
Abstract
Estrogen deficiency is associated with metabolic derangements in menopausal women. Black cohosh has been widely used as an alternative therapy in the treatment of menopausal syndrome. However, its role in metabolism needs to be defined. The aim of the present study was to investigate the long-term effect of black cohosh on glucose and lipid metabolism in a rat model of post-menopause. Adult female Sprague-Dawley rats were sham operated (SHAM), ovariectomized (OVX), OVX with the treatment of estradiol valerate (OVX + E), or OVX with the treatment of isopropanolic black cohosh extract (OVX + iCR). Body weight, body composition, and blood glucose levels of the animals were monitored. The rats were then sacrificed after 3 months of the treatments. At the end of the experiment, OVX + iCR and OVX + E rats exhibited a significant decrease in body weight gain, body and abdominal fat mass, serum triglycerides levels, hepatic fat accumulation, and adipocyte hypertrophy compared with OVX rats. In addition, insulin resistance and glucose intolerance were improved in OVX + iCR but not in OVX + E rats. No hepatotoxicity was detected in OVX + iCR animals. Furthermore, western blot analysis suggested the increased lipolysis in adipose tissue of OVX + iCR and OVX + E rats. Data from in vitro experiments using cultured primary rat adipocytes also showed that black cohosh could affect lipolysis of adipocytes. In conclusion, the long-term treatment of black cohosh at a proper dosage ameliorated metabolic derangements in OVX rats. Thus, this drug is promising for the treatment of metabolic disorders in menopausal and post-menopausal women.
Introduction
M
Hormone replacement therapy is currently the most common and effective treatment for menopausal syndrome. Many studies, however, have shown that the long-term usage of estrogen is associated with the increased risk for breast cancer and endometrial cancer. 5,6 Therefore, non-hormone therapy is necessary, and the ideal treatment should alleviate the symptoms but possess less adverse effects. Black cohosh (Cimicifuga racemosa, CR) is a botanical drug that has been used as an alternative treatment for menopausal symptoms, such as hot flushes, night sweats, vaginal atrophy, and dryness. Its efficiency and safety have been confirmed in many randomized controlled trials. 7 –10 However, the long-term effect of black cohosh on glucose and lipid metabolism remains unknown. Given the clinically wide use of black cohosh, the exact role for black cohosh in metabolism needs to be elucidated. In the present study, we treated female ovariectomized (OVX) rats with black cohosh for 3 months. OVX rats treated with estradiol valerate were used to compare its effect on metabolic parameters with black cohosh.
Materials and Methods
Drugs
Isopropanolic-aqueous C. racemosa (Remifemin tablets, 20 mg crude drug per tablet) was produced by Schaper & Brummer Ltd. & Co. KG, Germany. Estradiol valerate (1 mg per tablet) was produced by Bayer Health Care Co., Ltd., China.
Animals and treatments
Forty female Sprague-Dawley rats aged 8 weeks and weighing 220 ± 10 grams were purchased from the animal facility of the Peking University Health Science Center. The animals were acclimated for 1 week and then randomly divided into four groups (n = 10/group): (1) Sham operated (SHAM), (2) OVX, (3) OVX with estradiol valerate (OVX + E), and (4) OVX with isopropanolic black cohosh extract (OVR + iCR). Rats were anesthetized with isoflurane for surgery. For SHAM rats, the ovaries of rats were exposed but not resected. For the other three groups of rats, the bilateral ovaries were resected. Two weeks after operation, the rats were treated with drugs once daily by oral gavage for 3 months as follows: SHAM and OVX rats, 10 mL of saline/kg; OVX + E rats, 0.8 mg of E/kg; OVR + iCR rats, 60 mg of iCR/kg. All animals were maintained under controlled conditions (21–22°C, 55–65% humidity, 12 hr light:12 hr dark) with free access to rodent soy-free feed and water. All study protocols were approved by the Ethics Committee for Animal Experimentation of the Faculty of Medicine of Peking University Health Science Center.
Oral glucose tolerance test
An oral glucose tolerance test (OGTT) was performed as described 11 at 1 month and 3 months after the treatments, respectively. In brief, the rats were fasted overnight before administration of glucose (3 grams/kg) by oral gavage. Blood glucose levels were measured with the OneTouch Ultraglucometer (Accu-Check Sensor, Roche Diagnostics).
Insulin tolerance test
Rats were subjected to an insulin tolerance test (ITT) at 1.5 and 2.5 months after the treatments, respectively. The rats were fasted overnight before an intraperitoneal injection with insulin (0.75 IU/kg). Blood was collected before insulin administration and then at 5, 10, 15, 20, 25, and 30 min post-injection. Data were given as the values of blood glucose concentrations corrected for fasting glucose.
