
Abstract
Based on pivotal roles of oxidative stress, dopaminergic and cholinergic systems on the pathophysiology of Parkinson's disease (PD), the searching for functional food for patients attacked with PD from Cyperus rotundus and Zingiber officinale, the substances possessing antioxidant activity, and the suppression effects on monoamine oxidase B (MAO-B) and acetylcholinesterase (AChE) have been considered. In this study, we aimed to determine the effect of the combined extract of C. rotundus and Z. officinale (CP1) to improve motor and memory deficits, neurodegeneration, oxidative stress, and functions of both cholinergic and dopaminergic systems in the animal model of PD induced by 6-hydroxydopamine hydrochloride (6-OHDA). Male Wistar rats, weighing 180–220 g, were induced unilateral lesion at right substantia nigra by 6-OHDA and were orally given CP1 at doses of 100, 200, and 300 mg/kg body weight for 14 days after 6-OHDA injection. The results showed that the 6-OHDA rats treated with CP1 increased spatial memory, but decreased neurodegeneration, malondialdehyde level, and AChE activity in hippocampus. The decreased motor disorder and neurodegeneration in substantia nigra together with the enhanced catalase activity, but decreased MAO-B activity in striatum, were also observed. The memory enhancing effect of CP1 might occur through the improved oxidative stress and the enhanced cholinergic function, whereas the effect to improve motor disorder of CP1 might occur through the enhanced dopaminergic function in striatum by decreasing the degeneration of dopaminergic neurons and the suppression of MAO-B. Therefore, CP1 is the potential functional food against PD. However, further researches in clinical trial and drug interactions are essential.
Introduction
P
Abundant medicinal plants have been used for a long term in traditional folklore with safety and reputed for longevity promotion, neuroprotection, and as a memory enhancer. Most of the medicinal recipes in traditional folklore have been used as polyherbal recipes more than a single plant, based on the belief that the polyherbal recipes can provide more benefit due to the interaction effect of ingredients and the multitarget approach. 8 However, less scientific evidence concerning the beneficial effects of the either single plant or polyherbal recipe for treating PD was available. 9
Numerous lines of evidence demonstrated that Cyperus rotundus, a plant in a family of Cyperaceae, and Zingiber officinale, a plant in a family of Zingiberaceae, possess a neuroprotective effect, antioxidant activity, and acetylcholinesterase (AChE) and monoamine oxidase B (MAO-B) inhibitory. 10 –13 Based on these reputations and the critical roles of oxidative stress, cholinergic and dopaminergic systems on the pathophysiology of both motor and nonmotor symptoms of PD, 14 the beneficial effects of both plants to improve neurodegeneration in PD are focused.
To optimize the benefit of the plant extracts, the positive modulation effect on the interaction of both plants has gained attention. It was hypothesized that the combination extract of C. rotundus and Z. officinale (CP1) could enhance motor and memory deficits in PD. To test this hypothesis, this study aimed to determine the antioxidant effect, AChE and MAO-B inhibitory of CP1. In addition, the in vivo study was also carried out to determine the neuroprotective effect and the beneficial effect on both motor and nonmotor symptoms in the animal model of PD induced by 6-hydroxydopamine hydrochloride (6-OHDA).
Materials and Methods
Plant collection and extract preparation
Aerial part of C. rotundus and rhizome of Z. officinale were harvested from Khon Kaen province, Thailand, during September–November 2012. They were authenticated by the Associate Professor Panee Sirisa-Ard, from Faculty of Pharmacy, Chiang Mai University, Thailand. The plant materials were prepared as 95% ethanolic extract. A ratio of ethanolic extract of aerial part of C. rotundus and rhizome of Z. officinale, which provided the highest potential to enhance memory (1:5), was prepared. The percent yield of the C. rotundus and Z. officinale extracts were 7.41% and 10.48%, respectively. The combined extract was kept at −20°C in a dark bottle until used.
High-performance liquid chromatography analysis
The analysis of sample was carried out through gradient high-performance liquid chromatography (HPLC) system consisting of 515 HPLC pump and 2998 Photodiode array detector of Waters Company. Chromatographic separation was performed using Purospher®STAR, C-18 encapped (5 μm), LiChroCART®250-4.6, and HPLC-Cartridge, Sorbet Lot No. HX255346 (Merk). Two mobile phases consisting of methanol and 2.5% acetic acid in deionized water were used to induce gradient elution. The injection volume was 20 μL, and the flow rate was 1.0 mL/min. Data analysis was performed using Empower™ 3.
Determination of antioxidant activity
Radical scavenging activity against 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical of the combined extract of Z. officinale and C. rotundus (CP1) was determined spectrophotometrically.
