
Abstract
Anemone flaccida Fr. Schmidt is used in the clinical compound prescription for the treatment of rheumatoid arthritis (RA) in China and has the traditional use of draining dampness, diminishing swelling, and relieving pain. Total saponins (TS) are the characteristic components and also the main active ingredients of A. flaccida. Previous reports indicated that TS possess anti-inflammatory and immunoregulatory properties; however, the effects of TS on bone destruction of RA have not been evaluated. In this study, our data first showed the therapeutic effects of TS on severity of arthritis and arthritis progression in collagen-induced arthritis (CIA) rats. Then, by microfocal computed tomography (CT) quantification, TS significantly increased bone mineral density, bone volume fraction, and trabecular thickness and decreased trabecular separation of inflamed joints both at peri-articular and extra-articular locations. TS also diminished the level of the bone resorption marker CTX-I and simultaneously increased the bone formation marker osteocalcin in sera of CIA rats. Interestingly, TS prevented bone destruction by reducing the number of osteoclasts in inflamed joints, reducing the expression of receptor activator of nuclear factor-κF (RANK) ligand (RANKL) and RANK, increasing the expression of osteoprotegerin (OPG), at both mRNA and protein levels, and decreasing the ratio of RANKL to OPG in inflamed joints and sera of CIA rats. This was further confirmed in the co-culture system of human fibroblast-like synovial and peripheral blood mononuclear cells. In addition, TS inhibited the levels of pro-inflammatory cytokines implicated in bone resorption, such as interleukin-1β (IL-1β), tumor necrosis factor-α (TNFα), IL-6, IL-17, and IL-23 in sera and joints. These findings offer convincing evidence that TS attenuate RA partially by preventing both focal bone destruction and systemic bone loss. This anti-erosive effect results in part from inhibiting osteoclastogenesis by regulating the RANKL/RANK/OPG signaling pathway. The suppression of systemic and local pro-osteoclastogenic cytokines by TS was also highly effective.
Introduction
R
Osteoclasts, the primary bone resorptive cells, are abnormally activated in RA to cause local bone destruction and systemic osteoporosis. 13 –15 It has been demonstrated that osteoclasts are located mainly in the synovial inflammatory tissue. Bone destruction occurring in RA belongs to osteoclast-mediated bone destruction that is regulated by the receptor activator of nuclear factor-κB (RANK) ligand (RANKL). 16 During the pathological development of RA, the RANKL/RANK/osteroprotegerin (OPG) signaling pathway plays a crucial role in the process of bone destruction. 17 RANKL induces osteoclast-mediated bone destruction, and OPG protects against bone destruction by inhibiting the binding of RANKL with its receptor RANK. 18 Bone resorption is regulated locally by the relative levels of expression of RANKL and OPG, and, this signal pathway is up-regulated by numerous pro-inflammatory cytokines involved in the pathogenesis of RA. Therefore, osteoclasts and the RANKL/RANK/OPG signaling pathway have been considered as a potential target for preventing systemic joint destruction in RA patients.
There is a growing interest in using medicinal plants for prevention and treatment of bone disorders, including RA. 19 The plant Anemone flaccida Fr. Schmidt (Ranunculaceae) is widely distributed throughout southern China and is traditionally used for draining dampness, diminishing swelling, and relieving pains. The dry root of A. flaccida Fr. Schmidt, commonly known as “Di Wu” (Chinese name), is used extensively as a folk medicine in the clinical compound prescription for the treatment of rheumatism, arthralgia, and cancer. 20,21 Previous phytochemical and pharmacological studies showed that triterpenoid saponins are the main chemical components of this plant, 22 –25 and they possess anti-inflammation, immunoregulation, analgesia, and anti-tumor properties. 20,21,26,27 Moreover, the total saponin (TS) from the rhizomes of A. flaccida is undergoing a phase III clinical trial in China for the treatment of RA because of its significant anti-inflammatory and immunomodulatory effects. 20,26
In our recent study, TS has been demonstrated to abrogate osteoclast differentiation and bone resorption by inhibiting the activation of RANKL-induced NF-κB, JNK, and p38 MAPKs in Raw 264.7 cells. 28 However, its action on bone destruction of RA has not been evaluated. Because the bone destruction that occurs in RA belongs to osteoclast-mediated bone destruction that is regulated by the receptor activator of RANKL, here we investigated the effect of TS on bone destruction of RA and the possible mechanism associated with its action on osteoclastogenesis regulated by the RANKL/RANK/OPG signaling pathway in the collagen-induced arthritis (CIA) model and in vitro.
Materials and Methods
Preparation of herbal extraction
The rhizome of A. flaccida Fr. Schmidt was collected from Jiufeng County of Hubei Province, China. TS were isolated from A. flaccida and prepared as previously reported. 21,27 TS were dissolved in dimethyl sulfoxide (DMSO) and diluted with distilled water or phosphate-buffered saline (PBS) to the proper concentration before use in all subsequent experiments.
Induction of CIA and TS treatment
Eighty male SD rats (8–12 weeks old) were obtained from the Experimental Animal Center, Academy of Military Medical Sciences (no. SCXK-[Jun] 2007-004). They were kept in a temperature-controlled environment (22 ± 2°C), at 55 ± 5% relative humidity with a 12 hr:12 hr light–dark cycle and fed with standard chow, for at least 1 week before any manipulations. All procedures involving animal welfare were reviewed and approved by the ethical committee of China Academy of Chinese Medical Sciences.
CIA was induced as previously reported. 29 Briefly, bovine type II collagen (Chondrex, Redmond, WA) was dissolved in 0.01 M acetic acid overnight at 4°C and emulsified in an equal volume of incomplete Freund's adjuvant (Chondrex). The rats were immunized intradermally at the base of tail with 0.1 mL of emulsion containing 100 μg of type II collagen. On day 7 after the primary immunization, the rats were boosted like the first time.
Methotrexate (MTX) was purchased from Sigma (St. Louis, MO) and dissolved in 0.05% DMSO. TS and MTX were delivered by oral administration intragastrically using syringe feeding. Treatment was given daily for a period of 28 days from day 1 after primary immunization. SD rats were divided separately into six groups randomly with an equal number (n = 12) in each group. The normal control (control), the CIA model (vehicle), and CIA rats were treated with TS (15 mg/kg, 30 mg/kg, and 60 mg/kg, respectively), and CIA rats were treated with MTX (0.2 mg/kg).