Body composition analysis
Body composition analysis was performed at the end of the treatments using EchoMRI device (Echo Medical Systems, Houston, TX) to assess total body fat and lean mass of the rats.
Tissue collection and histology
Rats were sacrificed after 3 months of treatments by deep anesthesia with a mixture of ketamine hydrochloride and xylazine. Blood samples were then collected and liver samples were rapidly removed from the animals. One piece of the liver was fixed in 4% paraformaldehyde (PFA), post-fixed in sucrose solution, and then embedded into Tissue-Tek (O.C.T. compound; Sakura), frozen at −80°C overnight, and cut into sections (6-μm thickness) with a Leica Cryostat (LEICA CM1850, Germany) for Hematoxylin & Eosin (H&E) and Oil Red O Staining. The remaining liver tissues were snap frozen in liquid nitrogen and then kept at −80°C. Skeletal muscles from the hind limbs were dissected, snap frozen in liquid nitrogen, and then kept at −80°C. Abdominal fat was dissected and weighed. A piece of the fat was fixed overnight in 4% PFA at 4°C and then embedded in paraffin for H&E staining. The remaining fat tissues were snap frozen in liquid nitrogen and then kept at −80°C. Oil Red O staining for neutral lipid detection and H&E staining were performed according to the standard procedures.
Measurement of serum and hepatic lipids
Serum was collected and serum triglycerides (TGs), total cholesterol, high-density lipoprotein cholesterol (HDL-C) and low-density lipoprotein cholesterol (LDL-C) levels were measured using a Hitachi 7600-100 clinical analyzer (Roche, Japan). The lipids from the livers were extracted as described. 11 Hepatic TGs and cholesterol contents were determined with commercially available kits (BIOSINO Bio-Technology and Science Inc., China).
Measurement of adipocyte size
The adipocyte size was measured as described. 11
Isolation and culture of primary adipocytes
Primary adipocytes were isolated from abdominal fat pads of female Sprague-Dawley rats as described previously. 12 The mature adipocytes were collected and incubated in Phenol Red and serum-free Dulbecco's modified Eagle medium (DMEM) in an atmosphere of 5% CO2 at 37°C for 1 hr before treatments. Cells were then stimulated with black cohosh (0, 0.5, 5, and 50 μg/mL) for 18 hr. The culture medium was collected for glycerol assay, and cells were recovered for RNA extraction.
Glycerol assay
Glycerol content in the culture medium was determined with a commercially available kit (Applygen Technologies, China).
Western blot analysis
Western blotting was performed as described previously. 11 Antibodies to phosphorylated AKT (p-AKT) at Serine473, AKT, phosphorylated hormone-sensitive lipase (p-HSL) at Serine660, and HSL were purchased from Cell Signaling Technology. The antibody to ELF-5 was from Santa Cruz Biotechnology. ELF-5 was measured as an internal control.
Real-time PCR
Total RNA was extracted using TRIzol (Invitrogen, USA) and then reverse transcribed into cDNA. The primer sequences are shown in Table 1. Real-time PCR was performed using SYBR Green according to the manufacturer's instructions. The glyceraldehyde 3-phosphate dehydrogenase gene (GAPDH) was used as an internal standard. The expression of the target genes was normalized to that of GAPDH in the same sample using the 2−ΔCt method.
Statistical analysis
Data are expressed as the mean ± standard error of the mean (SEM). Statistical analyses were performed with GraphPad Prism 5.0 software. Effects of the treatments were analyzed by one-way analysis of variance (ANOVA) followed by Bonferroni's post test. p < 0.05 was considered statistically significant.
Results
Body weight and body composition
Body weight of the rats was similar among the four groups before the treatments. After 3 months, the OVX rats exhibited significantly more weight gain than SHAM rats (p < 0.01) (Fig. 1A, B). In contrast, the weight gain of OVX + E rats was markedly lower than OVX rats (p < 0.01), suggesting that the significant body weight gain in OVX rats was caused by estrogen deficiency. Similarly, OVR + iCR rats also showed marked less weight gain than OVX rats (p < 0.05). No apparent difference was found between OVX + E and OVR + iCR rats.