15
The principle of the assay is based on the color change of the DPPH solution from purple to yellow when the radical is quenched by the antioxidant. In brief, 2.96 mL of a 0.1 mM solution of DPPH in methanol was incubated with 40 μL of various concentrations of extract (1.0, 2.0, 5.0, 10.0, 20.0, 25.0 mg/mL) at room temperature for 30 minutes. The decrease in DPPH radicals was evaluated by the measurement of optical density at 515 nm. The stable-free radical scavenging capacity was presented as the percentage of inhibition of DPPH radicals, calculated according to the following equation:
Determination of antioxidant activity by ferric reducing antioxidant power
Ferric reducing antioxidant power (FRAP) assay was evaluated according to the procedure previously described 16 with some modifications. Briefly, the working FRAP reagent was mixed with 25 mL of 300 mM acetate buffer (3.1 g C2H3NaO2·3H2O and 16 mL C2H4O2), pH 3.6, 2.5 mL of 10 mM tripyridyltriazine solution in 40 mM HCl, and 2.5 mL of 20 mM FeCl3·6H2O solution. Then, 1.8 mL of the FRAP solution was mixed with the plant extract (10 μL) in 1 mL distilled water. The absorbance of reaction mixture at 593 nm was measured spectrophotometrically after incubation at 37°C for 10 minutes. The results were expressed as micromolar ascorbic acid per 100 g fresh weight.
Determination of AChE inhibition
AChE inhibitory activity was measured by using Ellman's colorimetric method.
17
Briefly, in the 96-well plates, 25 μL of 15 mM ATCI, 75 μL of 3 mM DTNB, and 50 μL of 50 mM Tris–HCl, pH 8.0, containing 0.1% bovine serum albumin, and 25 μL of the tested phytochemicals were added. The absorbance was measured at 405 nm after a 5-minute incubation at room temperature. Then 25 μL of 0.22 U/mL of AChE was added, incubated for 5 minutes at room temperature, and the absorbance was measured at 412 nm. AChE (5–1000 μM) was used as a reference standard. The percentage inhibition was calculated using the following equation:
where Asample is the absorbance of the sample extracts, and Acontrol is the absorbance of the blank (50% aqueous methanol in buffer).
Besides on the in vitro assay of AChE mentioned earlier, we also determined AChE activity in hippocampal and striatal homogenate. In brief, hippocampus and striatum were isolated and homogenized in ice cold 0.1 M phosphate-buffered saline (pH 8.0). The homogenate was centrifuged at 1000 g for 10 minutes at 4°C, and the supernatant was used as the source of enzyme in AChE assay. AChE activity in hippocampus was evaluated based on Ellman's method with slight modifications. 18
Determination of MAO-B inhibition
The MAO-B activity was assessed using the microplate reader method. 19 Rats were decapitated, and brains were rapidly removed and frozen on dried ice. Then, they were homogenized in a sodium phosphate buffer (0.1 M, pH 7.4) and centrifuged at 15,000 g for 20 minutes. Supernatant was removed and used for the estimation of MAO-B activity. The brain supernatant 150 μL was added into the solution containing 2.5 mL Tris buffer (0.1 M, pH 7.4) and 100 μL of 0.1 M benzylamine. Then, the absorbance was measured at 340 nm for 30 minutes against the blank containing Tris buffer and 5-hydroxytryptamine. In addition to the in vitro assay of MAO-B, we also determined MAO-B activity in the hippocampal and striatal homogenate. The rats were divided into various groups as described in the experimental protocol. At the end of experiment, all rats were sacrificed. The hippocampus and striatum were isolated and prepared as a homogenate to determine the activities of MAO-B. The activities of MAO-B were determined by using the colorimetric method. 20
Animals
Young adult male Wistar rats, 8 weeks old, were used as experimental animals. They were obtained from National Laboratory Animal Center, Salaya, Nakorn Pathom. The weights of the animals on the first day of experiment are 180–220 g. They were housed six per cage and maintained in a 12-h light:12-h dark cycle and given access to food and water ad libitum. The experiments were performed to minimize animal suffering, and the experimental protocols were approved by the Institutional Animal Care and Use Committee Khon Kaen University, Thailand (AEKKU 41/2554).
Drugs and chemicals
6-OHDA was purchased from Sigma-Aldrich Co. Sodium pentobarbital was obtained from Jagsonpal Pharmaceuticals Ltd. All other chemical substances were analytical grade and purchased from Sigma Chemical Company.
Experimental protocol
All rats were randomly assigned to 7 groups of 12 animals each as following:
Group I Vehicle+saline; rats were orally given propylene glycol, which served as the vehicle to suspend the combination extract of CP1 once daily for 14 days after the injection of saline into the right substantia nigra.
Group II Vehicle+6-OHDA; rats were orally treated with propylene glycol once daily at a period of 14 days after the injection of 6-OHDA into the right substantia nigra.
Group III Vitamin C + 6-OHDA; the animals were orally treated with vitamin C (200 mg/kg) once daily for a period of 14 days after the injection of 6-OHDA into the right substantia nigra.
Group IV L-DOPA +6-OHDA; the animals were orally treated with L-DOPA (5 mg/kg) once daily for a period of 14 days after the injection of 6-OHDA into the right substantia nigra.