Arthritis evaluation
Rats were observed once every 3–4 days after primary immunization. Arthritis severity was evaluated using arthritis scores and was performed by two independent, blinded observers. All four limbs of the rats were evaluated according to a visual assessment of inflammation or swelling and scored from 0 to 20 as in the reported scoring system. 30 The arthritis score was the total of the scores for all four limbs (maximum possible arthritis score 80). The incidence of arthritis was defined as the percentage of animals within each group exhibiting any sign of disease regardless of severity. In addition, the number of arthritic limbs of individual rats was counted and added to represent the number of arthritic limbs in a group. The percentage of arthritic limbs in a group was calculated as previously described. 31
Histopathology and tartrate-resistant acid phosphatase staining
Rats were sacrificed by cervical dislocation on day 28 after the first immunization. Both hind limbs, including the paws, ankles, and knees, were dissected, fixed immediately for 24 hr in 4% paraformaldehyde, decalcified in 10% EDTA for up to 1 month at 4°C, and embedded in paraffin. Tissue sections (4 μm) were mounted on common slides for staining with Hematoxylin & Eosin. All sections were randomized and evaluated by two trained observers who were blinded to the treatment groups and the arthritis severity of each mouse. Minor differences between observers were resolved by mutual agreement. The data were expressed as mean synovial mononuclear infiltrate, synovial hyperplasia, synovial fibrosis, pannus extension, cartilage erosion, and bone erosion scores. All scores were based on a scale of 0–3, as previously described. 32
To identify osteoclasts, sections were stained for tartrate-resistant acid phosphatase (TRAP) using an acid phosphatase kit (Sigma). TRAP-positive multinucleated cells that contained more than three nuclei were identified as osteoclasts, and these were counted by light microscopy.
Microfocal computed tomography
Three-dimensional reconstruction and sagittal images of the right knee joints were obtained by microfocal computed tomography (micro-CT; Skyscan 1174, Bruker micro-CT, Kontich, Belgium) at the day 28 after first immunization. Briefly, after the rats were killed, the hind limbs were removed and fixed in 4% paraformaldehyde for 24 h. The samples were scanned with micro-CT. Using the constructed sagittal image from micro-CT scanning data, bone morphometry analysis of the distal femoral epiphysis excluding the cortical bone was performed in the measurement area facing the articular cavity from the beginning of growth plate. Bone volume (BV)/total volume (TV) ratio, bone surface (BS)/BV ratio, trabecular thickness (Tb.Th), trabecular number (Tb.N), trabecular separation (Tb.Sp), and bone mineral density (BMD) at the femoral spongiosa were measured using an image analyzing system (CTAn V 1.13 software, Bruker micro-CT). Meanwhile, a core of 200 slides, each section 11 μm thick (7 mm long) at the tibial secondary spongiosa, was also used for bone morphometry evaluations.
Osteoclast formation in co-culture system of synovial fibroblasts and peripheral blood mononuclear cells
The co-culture system of human fibroblast-like synovial cells (HFLS) and peripheral blood mononuclear cells (PBMCs) was constructed according to the protocol of Nakano et al. 33 Briefly, HFLS were purchased from Cell Applications (San Diego, CA) and routinely cultured in synoviocyte growth medium (Cell Applications). Blood was collected from healthy volunteers, and PBMCs were isolated by centrifugation over Ficoll/Paque at 400 × g for 30 min. After isolation, PBMCs (2 × 105 cells/well) were re-suspended in μ-minimum essential medium (α-MEM) containing 10% fetal calf serum (FCS) and 50 ng/mL of macrophage colony-stimulating factor (M-CSF, PeproTech Inc. Rocky Hill, NJ) and then seeded in 24-well tissue culture plates. Three days later, adherent cells were used for subsequent co-cultures. HFLS were then added into 24-well tissue culture plates (2 × 104 cells/well) with PBMCs and co-cultured for further 9 days in α-MEM containing 10% FCS, 50 ng/mL of M-CSF, 10−7 M 1,25-dihydroxyvitamin D3 (1,25[OH]2D3) and/or TS (0.1, 0.5, 2.5 μg/mL, respectively). Some dishes were used for TRAP staining.
RNA isolation and real-time PCR
The left front limbs (including the paw and ankle) were dissected from rats on day 28, snap-frozen in liquid nitrogen, ground into powder, and homogenized. This procedure was done under RNase-free conditions. The RNA isolation and real-time PCR assay were carried out following the protocol of our previous study. 31,34,35 Briefly, total RNA was extracted with TRIzol® reagent (Invitrogen, Carlsbad, CA) from the tissue homogenates according to the manufacturer's instructions. The total RNA (1 μg) was reverse transcribed to cDNA using the QuantiTect Reverse Transcription Kit (Qiagen K.K., Tokyo, Japan) according to the instruction manual. The specific transcripts were quantified by quantitative real-time PCR using QuantiTect SYBR Green PCR Kit (Qiagen K.K.) and analyzed with an ABI 7500 Real-Time PCR system (Applied Biosystems, USA). Gene specific primers were used for RANK (forward, 5′-GTC TGC AGC TCT TCC CTG AC, and reverse 5′-GAG GAG CAG GAC GAT GAG AC-3′), RANKL (forward, 5′–ACC AGC ATC AAA ATC CCA AG-3′, and reverse, 5′-TTT GAA AGC CCC AAA GTA CG-3′), OPG (forward, 5′-GTT CTT GCA CAG CTT CAC CA-3′, and reverse, 5′–AAA CAG CCC AGT GAC CAT TC-3′), and GAPDH (forward, 5′–ACC CTA AGG CCA ACC GTG AAA AG-3′, and reverse, 5′–CAT GAG GTA GTC TGT CAG GT-3′). The mRNA levels of RANKL, RANK, and OPG were normalized to the glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA level. PCR was performed as 40 cycles at 94°C for 15 sec, 55°C for 30 sec, and 72°C for 30 sec. The relative mRNA expression was calculated with comparative cycle threshold (Ct) method.
Enzyme-linked immunosorbent assay
On the 28th day following the first immunization, right front limbs (including the paws and ankles) were amputated, cut into small pieces, and homogenized in 100 mg of tissue/1 mL of lysis buffer (physiological saline containing 2 mM phenylmethylsulfonyl fluoride and 1 mg/mL of aprotinin, leupeptin, and pepstatin A for cytokines). The protein concentration in the homogenate was determined with a BCA Kit (Pierce, USA). Sera were separated. The concentrations of interleukin-1β (IL-1β; R&D Systems, Minneapolis, MN), IL-6 (R&D), IL-17 (R&D), IL-23 (R&D), and tumor necrosis factor-α(TNF-α; R&D) in the supernatants of joints and sera, carboxy-terminal telopeptide α1 chain of type I collagen (CTX-1) (RatLaps; Nordic Bioscience), and osteocalcin (Nordic Bioscience) in sera were measured by enzyme-linked immunosorbent assay (ELISA) according to the manufacturer's protocol, and absorbance was measured at 450 nm. Conditioned media from HFLS or PBMCs cultured in the presence of TS (0.1, 0.5, 2.5 μg/mL, respectively) for different days (3, 6, and 9, respectively) were obtained and stored at −80°C until use. At the 9th day, the cells were lysed and collected. The supernatant and intracellular amounts of RANKL (R&D) and OPG (R&D) in HFLS and RANK (R&D) in PBMCs were detected by ELISA. All of the in vitro experiments were done in triplicate.