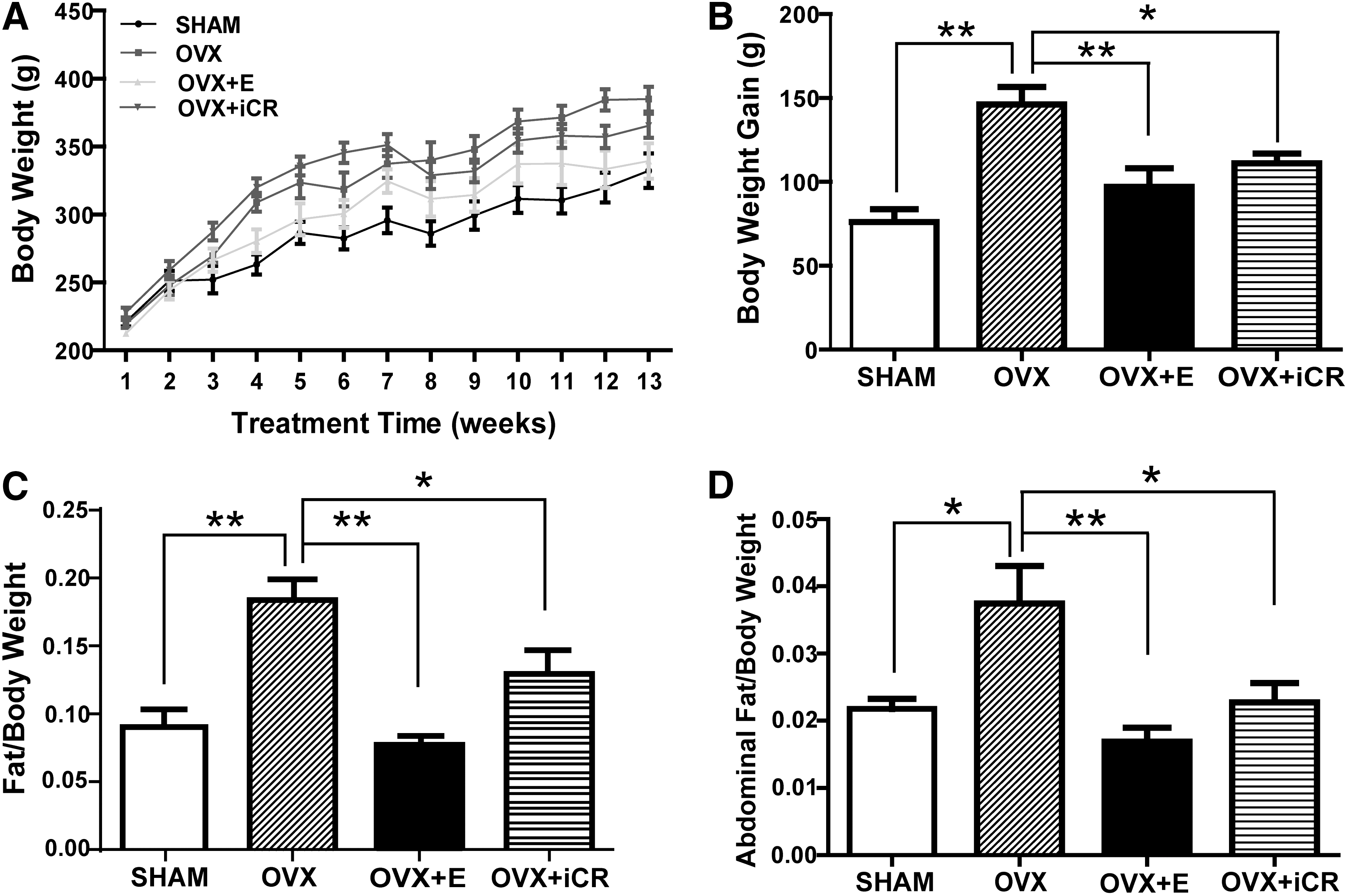
Effects of the treatments on body weight and body composition in the rats. (
To determine the effect of the treatments on body fat and lean mass in vivo, body composition analysis was performed. Similar to body weight gain, the ratio of fat to body weight was significantly higher in OVX rats than in SHAM (p < 0.01), OVX + E (p < 0.01), and OVR + iCR rats (p < 0.05) (Fig. 1C). However, the lean mass/body weight ratio was similar among the four groups of rats (data not shown). These data suggested that estrogen deficiency led to the increased body fat mass in OVX rats and this detrimental effect could be partially reversed by iCR treatment. Abdominal fat mass was collected when the rats were killed. The ratio of abdominal fat/body weight was markedly higher in OVX than in SHAM rats (p < 0.01). (Fig. 1D). Treatment of estradiol or iCR dramatically decreased the abdominal fat/body weight ratio compared with OVX rats (p < 0.01 and p < 0.05, respectively).
Serum lipid profile
After the treatments for 3 months, the OVX rats exhibited significantly higher serum TG levels than SHAM, OVX + E, and OVR + iCR rats (p < 0.01) (Fig. 2A). There was no marked difference in total cholesterol levels between SHAM and OVX rats (Fig. 2B). Treatment with estradiol, however, significantly decreased (p < 0.05), whereas iCR did not affect serum cholesterol levels in OVX rats. There was no difference in serum HDL levels among the four groups of rats (Fig. 2C). No difference was found in serum LDL levels between SHAM and OVX rats (Fig. 2D). Treatment of estradiol or iCR, however, significantly reduced the levels of LDL in OVX rats (p < 0.01 and p < 0.05, respectively) (Fig. 2D).