Group V–VII CP1 + 6-OHDA; rats were treated with CP1 at doses of 100, 200, and 300 mg/kg for a period of 14 days after the injection of 6-OHDA into the right substantia nigra; rats in all groups were orally given the assigned substances at a period of 14 days after the injection of 6-OHDA into the right substantia nigra. The memory and motor assessment was performed every 7 days throughout a 14-day study period, whereas the determinations of malondialdehyde (MDA) level and the activities of superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GSH-Px), AChE, and MAO-B in the hippocampus and striatum were performed at the end of the study. Moreover, the density of survival neurons in various subregions of hippocampus, including CA1, CA2, CA3, and dentate gyrus, was also determined. Immunohistochemical staining to investigate the density of dopaminergic neurons in substantia nigra was performed using the tyrosine hydroxylase.
Substantia nigra lesion
The animals were anesthetized by intraperitoneal injection of sodium pentobarbital (Jagsonpal Pharmaceuticals Ltd.) at a dose of 60 mg/kg body weight (BW). Each animal was mounted on a stereotaxic stand, the skin overlying the skull was cut to expose the skull, and the coordinates for the substantia nigra pars compacta were accurately measured (anteroposterior −0.5 mm from bregma, mediolateral 2.1 mm from midline, and dorsoventral −7.7 mm from the skull). A total of 6 μg of 6-OHDA was dissolved in 2 μL 0.2% ascorbic acid saline 21 and was perfused into the right substantia nigra through a 30-gauge stainless needle. After the surgery, all animals were allowed to recover from anesthesia and then placed in their cages.
Determination of spatial memory
Spatial memory was evaluated through the Morris water maze test. The water maze consists of a metal pool (170 cm in diameter ×58 cm tall) filled with tap water (25°C, 40 cm deep). The pool was divided into four quadrants (Northeast, Southeast, Southwest, and Northwest). The water surface was covered with nontoxic milk. The removable platform was placed below the water level at the center of one quadrant. For each animal, the location of the invisible platform was placed at the center of one quadrant and remained there throughout training. The times that animals spent to climb on the hidden platform were recorded as escape latency. To determine the capability of the animals to retrieve and retain information, the platform was removed 24 hours later and the rats were released into the quadrant diagonally opposite to that which contained the platform. Time spent in the region that previously contained the platform was recorded as retention time. 22
Determination of motor function
When all rats regained consciousness after the last laser acupuncture treatment, the changes in apomorphine-induced rotation behavior (0.5 mg/kg, s.c.) were assessed using a cylindrical container. The diameter of the cylindrical container used for measuring rotational activity was 40 cm. The animals were habituated in the cylindrical container for 10 minutes, and the rotations made by the animals to either side were counted for 45 minutes. The net number of rotations was counted as follows: the number of contralateral rotations—the number of ipsilateral rotations with respect to the 6-OHDA injection side. This behavioral test was performed blindly. 23,24
Histological procedure
After the anesthesia with sodium pentobarbital (60 mg/kg BW), brains were subjected to transcardial perfusion with fixative solution containing 4% paraformaldehyde in 0.1 M phosphate buffer pH 7.3. After the perfusion, brains were removed and stored overnight in a fixative solution that was used for perfusion. Then, they were infiltrated with 30% sucrose solution at 4°C. The specimens were frozen rapidly, and 10-μM-thick sections were cut on cryostat. The selected sections were rinsed in the phosphate buffer and picked up on slides coated with 0.01% of aqueous solution of a high molecular weight poly
Immunohistochemical evaluation
Immunohistochemical staining to investigate dopaminergic neuron was performed using the monoclonal anti-tyrosine hydroxylase antibody produced in mouse (Sigma Chemical Company) and a modification of a previously described protocol using the DAKO Strept ABC Complex/HRP duet kit. In brief, the sections were eliminated endogenous peroxidase activity by 0.5% H2O2 in methanol. Sections were washed in running tap water and distilled water for 1 minute each, then rinsed in KPBS and KPBS-BT for 5 minutes per each process. Excess buffer was removed, then incubated for 30 minutes in a blocking solution composed of 5% normal goat serum in KPBS-BT. The sections were then incubated in mouse monoclonal anti-tyrosine hydroxylase antibody diluted 1:400 in KPBS-BT at room temperature for 2 hours and then incubated at 4°C for 48 hours. The tissue was rinsed in KPBS-BT (two washes × 7 minutes), incubated for 1 hour in biotinylated goat anti-mouse IgG antibody, rinsed in KPBS-BT (two washes × 7 minutes), and then incubated in Strep ABC Complex/HRP for 4 hours. According to the preparation for visualization step, sections were rinsed in KPBS-BT (1 minute) and KPBS (two washes × 10 minutes). Tyrosine hydroxylase immunoreactivity was visualized using 0.025% 3,3′-diaminobenzidine (DAB; Sigma-Aldrich Co.) and 0.01% H2O2 for 48 hours. Finally, sections were rinsed in running tap water, air-dried, and coverslipped using Permount.
Morphological analysis
Five coronal sections of each rat in each group were studied quantitatively. Neuronal counts in hippocampus and substantia nigra were performed by eye using a 40× and 20× magnification, respectively, with final field 255 μm2. The observer was blind to the treatment at the time of analysis. Viable stained neurons were identified on the basis of a stained soma with at least two visible processes. Counts were made in five adjacent fields, and the mean number extrapolated to give total number of neurons per 255 μm2. All data are represented as number of neurons per 255 μm2.