Western blot
The protein expression levels of RANK, RANKL, and OPG in left front limbs (including the paws and ankles) were detected by western blot analysis. The protocol and semi-quantitative analysis were carried out as in our previous study. 34 The following antibodies were used: RANK antibody (rabbit antibody, dilution 1:50; Cell Signaling Technology, Inc., Danvers, MA), RANKL antibody (rabbit antibody, dilution 1:100; Millipore Corporation, Billerica, MA), OPG antibody (rabbit antibody, dilution 1:100; Santa Cruz Biotechnology, Inc., Santa Cruz, CA), and GAPDH antibody (rabbit antibody, dilution 1:200; Santa Cruz).
Statistical analysis
The software of SPSS version 13.0 for Windows (SPSS Inc, Chicago, IL) and SAS 9.1 (SAS Institute, Cary, NC) was used for statistical analysis. Continuous variables were expressed as
Results
TS prevents arthritis progression and decreases severity of arthritis in CIA rats
To investigate the effect of TS on arthritis, the CIA model in SD rats was used. Oral administration of TS once a day started from day 1 to day 28 after initial immunization. As shown in Fig. 1A, TS dose-dependently interfered with increasing of arthritis scores in CIA rats, with significant suppression observed at both the 30- and 60-mg/kg doses. Consistent with the clinical scoring, the assessment of arthritis incidence (Fig. 1B) and the percentage of arthritic limbs (Fig. 1C) also showed TS to be highly effective. In groups receiving TS (30 and 60 mg/kg), the rate of incidence and the percentage of arthritic limbs were markedly reduced from day 15 after the first immunization. Macroscopic evidence of arthritis, such as erythema or swelling, was markedly observed in vehicle-treated CIA rats, whereas a dose of 60 mg/kg TS significantly attenuated arthritis severity in CIA rats (Fig. 1D). MTX treatment had the similar effect to that of TS at the dose of 60 mg/kg.
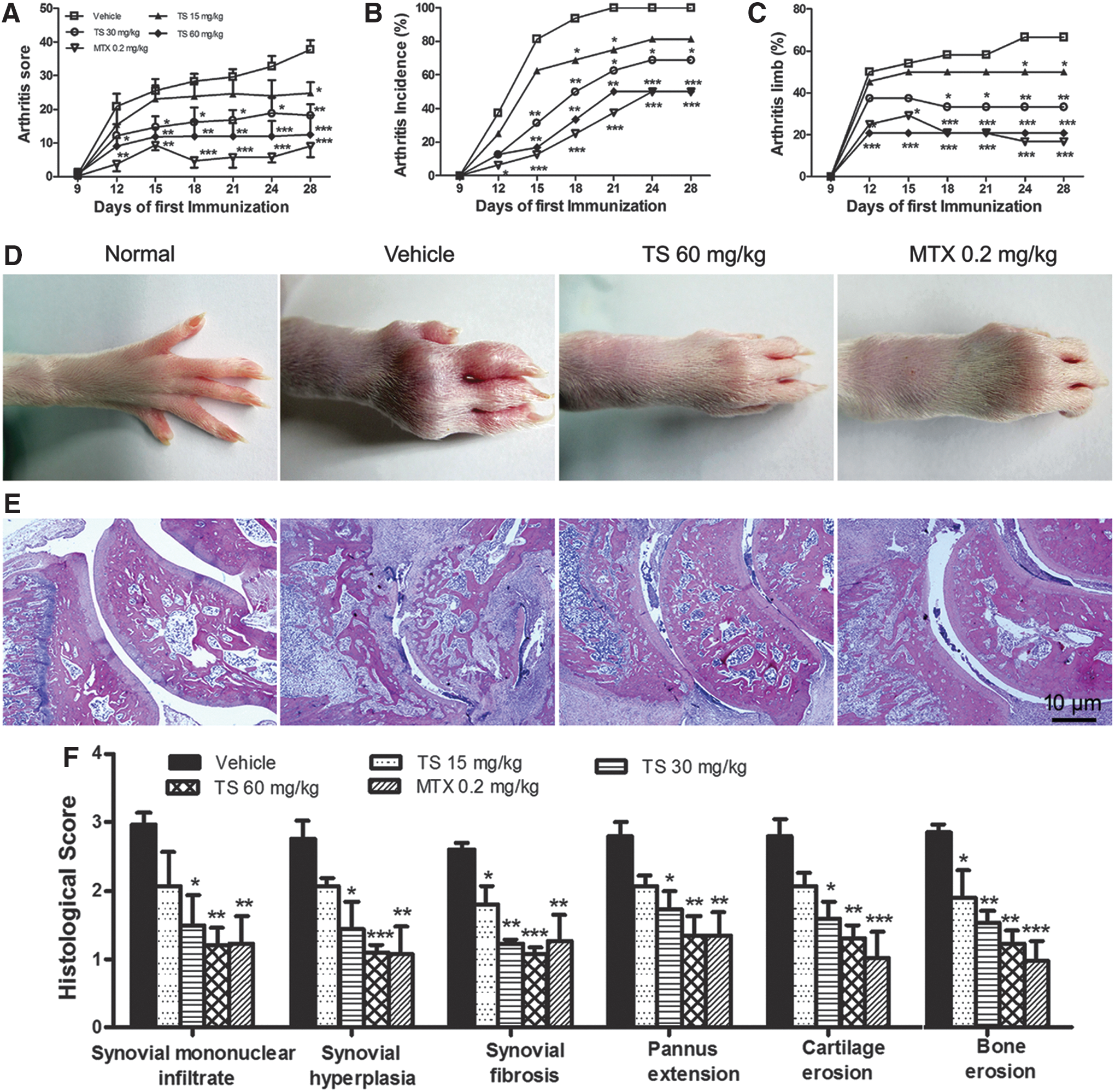
Total saponins (TS) prevent arthritis progression and decrease severity of arthritis in collagen-induced arthritis (CIA) rats. Rats were orally administered TS (15, 30, and 60 mg/kg, respectively), methotrexate (MTX, 0.2 mg/kg), or vehicle for 28 days from the day of first immunization. At the end of the experiment, the arthritis score and arthritis incidence, as well as the percentage of arthritic limbs, were evaluated. TS (15–60 mg/kg) significantly decreased the mean arthritis score (
After the rats were sacrificed, the ankle and knee joints were evaluated histologically and then graded semi-quantitatively for severity of synovial mononuclear infiltrate, synovial hyperplasia, synovial fibrosis, extension of pannus, cartilage, and bone erosion. As shown in Fig. 1E, massive cellular infiltration, hypertrophy of the synovial membrane, pannus formation, and loss of cartilage and bone observed in vehicle-treated rats were repressed by a 60-mg/kg dose of TS. In accordance with the clinically observed effects on the incidence and the severity of arthritis, TS dose-dependently inhibited the histological damage of cartilage and bone, and cumulative arthritis injury scores, as compared with those of the vehicle-treated rats (Fig. 1F). MTX also effectively inhibited these histological severity scores.