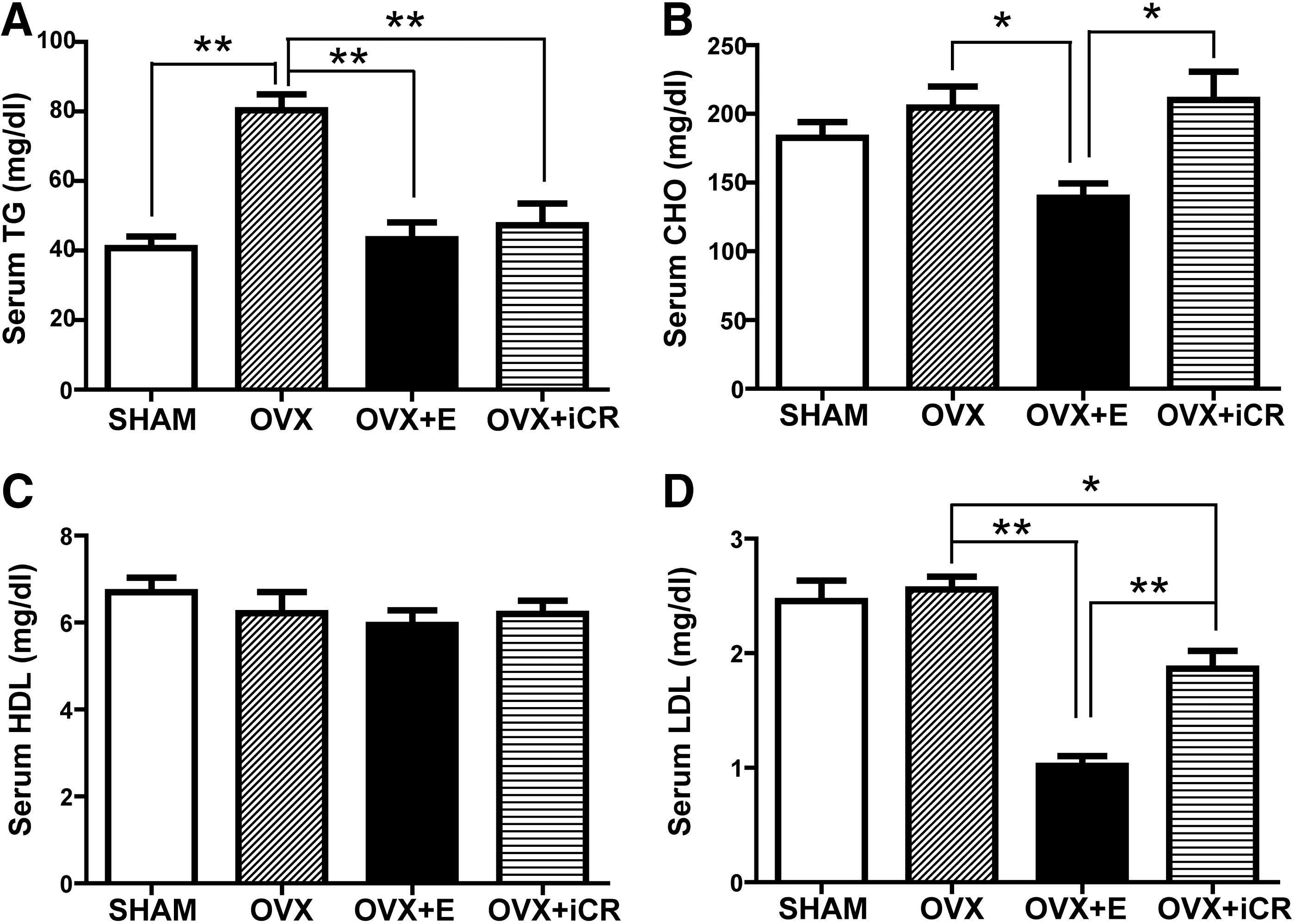
Effects of the treatments on serum lipid profile in the rats. (
OGTT and ITT experiments
To evaluate the effect of the treatments on glucose metabolism, OGTT and ITT were performed. After 1 month of the treatments, OVX rats showed significantly higher serum glucose levels than SHAM rats at 30 min after administration of glucose (Fig. 3A). Statistical analysis revealed a significant elevation in the 0- to 120-min area under curve (AUC) value in OVX rats compared with SHAM rats (p < 0.05) (Fig. 3B), suggesting impaired glucose tolerance in OVX animals. AUC values were lower in OVX + E and OVR + iCR rats than in OVX rats, implying the improved glucose intolerance in OVX animals treated with estradiol or iCR (Fig. 3B). Three months after the treatments, OVX rats still exhibited higher AUC values than SHAM rats, suggesting the presence of impaired glucose tolerance in OVX rats (Fig. 3C, D). No difference was found between OVX and OVX + E rats. OVR + iCR animals displayed decreased AUC values compared with OVX rats. These results suggested that iCR, rather than estradiol valerate, could improve glucose intolerance in OVX rats after 3-month treatment.