Determination of oxidative stress markers
Hippocampus was isolated and prepared as a hippocampal homogenate, and the determination of the oxidative stress markers in hippocampus was performed. MDA level was indirectly estimated by determining the accumulation of thiobarbituric acid reactive substances. 25 To determine the activities of antioxidant enzymes, including SOD, CAT, and GSH-Px, hippocampus of each rat was weighed and homogenized with a buffer consisting of 10 mM sucrose, 10 mM Tris-HCl, and 0.1 mM EDTA (pH 7.4). Then, a hippocampal homogenate was centrifuged at 3000 g at 4°C for 15 minutes. The supernatant was separated and used for bioassays. The activity of SOD was determined using a xanthine/xanthine oxidase system for the production of superoxide radical and subsequent measurement of cytochrome c as a scavenger of the radicals. Optical density was determined using a spectrometer (UV-1601; Shimadzu) at 550 nm. 26 SOD activity was presented as units per milligram of protein (U/mg protein). One unit of enzyme activity was defined as the quantity of SOD required to inhibit the reduction rate of cytochrome c by 50%. CAT activity in the supernatant was measured by recording the reduction rate of H2O2 absorbance at 240 nm. 27 The activity of CAT was expressed as μmol H2O2/min/mg protein. GSH-Px was determined using t-butyl hydroperoxide as a substrate. The optical density was spectrophotometrically recorded at 340 nm and expressed as U/mg protein. 28 One unit of the enzyme was defined as micromole of reduced nicotinamide adenine dinucleotide phosphate oxidized per minute.
Statistical analysis
Data are expressed as mean ± standard error of the mean and analyzed statistically by one-way ANOVA, followed by the post hoc (least significant difference) test. The results were considered statistically significant at p < 0.05.
Results
HPLC analysis
The finger print chromatogram of CP1 is shown in Figure 1. It was found that the combined extract contained gingerol and quercetin at a concentration of 138.94 and 87.83 mg/g of the combined extract, respectively.

High-performance liquid chromatography chromatogram of the combined extract of Zingiber officinale and Cyperus rotundus
Antioxidant activity and AChE inhibition of CP1
In the first part of this study, the antioxidant effect of C. rotundus, Z. officinale, and the combination extract of C. rotundus and Z. officinale (CP1) was determined using DPPH and FRAP assays. In addition, the AChE inhibition was also determined using Ellman's colorimetric method. The results are shown in Table 1. Interestingly, the current data clearly demonstrated that the combination extract of C. rotundus and Z. officinale (CP1) had lower IC50 of FRAP (1.743 mg/mL), DPPH (1.008 mg/mL), AChEI (0.100 mg/mL), and MAO-BI (0.100 mg/mL) than those of C. rotundus or Z. officinale extracts.
DPPH, 2,2-diphenyl-1-picrylhydrazyl; FRAP, ferric reducing antioxidant power.
Effect of CP1 on spatial memory of 6-OHDA lesion rats
Figure 2 shows that the administration of 6-OHDA significantly enhanced escape latency (p < 0.001 all; compared to vehicle+saline group), but decreased retention time (p < 0.001 all; compared to vehicle+saline group) at the 7- and 14-day period. Both vitamin C and L-DOPA treatments significantly improved the reduction of escape latency induced by 6-OHDA (p < 0.001 all; compared to vehicle+6-OHDA group) throughout the experimental period. Both mentioned substances also mitigated the decreased retention time induced by 6-OHDA at 14 days of treatment (p < 0.05, 0.01, respectively; compared to vehicle+6-OHDA group). Interestingly, medium and high doses of CP1 significantly mitigated the enhanced escape latency at the 7-day period (p < 0.01 and 0.001, respectively; compared to vehicle+6-OHDA group) and at the 14-day period (p < 0.001 all; compared to vehicle+6-OHDA group). Low dose of CP1 could significantly improve the reduction of escape latency only at 14 days of treatment (p < 0.001; compared to vehicle+6-OHDA group). Moreover, CP1 at all doses also mitigated the decreased retention time induced by 6-OHDA at 14 days of treatment (p < 0.05, 0.01, 0.01, respectively; compared to vehicle+6-OHDA group).