TS suppresses peri-articular bone destruction of CIA rats
A micro-CT scan was performed to validate the efficiency of TS in CIA rats. Three-dimensional reconstruction images from the micro-CT scanning data revealed the degree of articular bone erosion on the surface of epiphysis of femur (Fig. 2A) and tibia (Fig. 2B) facing along the articular cavity at the knee joints. Severe bone erosion was observed in vehicle-treated rats with CIA. Mild erosion was observed in the TS- (60 mg/kg) treated and MTX- (0.2 mg/kg) treated groups, and there was no erosion in normal group. Figure 2C showed the two-dimensionally reconstructed bones of knee joints in different groups. Compared with vehicle-treated CIA rats, a dose of 60 mg/kg TS markedly reduced the extent of joint destruction in TS-treated CIA rats. In addition, six parameters, including BV/TV, BS/BV, Tb.N, Tb.Sp, Tb.Th, and BMD, at the distal end of the femoral epiphysis in different groups were detected to quantify the extent of joint destruction. As shown in Fig. 2D, compared with vehicle-treated CIA rats, doses of 30–60 mg/kg TS significantly increased BV/TV, Tb.N, Tb.Th, and BMD, and decreased BS/BV and Tb.Sp, suggesting a protective role of TS on volume and quality of preserved trabecular bone. MTX effectively changed these parameters to the same extent as dose of 60 mg/kg TS.
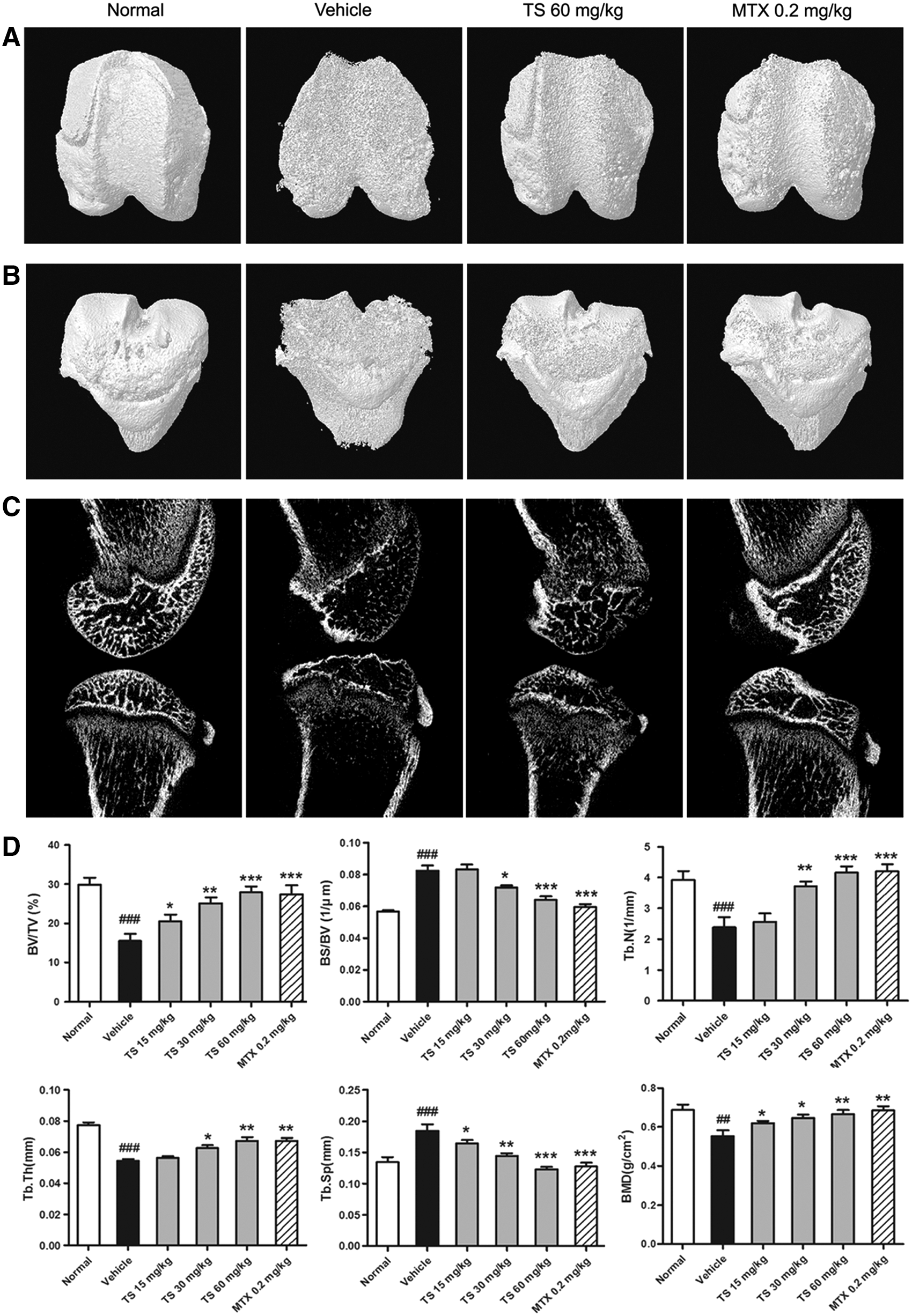
Total saponins (TS) suppress peri-articular bone destruction in collagen-induced arthritis (CIA) rats. Rats were orally administered TS (15, 30, and 60 mg/kg, respectively), methotrexate (MTX, 0.2 mg/kg), or vehicle for 28 days from the day of first immunization. At the end of the experiment, a microfocal computed tomography (micro-CT) scan was performed to validate the efficiency of TS in CIA rats. (
TS inhibits systemic bone loss of CIA rats
As it has been reported that the disease activity of CIA causes marginal (systemic) bone loss as well as peri-articular bone destruction, 36 the secondary spongiosa in the proximal end of the tibial metaphysis was analyzed using micro-CT reconstruction images. Compared to normal rats, the bone area at the secondary spongiosa was significantly reduced in vehicle-treated rats (Fig. 3A). TS (60 mg/kg) inhibited the CIA-induced systemic bone loss. To investigate whether the inhibition of this systemic bone loss by TS was accompanied by structural changes, quantitative histomorphometric analysis of the tibial metaphysis was performed. The bone structure parameters BV/TV, Tb.N, Tb.Th, and BMD were significantly decreased in vehicle-treated rats with CIA compared with normal rats (Table 1). The bone loss reflected by these parameters was significantly blocked in both groups of 30 and 60 mg/kg TS-treated rats. Other structure parameters, BS/BV and Tb.Sp, were significantly increased in vehicle-treated rats compared with normal rats, but these parameters in three groups of TS-treated rats were decreased in a dose-dependent manner compared with those in vehicle-treated rats, with significance in 30 and 60 mg/kg TS-treated rats. MTX treatment significantly attenuated the systemic bone loss and to the similar extent as 60 mg/kg TS treatment.