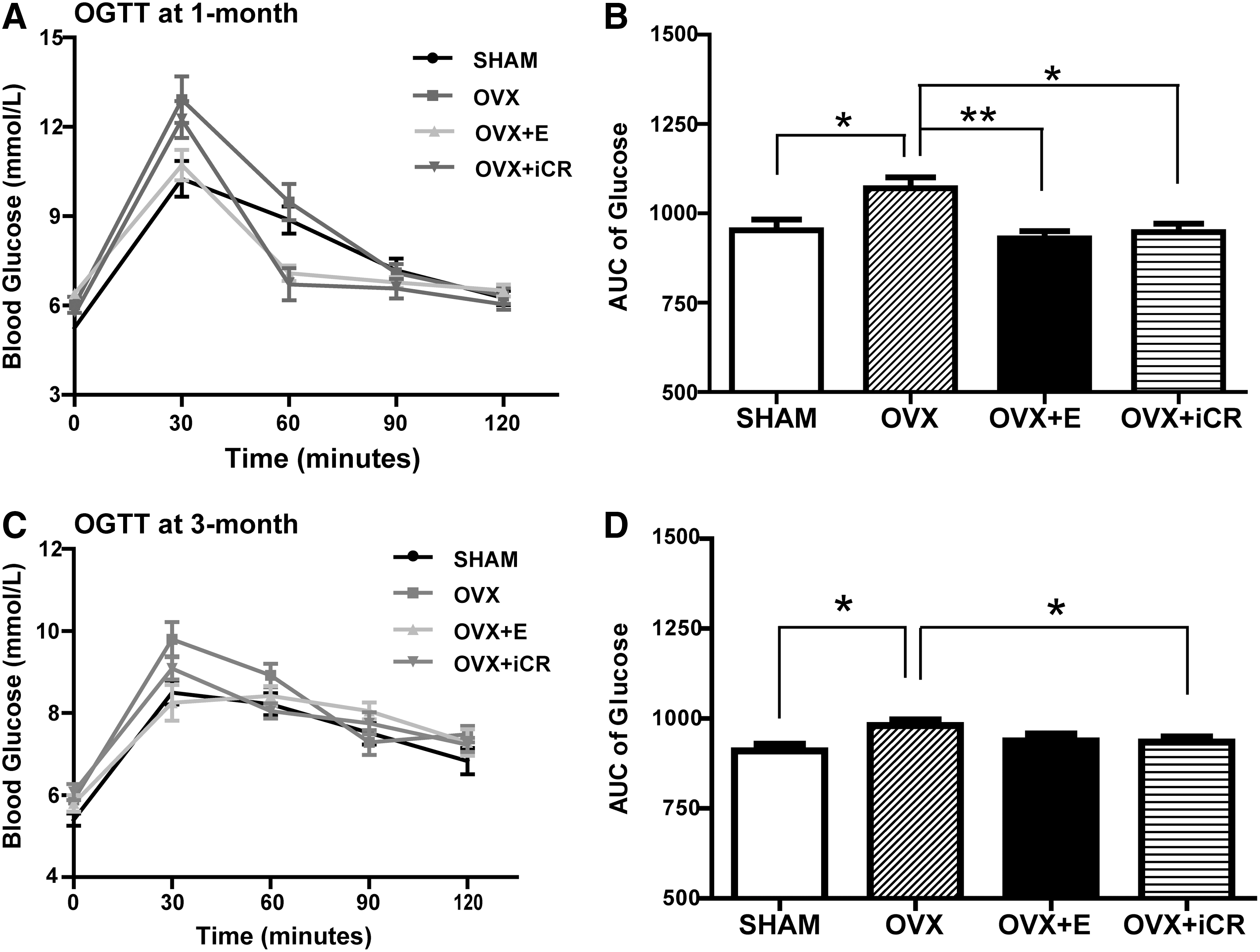
Oral glucose tolerance test (OGTT) experiment. (
In ITT experiments, blood glucose levels were corrected for fasting glucose (Fig. 4A, C). The AUC values were significantly higher in OVX rats than in SHAM at both 1.5- and 2.5-months after the treatment (p < 0.05) (Fig. 4B, D), suggesting insulin resistance in OVX rats. OVR + iCR rats showed reduced AUC values compared with OVX rats at 1.5 and 2.5 months after the treatment (p < 0.05) (Fig. 4B, D), indicating that iCR treatment improved insulin sensitivity in OVX rats. In contrast, estradiol did not influence insulin sensitivity in OVX rats, as evidenced from the blood glucose levels and the AUC values (Fig. 4).