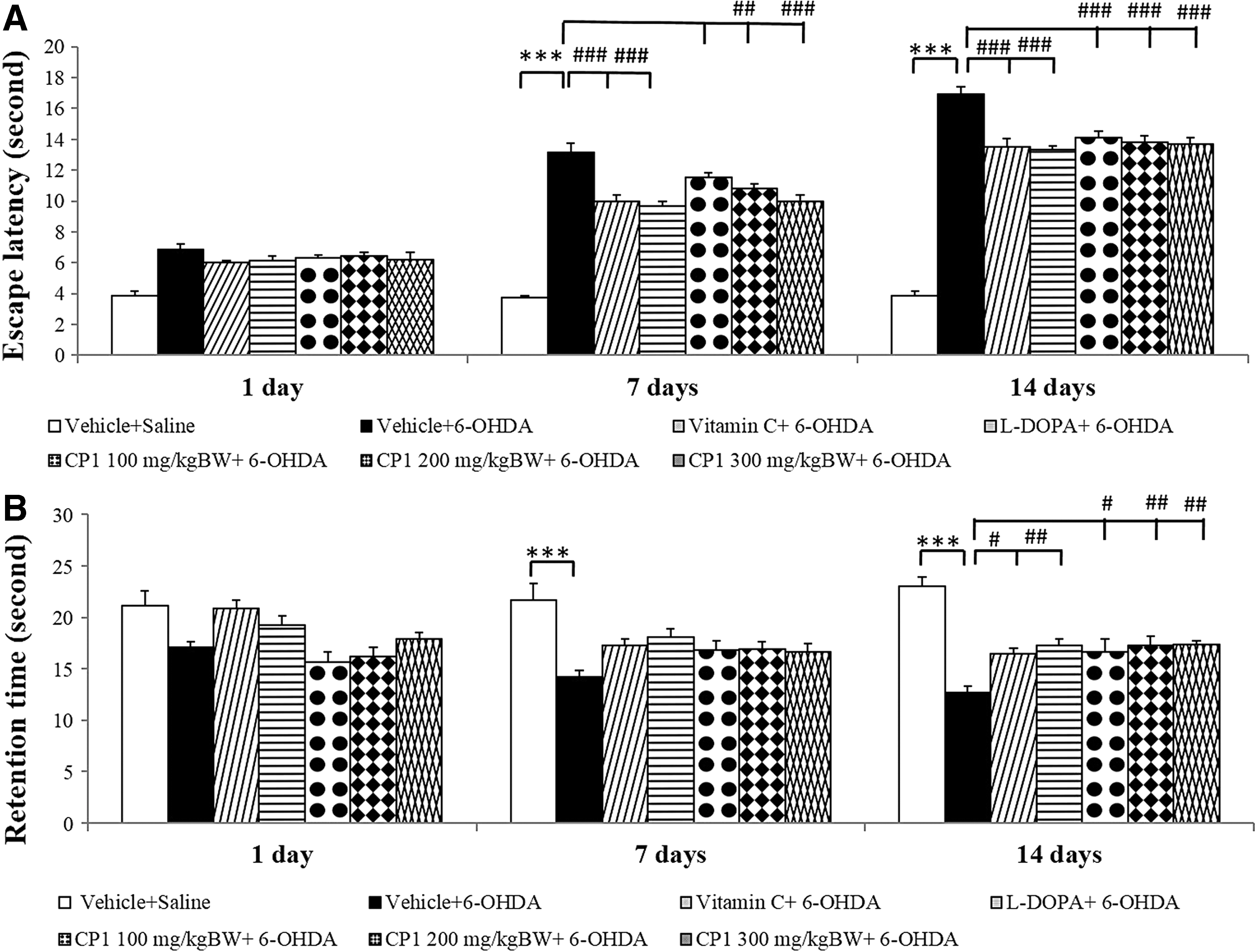
Effect of CP1 on spatial memory using the Morris water maze test in rats subjected to 6-hydroxydopamine hydrochloride (6-OHDA) treatment
Effect of CP1 on apomorphine-induced rotation behavior
Figure 3 shows that the administration of 6-OHDA into the right substantia nigra significantly increased rotation number at the 7- and 14-day period after the 6-OHDA administration (p < 0.001 all; compared to vehicle+saline group). Both vitamin C and L-DOPA treatments significantly suppressed the elevation of rotation number induced by 6-OHDA (p < 0.01 all; compared to vehicle+6-OHDA group) at the 7- and 14-day period after the 6-OHDA administration (p < 0.001 all; compared to vehicle+6-OHDA group). In addition, it was found that the medium and high doses of CP1 significantly suppressed the elevation of rotation number induced by 6-OHDA at only the 14-day intervention period (p < 0.001 all; compared to vehicle+6-OHDA group).