Total saponins (TS) inhibit extra-articular bone loss in collagen-induced arthritis (CIA) rats. Rats were orally administered TS (15, 30, and 60 mg/kg, respectively), methotrexate (MTX, 0.2 mg/kg), or vehicle for 28 days from the day of first immunization. At the end of the experiment, microfocal computed tomography (micro-CT scan) and enzyme-linked immunosorbent assay were performed to validate the efficiency of TS in CIA rats. (
Rats were orally administered total saponins (TS; 15, 30, and 60 mg/kg, respectively), methotrexate (MTX, 0.2 mg/kg), or vehicle for 28 days from the day of first immunization. At the end of the experiment, a microfocal computed tomography (micro-CT) scan was performed and six parameters including bone volume (BV)/tissue volume (TV) ratio, bone surface (BS)/BV ratio, trabecular thickness (Tb.Th), trabecular number (Tb.N), trabecular separation (Tb.Sp), and bone mineral density (BMD) of distal tibia in different groups were evaluated.
Data are represented as the mean ± standard error of the mean (SEM) (n = 12).
p < 0.01 and ### p < 0.001, comparison with the normal group.
p < 0.05 and ** p < 0.01, and *** p < 0.001, comparison with the vehicle-treated CIA group.
To further verify the effect of TS on systemic bone loss, circulating markers of bone metabolism, such as CTX-I and osteocalcin, were detected in sera. In line with the radiological observation, the level of bone resorption marker CTX-I remained high in vehicle-treated rats, but reduced significantly in TS-treated CIA rats with a dose-dependent manner (Fig. 3B). Conversely, the level of osteocalcin, a bone formation marker, decreased remarkably in vehicle-treated CIA rats compared to normal rats, whereas treatment with TS (60 mg/kg) increased the protein (Fig. 3C).
TS represses osteoclastogenesis in vivo and in vitro
To confirm the effect of TS on the number of osteoclasts, knee joint sections were stained with TRAP. Only TRAP-positive multinucleated cells located at the bone surface within the bone destruction were considered as osteoclasts (Fig. 4A). Compared with vehicle-treated CIA rats, the number of osteoclasts in the areas of bone destruction was significantly decreased in TS-treated CIA rats with a dose-dependent tendency (Fig. 4B). MTX also reduced significantly the number of osteoclasts in the areas of bone destruction compared with vehicle-treated CIA rats.
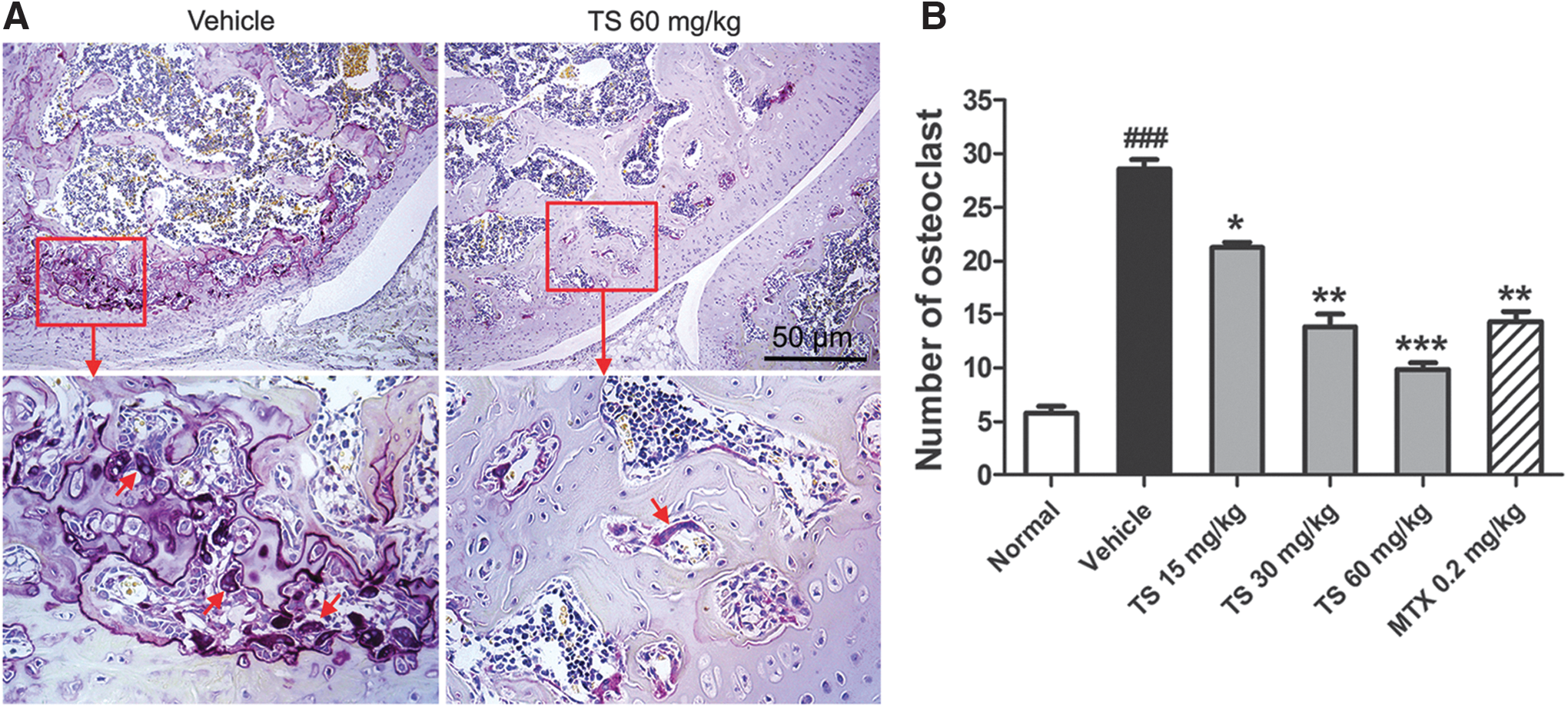
Total saponins (TS) suppresses osteoclast differentiation in collagen-induced arthritis (CIA) rats. Rats were orally administered TS (15, 30, and 60 mg/kg, respectively), methotrexate (MTX, 0.2 mg/kg), or vehicle for 28 days from the day of first immunization. At the end of the experiment, the osteoclast differentiation was evaluated. (
To further validate above effects, we also assessed the effects of TS on osteoclastogenesis in the co-culture system of HFLS and PBMCs. Numerous TRAP-positive multinucleated cells considered as osteoclasts were formed, but few TRAP-positive multinucleated cells were formed when concentrations of 0.1, 0.5, and 2.5 μg/mL TS were added into this co-culture system (Fig. 5A). The number of osteoclasts per area counted under a light microscope was significantly decreased in the co-culture system with the addition of 0.1–2.5 μg/mL TS compared to vehicle-treated group (Fig. 5B).