Insulin tolerance test (ITT) experiment. (
Effects of the treatments on lipid metabolism in the liver
Results from H&E staining revealed severe cytoplasmic vacuoles and swelling in the livers of OVX rats, suggesting hepatotoxicity induced by ovariectomization (Fig. 5A). The morphology of the livers was normal in SHAM, OVX + E, and OVR + iCR rats. Masson staining was also performed to detect fibrosis, and no positive staining was found in the liver samples of the four groups of animals (data not shown). We next assessed hepatic lipid contents. Compared with SHAM, OVX + E, and OVR + iCR rats, there was a marked increase in hepatic lipid accumulation in OVX animals, as revealed by Oil Red O staining (Fig. 5A). No obvious difference was found between OVX + E and OVR + iCR rats. The hepatic TGs and cholesterol contents were then determined. Consistent with Oil Red O Staining analysis, the TGs contents were significantly higher in OVX rats than in SHAM (p < 0.01), OVX + E (p < 0.05), and OVR + iCR rats (p < 0.05) (Fig. 5B). These data suggested that treatment of estradiol or iCR ameliorated hepatic steatosis in OVX animals. There was no apparent difference in the hepatic cholesterol contents among the four groups of rats (Fig. 5C).

Effects of treatments on the liver. (
To investigate the underlying mechanisms, we measured the expression of genes governing lipid metabolism. Data from real-time PCR revealed that the expression of fatty acid translocase (CD36), a gene responsible for transporting free fatty acid (FFA) and LDL from blood to the liver, was dramatically reduced in the liver samples of OVX, OVX + E, and OVR + iCR rats compared with SHAM rats (p < 0.01). These results suggested that ovariectomization led to a decrease in fat transport and treatment of estradiol or iCR could not reverse it (Fig. 5D). Fatty acid synthase (FAS) is a critical gene regulating the synthesis of fatty acids (FAs) in the liver. The expression of FAS was significantly higher in OVX than in SHAM rats (p < 0.05), suggesting that ovariectomization induced hepatic FA synthesis in the animals. No obvious difference was found among OVX + E, OVR + iCR, and OVX rats, indicating that treatment of estradiol or iCR did not affect hepatic FA synthesis.
Expression of ELOVL family member 6 (ELOVL6), an enzyme critical for the development of obesity-induced insulin resistance, was markedly higher in OVX than in SHAM rats (p < 0.01). No statistical difference was observed between OVX and OVX + E animals. In contrast, ELOVL6 expression was significantly decreased in OVR + iCR rats compared with OVX rats (p < 0.05). These data suggested that ovariectomization up-regulated ELOVL6 expression, which could be rescued by iCR but not estradiol. We also measured the expression of genes important for the oxidation of fatty acids in the liver. The mRNA level of peroxisome proliferator activated receptor α (PPARα) was significantly lower in OVX than in SHAM rats (p < 0.05). Treatment of estradiol or iCR induced an elevation in PPARα expression in OVX rats (p < 0.05), suggesting that estradiol or iCR treatment increased the fatty acid oxidation, thus contributing to the attenuated hepatic TG accumulation in the OVX + E and OVR + iCR rats.
Microsomal triglyceride transfer protein (MTTP) is essential for the assembly and secretion of very low density lipoprotein (VLDL). In OVX rats, the expression of MTTP was significantly lower than that in SHAM rats (p < 0.01). Treatment of estradiol or iCR elevated MTTP expression in OVX rats (p < 0.05). These results suggested that ovariectomization impaired transporting TGs out of the liver, which could be ameliorated by estradiol or iCR treatment.
We also measured the expression of genes important for cholesterol metabolism, including liver X receptor α (LXRα), LXRβ, and sterol regulatory element-binding protein-2 (SREBP-2). No apparent difference was observed among the four groups of animals (data not shown), which was consistent with data on hepatic cholesterol content measurement. The expression of the genes critical for gluconeogenesis was detected as well, including phosphoenolpyruvate carboxykinase (PEPCK) and glucose-6–phosphatase (G6Pase). There was no difference in the gene expression among the four groups of rats (data not shown).
In addition, the expression of p-AKT and AKT was measured in the liver. Western blot analysis revealed that the ratio of p-AKT/AKT was significantly lower in OVX than in SHAM rats (p < 0.01) and significantly higher in OVR + iCR than in OVX rats (p < 0.05) (Fig. 5E). No apparent difference was observed between OVX + E and OVX rats.