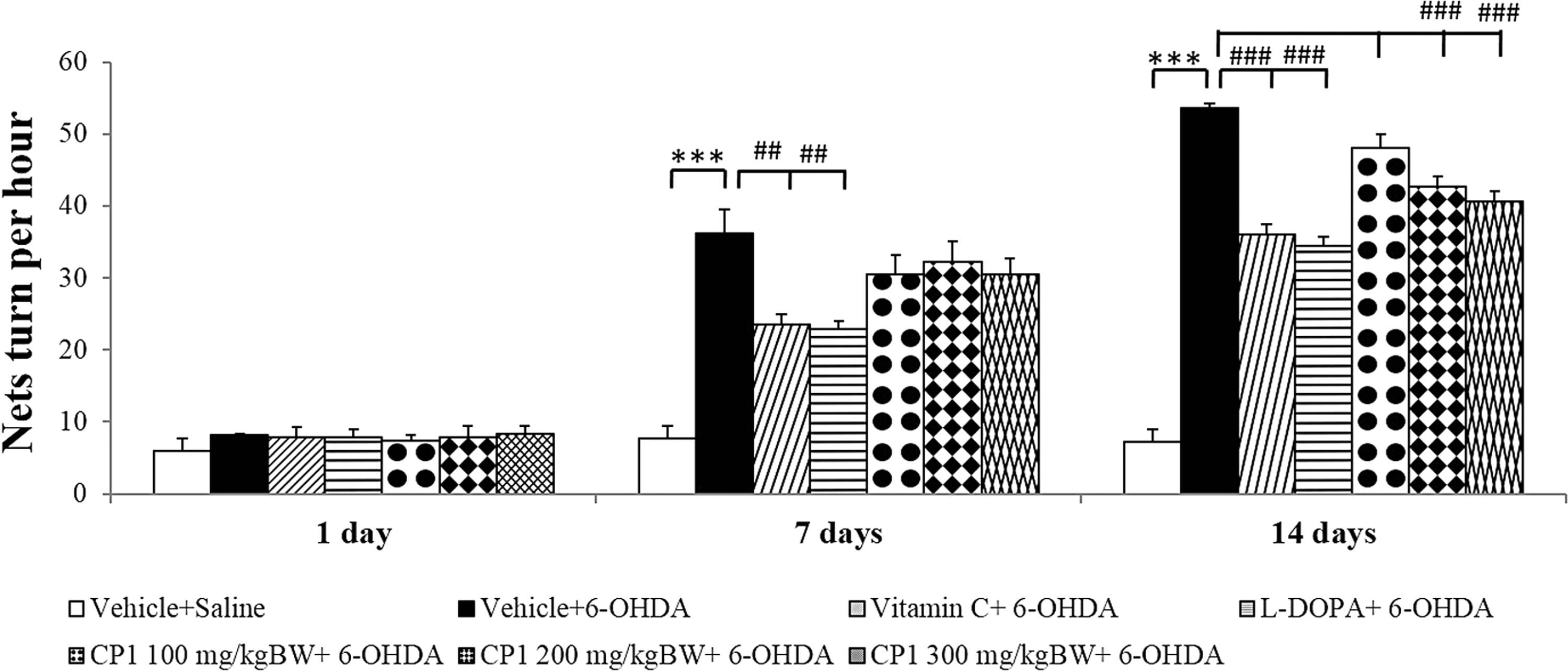
Effect of CP1 on apomorphine-induced rotations in rats subjected to the 6-OHDA. Values are given as mean ± SD (n = 6/group) ***p < 0.001 compared with vehicle+saline group and ## p < 0.01, ### p < 0.001 compared with vehicle+6-OHDA group.
Effect of CP1 on hippocampal neurons
Figure 4 shows the effect of CP1 on neuron density in hippocampus. The results showed that the administration of 6-OHDA into the right substantia nigra induced the decreased survival neuron density in CA1, CA2, and CA3 of hippocampus (p < 0.001 all; compared to vehicle+saline group). It was found that rats subjected to 6-OHDA, which received vitamin C, significantly enhanced neuron density only in CA1 (p < 0.01; compared to vehicle+6-OHDA group). L-DOPA significantly enhanced neuron density in CA1 and CA3 in rats, which subjected to 6-OHDA treatment (p < 0.001 all; compared to vehicle+6-OHDA group). Interestingly, CP1 at all doses significantly mitigated the reduction of neuron density in CA1 (p < 0.05 all; compared to vehicle+6-OHDA group) and CA3 (p < 0.01 all; compared to vehicle+6-OHDA group) in rats, which received 6-OHDA. No significant changes were observed in CA2 and dentate gyrus.
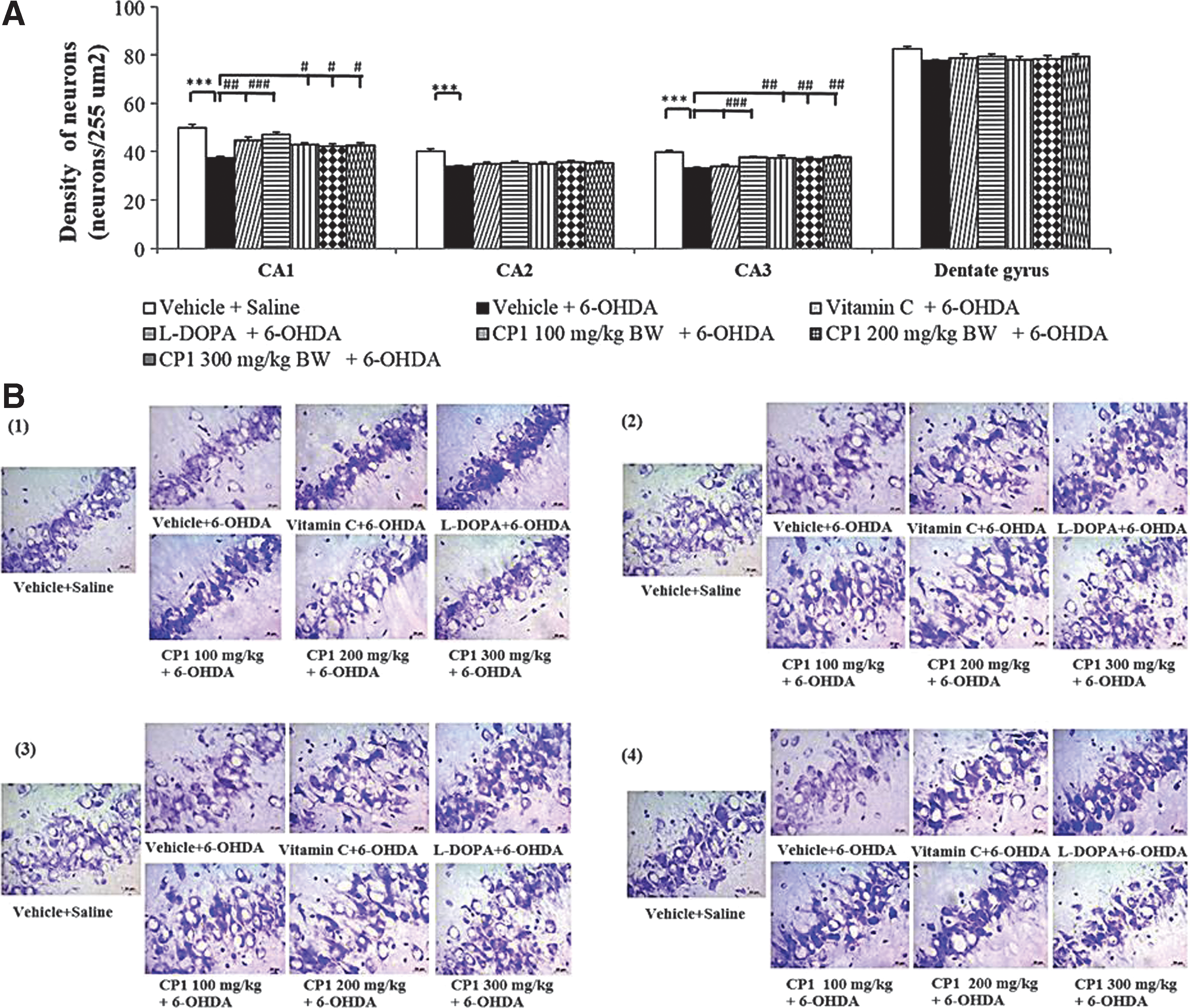
The effect of CP1 on neuron density in various subregions of hippocampus, including CA1, CA2, CA3, and dentate gyrus of rats subjected to 6-OHDA.
Effect of CP1 on tyrosine hydroxylase-positive neuron
In this part, the density of tyrosine hydroxylase-positive neuron in the substantia nigra was investigated by using the immunohistochemistry technique. The rats subjected to the unilateral lesion of substantia nigra induced by 6-OHDA showed the reduction of dopaminergic neuron in the substantia nigra (p < 0.001; compared to vehicle+saline group). Both vitamin C and L-DOPA treatments significantly enhanced the reduction of dopaminergic neuron induced by 6-OHDA (p < 0.001 all; compared to vehicle+6-OHDA group). Interestingly, CP1 at doses of 200 and 300 mg/kg BW also significantly attenuated the reduction of dopaminergic neuron density (p < 0.05, 0.01, respectively; compared to vehicle+6-OHDA group) while no significant change was observed in the low dose of CP1 group as shown in Figure 5.