Total saponins (TS) repress osteoclastogenesis in the co-culture system of human fibroblast-like synovial cells (HFLS) and peripheral blood mononuclear cells (PBMCs). HFLS and PBMCs were co-cultures in α-minimum essential medium (α-MEM) containing 10% fetal calf serum (FCS), 50 ng/mL of macrophage colony-stimulating factor (M-CSF), 10−7 M 1,25- dihydroxyvitamin D3 with or without TS (0.1, 0.5, 2.5 μg/mL, respectively) for 9 days. After that, some dishes were used for tartrate-resistant acid phosphatase (TRAP) staining. (
TS regulates the RANKL/RANK/OPG signaling pathway in vivo and in vitro
To obtain insights into the mechanisms of the inhibitory effects of TS on bone destruction in inflamed joints of CIA rats, the expression of RANK, RANKL, and OPG both at protein and mRNA levels in inflamed joints was detected, respectively, by western blot (Fig. 6A) and quantitative real-time RT-PCR (Fig. 6B), and the serum levels of RANKL and OPG proteins were detected by ELISA assay (Fig. 6C). Compared with vehicle-treated CIA rats, doses of 15–60 mg/kg TS significantly reduced the expression of RANK and RANKL and enhanced the expression of OPG. More interestingly, TS treatments markedly decreased the ratio of RANKL to OPG in the inflamed joints and sera of CIA rats (Fig. 6A–C). MTX also reduced significantly the ratio of RANKL to OPG in the inflamed joints and sera of CIA rats compared with vehicle controls.
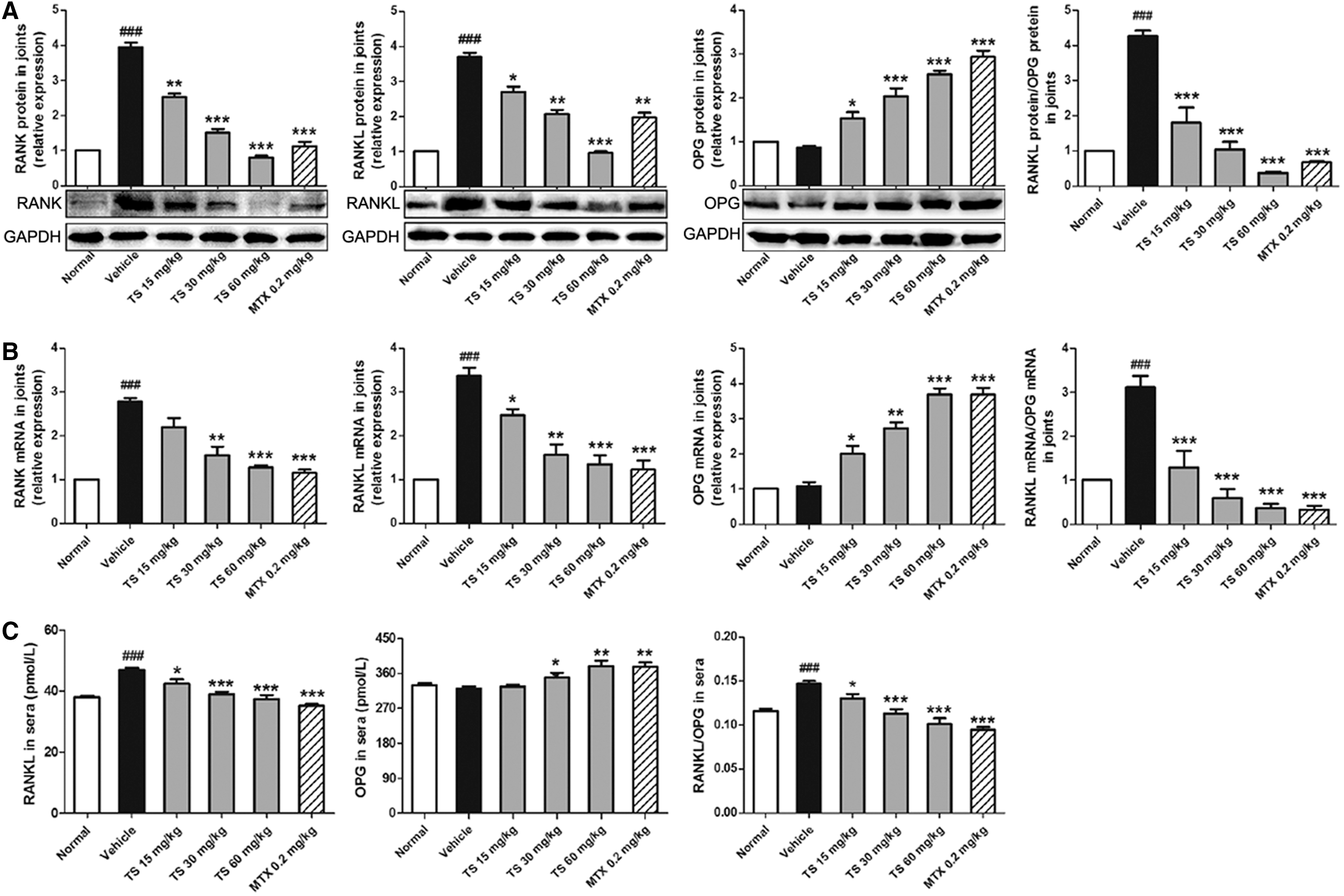
Total saponins (TS) regulate the receptor activator of nuclear factor-κB ligand (RANKL)/RANK/osteoprotegerin (OPG) signaling pathway in collagen-induced arthritis (CIA) rats and in vitro. For in vivo experiments (

We further detected the changes into RANKL and OPG protein expression in HFLS, as well as RANK protein in PBMCs with or without TS treatment for 3, 6, and 9 days, respectively. As shown in Fig. 6D, the supernatant levels of RANK in PBMCs and RANKL in HFLS were increased from day 3 to day 9 during the culture period. TS (0.1, 0.5, and 2.5 μg/mL) treatment was able to reverse the amounts of these proteins dose-dependently, with significance at the 9th day in all groups and at the 6th day only in TS-treated (2.5 μg/mL) group. However, the supernatant levels of OPG in HFLS were decreased from day 3 to day 9 during the culture period, and TS treatment significantly increased the levels of OPG at the 6th and 9th day in a dose-dependent manner. Moreover, TS treatments also markedly decreased the ratio of RANKL to OPG in the supernatant of HFLS at the 6th and 9th day. Similar results were observed for the intracellular levels of these parameters, respectively (Fig. 6E).