Effects of the treatments on abdominal adipose tissue
To evaluate the adipocyte function, we first performed H&E staining of the adipose tissue. The enlarged adipocytes were present in OVX rats, but not in the other animals (Fig. 6A). The adipocyte size was measured and then the distribution curve was obtained. The number of enlarged adipocytes was increased in OVX rats compared with SHAM, OVX + E, and OVR + iCR rats, as reflected by the right shift of the distribution curve (Fig. 6B). Correspondingly, the mean adipocyte diameter was significantly greater in OVX rats than in SHAM (p < 0.01), OVX + E (p < 0.01), and OVR + iCR rats (p < 0.01) (Fig. 6C), suggesting that ovariectomization induced hypertrophic adipocytes, which was rescued by estradiol or iCR.

Effects of the treatments on the rat abdominal adipose tissue and effect of black cohosh on lipolysis and lipogenesis in rat adipocytes. (
Moreover, the expression of p-AKT, AKT, p-HSL, and HSL was measured in the adipose tissue. The ratio of p-AKT/AKT was significantly lower in OVX than in SHAM rats (p < 0.05) (Fig. 6D). Treatment of OVX rats with iCR, but not estradiol, markedly increased the p-AKT/AKT ratio compared with OVX rats (p < 0.05). The ratio of p-HSL/HSL did not differ between SHAM and OVX rats, but was significantly higher in OVX + E and OVX + iCR than in OVX rats (Fig. 6E).
Effects of black cohosh on lipolysis and lipogenesis in rat adipocytes
To investigate the possible mechanisms underlying the effects of black cohosh on adipose tissue, mature rat adipocytes were cultured and treated with different concentrations of iCR for 18 hr. Glycerol content serves as an index of lipolysis of adipocytes, and therefore glycerol concentrations were measured in the culture media. As illustrated in Fig. 6F, iCR up-regulated lipolysis in a dose-dependent manner, but the high and low doses of iCR affected lipolysis differently. At low concentrations (0.5 and 5 μg/mL), iCR significantly decreased glycerol contents compared with controls (p < 0.01), suggesting reduced lipolysis by low doses of iCR. In contrast, 50 μg/mL iCR induced a marked increase in glycerol. In addition, we also treated the cells with estradiol (10−7 M) for 18 hr and found that estradiol did not affect lipolysis of adipocytes (data not shown).
To evaluate whether iCR influences lipogenesis in adipocytes, we measured the expression of genes critical for lipogenesis, including PPARγ, SREBP-1, and FAS. Results from real-time PCR analysis revealed that 50 μg/mL iCR significantly increased PPARγ expression (Fig. 6F). The levels of SREBP-1 and FAS did not change in response to iCR (data not shown). These data suggested that 50 μg/mL iCR promoted, whereas iCR at low concentrations (0.5 and 5 μg/mL) did not affect, lipogenesis in rat adipocytes.
Discussion
Estrogen plays an important role in the regulation of metabolic homeostasis. In menopausal and post-menopausal women, estrogen deficiency-caused obesity and obesity-related chronic diseases are important public health issues. As a plausible alternative treatment, black cohosh can effectively relieve menopausal symptoms. Nevertheless, current data on the effect of black cohosh on metabolism are conflicting. Rachon and colleagues have reported that black cohosh attenuated body weight gain and intra-abdominal fat accumulation in OVX rats after 6 weeks of treatment. 13 Studies by Seidlova-Wuttke et al. have showed the smaller paratibial fat depot size in black cohosh-treated OVX rats for 12 weeks. 14 In humans, however, the short-term use of black cohosh did not seem to have any obvious effects. 15 This striking disparity is likely due to the different treatment period, the dosage of the drug used, and so forth. Therefore, we treated the OVX rats with black cohosh for 3 months, aiming at elucidating the long-term effect of black cohosh on metabolic parameters.
In agreement with other studies, 16,17 OVX rats showed marked body weight gain, significantly increased body fat mass, and dyslipidemia. Dyslipidemia is common in post-menopausal women, such as reduced HDL and elevated total cholesterol, TGs, and LDL. 18 In the present study, black cohosh markedly reduced body weight gain and body fat mass in OVX rats. In addition, it decreased serum TGs and LDL, but not cholesterol levels. Rachon et al. have shown a decrease in TGs, an increase in LDL, and unchanged cholesterol levels in the rats after black cohosh treatment. 13 The discrepancy was likely due to the different treatment period and the drug dosage.
Many post-menopausal women exhibit decreased insulin sensitivity and increased glucose intolerance. 1,2,4,7 Consistent with previous studies, 19 OVX rats showed glucose intolerance and insulin resistance in the present work. Estradiol valerate treatment seemed to improve glucose tolerance only after a short period of the treatment, and that effect was lost with time. Moreover, estradiol valerate did not affect insulin sensitivity. In contrast, treatment of black cohosh improved both glucose tolerance and insulin sensitivity in OVX rats. AKT is a crucial node of the insulin signaling pathway regulating glucose metabolism. An increased p-AKT/AKT ratio was detected in the liver and adipose tissue, but not in the skeletal muscle (data not shown) in OVR + iCR rats compared with OVX rats. These data suggest a possible role for AKT in the improved glucose metabolism in the liver and adipose tissue as insulin target organs of OVR + iCR rats. Indeed, the black cohosh extract Ze 450 has been shown to improve glucose metabolism and insulin sensitivity in diabetic ob/ob mice. 19 The beneficial effect of black cohosh on glucose metabolism gives it an advantage over estradiol valerate.