The effect of CP1 on tyrosine hydroxylase-positive neurons in rats subjected to 6-OHDA treatment.
Effect of CP1 on AChE and MAO-B activities
Since acetylcholine and MAO-B had been reported to play the crucial roles on cholinergic and dopaminergic function, this study also focused on the effect of CP1 on the activities of AChE and MAO-B. The results are shown in Figure 6. Rats that exposed to 6-OHDA showed the elevation of AChE in the hippocampus and striatum (p < 0.001 and 0.05, respectively; compared to vehicle+saline). However, the elevated AChE in the hippocampus was reversed by L-DOPA, vitamin C, and all doses of CP1 (p < 0.001 all; compared to vehicle+6-OHDA group). No significant changes of AChE induced by 6-OHDA in the striatum were observed in rats treated with L-DOPA, vitamin C, and all doses of CP1.

Effect of CP1 on the activity of acetylcholinesterase (AChE) in the hippocampus and striatum. Values are given as mean − SD (n = 6/group) *p < 0.05, ***p < 0.001 compared with vehicle+saline group and ### p < 0.001 compared with vehicle+6-OHDA group.
Figure 7 shows the effect of CP1 on MAO-B activity in hippocampus and striatum. Rats that subjected to the unilateral lesion of substantia nigra induced by 6-OHDA demonstrated the significant reduction of MAO-B in the striatum (p < 0.01; compared to vehicle+saline group). In addition, it was found that L-DOPA treatment significantly mitigated the elevation of MAO-B activity in striatum (p < 0.05; compared to vehicle+6-OHDA group). Interestingly, the high dose of CP1 significantly mitigated the elevation of MAO-B activity in striatum (p < 0.05; compared to vehicle+6-OHDA group). However, no significant change was observed in hippocampus.