TS inhibits the expression of pro-inflammatory cytokines related to bone destruction in sera and joints of CIA rats
Because pro-inflammatory cytokines have been shown to be dominant in the induction of inflammatory bone erosion, we monitored the production of IL-1β, TNFα, IL-6, IL-17, and IL-23 in the sera and joints of CIA rats by ELISA. As shown in Fig. 7, there were relatively low levels of IL-1β, TNFα, IL-6, IL-17, and IL-23 in the sera and joints of normal rats. However, levels of these cytokines were dramatically increased in the sera and joints of CIA rats. All TS-treated rats (15–60 mg/kg) showed effectively reduced serum levels of IL-1β and TNFα, but the suppression of IL-6, IL-17, and IL-23 were only observed in the sera of rats that received 30–60 mg/kg TS (Fig. 7A). Moreover, the levels of IL-1β, IL-6, IL-17, and IL-23 in joints were also reduced by TS (15–60 mg/kg) in a dose-dependent manner (Fig. 7B). A similar result was observed for TNFα in the joints of rats with 30–60 mg/kg TS treatment.
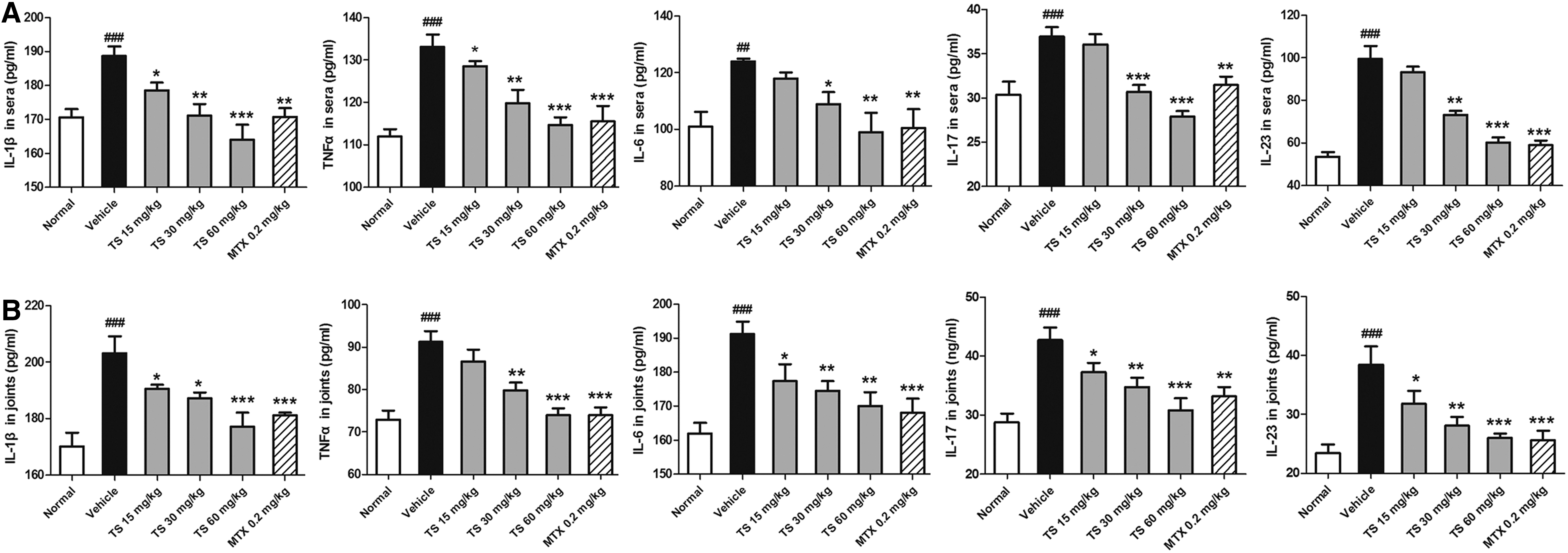
Total saponins (TS) reduce the expression levels of pro-inflammatory cytokines in sera and joints of collagen-induced arthritis (CIA) rats. Rats were orally administered TS (15, 30, and 60 mg/kg, respectively), methotrexate (MTX, 0.2 mg/kg), or vehicle for 28 days from the day of first immunization. At the end of the experiment, the sera and joints were collected from the rats. The levels of interleukin-1β (IL-1β), tumor necrosis factor-α (TNFα), IL-6, IL-17, and IL-23 in sera (
Discussion
The root of A. flaccida has long been used for the treatment of RA in Chinese folk medicine. TS, the main active constituent of this herbal medicine, has been previously demonstrated to exhibit beneficial effects on adjuvant-induced arthritis by its immunomodulatory and anti-inflammatory effects. 20,26 However, the effects of TS on the bone destruction of RA remain unclear. Data presented in this report indicate that TS attenuates RA partially by preventing the focal (peri-articular) bone destruction and systemic (extra-articular) bone loss in the CIA model. Several mechanisms may account for this effect. TS inhibits osteoclast formation both in vivo and in vitro. On the other hand, TS modulates the RANK/RANKL/OPG system, which is biased toward bone destruction. Finally, TS inhibits systemic and local levels of pro-inflammatory cytokines implicated in bone resorption, such as IL-1β, TNFα, IL-6, IL-17, and IL-23.