Accumulation of fat mass in obesity may result from hypertrophy and/or hyperplasia. When obesity occurs, hypertrophy of adipocytes first appears to meet the need for excessive fat accumulation. 20 Lafontan has shown that enlarged adipocytes are strongly associated with insulin resistance. 21 In agreement with previous studies, 22,23 we have found significantly increased abdominal fat mass, enlarged adipocytes, and reduced insulin sensitivity in OVX rats. Treatment of black cohosh decreased abdominal fat mass and adipocyte size, which may contribute to the up-regulated insulin sensitivity. HSL is a critical enzyme in lipolysis. The increased HSL phosphorylation in adipose tissue suggests the increased lipolysis in OVX + E and OVX + iCR animals. In addition, results from in vitro experiments also supported that black cohosh affects lipolysis of adipocytes. The effect of black cohosh on lipolysis may thus contribute to the decrease in body weight gain, body fat mass, and adipocyte size. The effect of estradiol on lipolysis is complicated. In humans, it has been shown that estrogen attenuates the lipolytic response only in subcutaneous and not in visceral fat depots. 24 In rodents, estrogen does not seem to affect lipolysis in vitro 25 or in vivo. 26 In the present study, estradiol valerate treatment affects lipolysis in vivo but not in vitro. The effects of black cohosh and estradiol on lipolysis may help to better understand their roles in regulating weight gain, fat mass, adipocyte size, and insulin sensitivity.
Liver is an important organ in glucose and lipid metabolism. 22 Abnormal hepatic gluconeogenesis and lipid accumulation are closely associated with insulin resistance. In OVX + iCR rats, the expression of genes critical for gluconeogenesis did not change. The reduced hepatic lipid content may thus contribute to the improved systemic insulin sensitivity of OVX + iCR animals. Abnormal fat accumulation in the liver is a characteristic of NAFLD. 27 Emerging evidence has shown that the risk of NAFLD is increased by two-fold in menopausal women, 28,29 and the lack of estrogen might accelerate the development and severity of NAFLD in post-menopausal women. 30 Consistent with other studies, 17,31,32 there was apparent liver steatosis in OVX rats, which was rescued by the treatment of estradiol or black cohosh. The hepatic fat content is determined by a dynamic balance among fat uptake, synthesis, oxidation, and exportation. 33 The analysis of gene expression revealed that black cohosh reduced hepatic fatty acid synthesis, increased fat oxidation, and up-regulated exportation of TG, thus leading to the decreased fat accumulation in the liver of OVR + iCR rats. The effect of estradiol on reducing hepatic fat contents, however, seemed to be caused by the increased oxidation and exportation of TGs.
Acute liver damage following consumption of black cohosh was reported in certain clinical cases. 34 –36 Lude et al. have also found the impairment of the mitochondrial β-oxidation, membrane potential, and oxidative phosphorylation of HepG2 cells by high concentrations of black cohosh. 37 Nevertheless, Nasr et al. did not observe the liver disease after the use of black cohosh for 1 year in healthy post-menopausal women. 38 In the current study, no liver toxicity was observed in OVR + iCR rats after the treatments for 3 months. Mazzanti and colleagues 39 have suggested the use of 300 mg/kg per day of black cohosh as a safe dosage in rats. In our study, a dose of 60 mg/kg per day of black cohosh efficiently ameliorated liver steatosis and glucose metabolism. Thus, the dosage of black cohosh administered and the physical status of the patients might affect the influence of this drug on the liver, especially the possibility of inducing liver toxicity.
In summary, our results showed that treatment of black cohosh ameliorated metabolic disorders in OVX rats, including reducing weigh gain, serum TGs, and liver steatosis without inducing hepotoxicity. In particular, black cohosh has a better effect on improving glucose metabolism than estradiol valerate. Moreover, we show for the first time that black cohosh influences lipolysis, which is likely to contribute to reduced body weight gain, fat mass, adipocyte hypertrophy, and increased insulin sensitivity. Our data suggest that black cohosh at a proper dose is promising for the treatment of metabolic derangements in women around or after menopause.
Footnotes
Acknowledgments
This work was supported by the National Basic Research Program of China (973 Program) (2013CB945202) and the National Natural Science Foundation of China (81270666, 81471077, and 81270943).
Author Disclosure Statement
No competing financial interests exist.