stem cells, must accumulate some mutations as a consequence of DNA replication. 85,86 Further studies are required to illuminate the extent of this rejuvenation and why varied results have been obtained.
Rejuvenation of Aged Cells
Cell-based therapies using the body's own stem cells and growth factors have recently been used as an alternative therapeutic strategy to repair damaged tissue, including tissue rejuvenation. Stem cells may exert their beneficial effects on tissue regeneration through complex paracrine mechanisms in addition to their proposed direct cellular effect. 87 Rejuvenation of adult tissue is regulated by a population of stem cells that, by undergoing self-renewal, maintain their own pool and give rise to differentiated progenitors that replace cells used up during the life cycle.
A study of mechanisms of molecular aging improves our understanding of stem cell behavior and reveals therapeutic methods for rejuvenating tissue. Acquired epigenomic changes that are dynamically maintained by a balance among chromatin-remodeling complexes are reversible. Considering the influence of cell extrinsic factors on the epigenome and the reversibility of chromatin modifications, epigenomic changes can be a significant factor of aging. 88 It is demonstrated that the phenotype of the old stem cells can be changed if they are exposed to a “young” microenvironment. Extracellular matrix from young donors rejuvenated old stem cells in expansion in vitro, as evidenced by increased cell proliferation and resistance to apoptosis. 38 Furthermore, the elimination of tissue-specific senescent cells could also serve to promote tissue regeneration, because dysfunctional senescent cells impede regenerative processes, not only by occupying tissue compartments but also by secreting harmful molecules.
Baker et al. made use of a biomarker for senescence, p16Ink4a, to design a novel transgene, INK-ATTAC, for elimination of p16Ink4a-positive senescent cells upon administration of a drug. They showed that INK-ATTAC removes p16Ink4a-positive senescent cells upon drug treatment, thus attenuating progression of already established age-related disorders and preventing or delaying tissue dysfunction and extending health span. 89 Moreover, it is important to note that the elimination of the technical constraints of reprogramming somatic cells also leads to rejuvenation of aged cells, such as differential gene expression profiles (genetic signatures), use of potentially harmful genome integrating viruses, and induction of carcinogenesis, which remains a pending threat. 90 To this end, several numbers of strategies were recently developed, including using the non-integrating adenovirus and Sendai virus, plasmid vectors, recombinant proteins, or synthetic mRNA delivery. 91
Lapasset et al. 81 showed that it is possible, using an adequate reprogramming strategy, to efficiently reprogram senescent cells and cells derived from very old individuals into iPSCs and that cellular senescence and aging are not barriers to reprogramming toward pluripotency. Crucially, they also demonstrated for the first time that, when re-differentiated back into fibroblasts, cells have rejuvenated extended life spans and the characteristics of young proliferative embryonic fibroblasts; thus, these cells have been completely rid of their former aged cellular phenotype. Their demonstration of the reversibility of major aspects of the cellular aging physiology provides a totally unexpected insight into the perceived importance of epigenetic modifications in aging and provides a new paradigm for cell rejuvenation. 81 Although age-associated DNA methylation changes are counteracted in iPSCs, it is important to say that epigenetic rejuvenation does not counteract the mutations that may accumulate during in vitro culture. 92,93 In view of this, we can expect to witness soon the clinical application of “epigenetic drugs,” such as the HDAC inhibitor valproic acid in the treatment of aging and cancer. 94
Conclusion
This review has summarized mechanisms that regulate declines in stem cell function with age. Taken together, stem cell aging is affected not only by their origin and growth condition but also by many different cell pathways that disrupt stem cell function. Because of their potential for proliferation and differentiation, as well as their immunomodulatory characteristics, stem cells are promising therapeutic agents for the therapy of various diseases and the fight against aging. Great progress has been made in the last few years in understanding how cells limit their proliferative potential and in linking these mechanisms to aging. However, there are still several problems, including conflicting data regarding aging stem cells and their long-term efficacy. The topic of aging stem cells will continue to be current, especially if we know that human life span will continue to increase.
Footnotes
Acknowledgments
This study was supported by Serbian Ministry of Science (Grants ON175069, ON175103 and ON 175061). We appreciate and acknowledge the generous assistance of Mr. Dzenan Hajrovic, who contributed to the creation of figures in this article.
Author Disclosure Statement
No competing financial interests exist.