CIA is a murine autoimmune model useful for investigating the possible therapeutic strategy for joint inflammation, focal bone erosion, and bone atrophy in RA. 37,38 In this study, we analyzed the effect of TS on CIA rats. Our results showed CIA was developed in most rats at about 2 weeks following the primary immunization. Red and/or swelling in the paws, synovial hyperplasia, synovial fibrosis, extension of pannus, cartilage and bone erosion, and joint deformation were observed in vehicle-treated rats. Meanwhile, we noticed that TS attenuated the severity of arthritis by reducing the mean arthritis score as well as arthritis incidence in a dose-dependent manner. In addition, pathological observation indicated that the inflammatory cell infiltration, synovial hyperplasia, pannus, cartilage and bone destruction, and joint deformation of CIA rats were improved significantly by TS treatment. These results suggest that TS effectively prevents arthritis progression and decreases severity of arthritis in CIA rats.
Bone loss in RA occurs both in the joints and throughout the skeleton 39 –41 as a result of a multi-factorial increase in bone resorption. In this study, the results of three-dimensional micro-CT showed that peri-articular bone erosion and extra-articular osteopenia were the common alterations of the bone in the inflamed joints in RA. Six parameters, including BV/TV, BS/BV, Tb.N, Tb.Sp, Tb.Th, and BMD, of inflamed joints in three-dimensional micro-CT were detected to quantify these alterations in all CIA rats of different groups. Our data suggest that oral administration of TS effectively preserves both peri-articular and extra-articular bone density and trabecular thickness of inflamed joints. In addition, TS inhibited the systemic bone resorption, which supported by a dramatic decrease in serum level of CTX-I fragments in CIA rats treated with TS. Because bone resorption and bone formation are coupled to each other, the circulating bone formation marker was also observed. We found that TS significantly increased the reduced serum level of osteocalcin in CIA rats. Taken together, these results indicate that TS can prevent focal (peri-articular) and systemic (extra-articular) bone loss in CIA rats.
Recent studies have identified osteoclasts as the principal cell type responsible for bone destruction in RA. 42,43 Thus, we hypothesized that TS would play a role in osteoclastogenesis in rheumatoid synovium. Consistent with this hypothesis, we found that the administration of TS significantly reduced the number of osteoclasts in the areas of bone destruction with a dose-dependent tendency. Additionally, we also investigated the effects of TS on osteoclast formation in the co-culture system of HFLS and PBMCs because HFLS can efficiently induce the formation of TRAP-positive multinucleated cells when co-cultured with PBMCs. 33,44 According to the results of in vitro study, we confirmed the inhibitive effects of TS on osteoclast formation in this co-culture system, which was consistent with our recent report. 28
Accumulating studies have indicated that osteoclast-mediated bone destruction in RA is regulated by the RANKL/RANK/OPG signaling pathway. 45,46 Both RANK and its ligand RANKL are crucial regulators of osteoclast differentiation. RANK is expressed on the surface of hematopoietic osteoclast progenitors that belong to the monocyte/macrophage lineage and also on mature osteoclasts, as well as on T cells and dendritic cells. In arthritis, osteoclast precursors that express RANK recognize RANKL through cell-to-cell interaction with obsteoblasts/stromal cells, and differentiate into osteoclasts. 47 RANKL expression can be up-regulated by bone resorbing factors, such as vitamin D, IL-1β, IL-6, IL-17, or TNFα in osteoblasts. RANKL is expressed on the cell surface of activated T cells and can be detected in both synovial cells and infiltrating cells by in situ hybridization at the onset of clinical signs of arthritis in animal models. 48
In the current study, we reported the high level of RANK and RANKL expression both in the sera and joints of CIA rats and the increased ratio of RANKL to OPG in the sera and joints of CIA rats, which were reversed by the treatment of TS. The results account for the bone-protective properties of TS in RA. On the other hand, its effects on the expression of OPG further support the postulated bone protective property of TS. OPG is secreted by stromal cells and osteoblasts and competitively inhibits RANKL binding to RANK on the cell surface of osteoclast precursor cells and mature osteoclasts, thus inhibiting the osteoclastogenic actions of RANKL. 49 Excessive production of RANKL and/or a deficiency of OPG could, therefore, contribute to the increased bone resorption typified by the focal bone erosion and bone loss in RA. Our data indicate that TS increased the circulating and local levels of OPG in CIA rats. And the ratio of RANKL to OPG that determines the erosive nature of RA is greatly reduced by TS, accounting for the bone protection achieved by the treatment. Similarly, in vitro results showed that TS could also inhibit RANK and RANKL expression as well as the ratio of RANKL to OPG, and simultaneously increase OPG. In this respect, we speculate that TS regulate the RANKL/RANK/OPG signaling pathway, which has been considered as a potential target for preventing bone destruction in RA patients.
Chronic inflammation is the key mediator for local and systemic bone loss in RA patients. In RA patients, cytokines are abundantly present in the arthritic synovium as well as secreted into the systemic circulation. 50,51 Inflammatory cytokines induce pre-osteoclast fusion, support the survival of mature osteoclasts, and stimulate osteoclastic bone resorption. 42,52 In the present study, TS strongly suppressed both systemic and local levels of pro-osteoclastogenic cytokines, such as IL-1β, TNFα, IL-6, IL-17, and IL-23, partly explaining the application of TS in diseases with inflammatory bone destruction.
In conclusion, our data offer convincing evidence for the first time that TS attenuate RA partially by preventing focal bone destruction and systemic bone loss. This anti-erosive effect resulted in part from the inhibiting osteoclast osteoclastogenesis by regulating the RANKL/RANK/OPG signaling pathway. The suppression of systemic and local pro-osteoclastogenic cytokines implicated in bone resorption by TS was also highly effective. These findings suggest that TS might be an attractive and suitable therapeutic agent for the management of bone destruction in RA.
Footnotes
Acknowledgments
This study was supported by grants from the National Major Scientific and Technological Special Project for “Significant New Drugs Creation” (no. 2011ZX09101-006-03 and 2013ZX09301307), the Beijing Natural Science Foundation (no. 7112096), and the Fundamental Research Funds for the Central Public Welfare Research Institutes (no. ZZ070836).
Author Disclosure Statement
No competing financial interests exist.