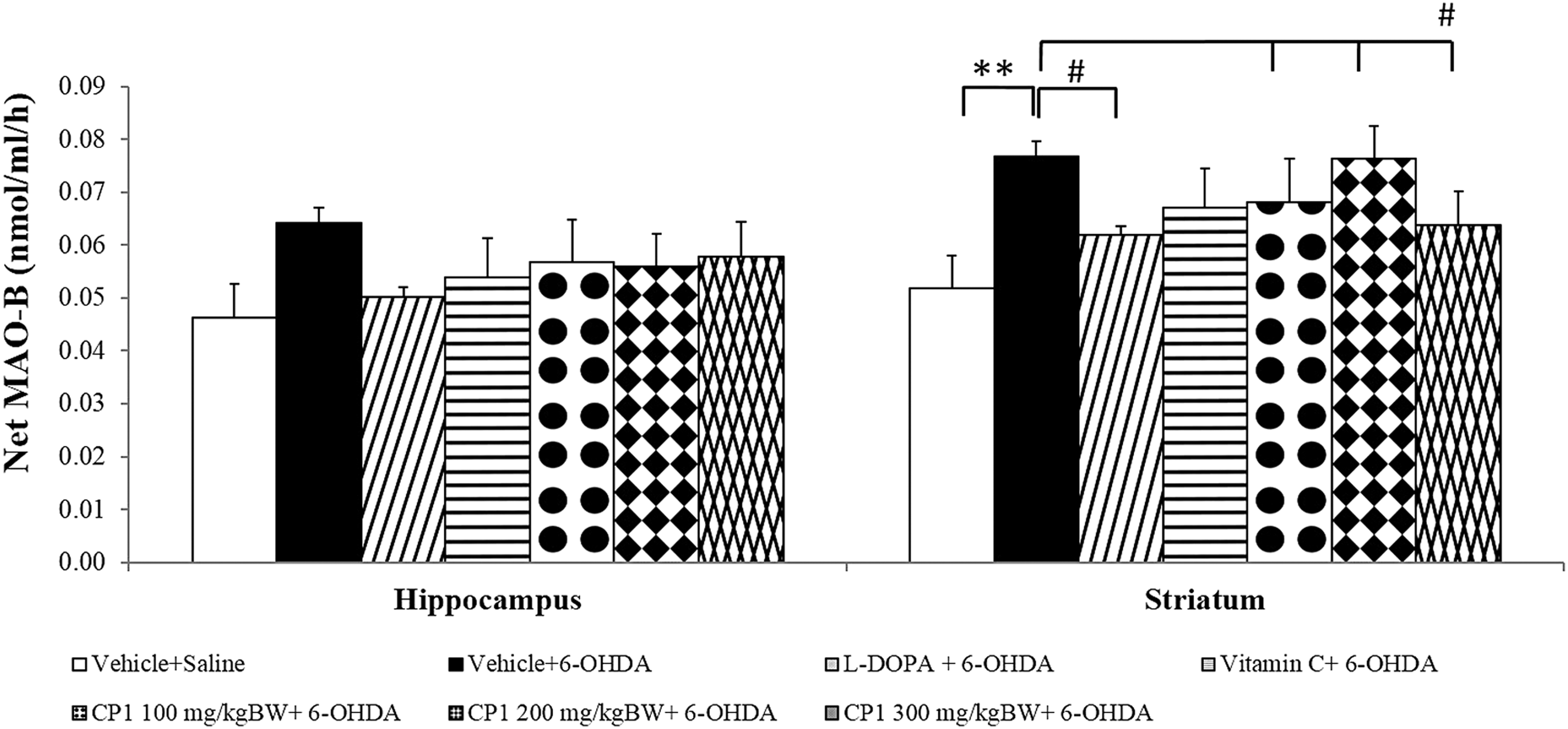
Effect of CP1 on the activity of monoamine oxidase B (MAO-B) in the hippocampus and striatum of rats subjected to 6-OHDA rats. Values are given as mean ± SD (n = 6/group) **p < 0.01 compared with vehicle+saline group and # p < 0.05 compared with vehicle+6-OHDA group.
Effect of CP1 on oxidative stress markers
The effects of CP1 on oxidative stress markers, including the level of MDA and the activities of SOD, CAT, and GSH-Px in the hippocampus and striatum, were also evaluated. The results are shown in Table 2. It was found that 6-OHDA injection into the right substantia nigra significantly increased MDA level, but decreased the activities of CAT and GSH-Px in the hippocampus (p < 0.001, 0.05 and 0.01, respectively; compared to vehicle+saline group). However, the elevation of MDA level in the hippocampus was mitigated by L-DOPA, vitamin C, and CP1 at doses of 100 and 200 mg/kg BW (p < 0.01, 0.01, 0.05, and 0.05, respectively; compared to vehicle+6-OHDA group). No significant changes of CAT, SOD, and GSH-Px in the hippocampus were observed in rats subjected to 6-OHDA, which treated with L-DOPA, vitamin C, and all doses of CP1. In addition, rats that exposed to 6-OHDA showed the elevation of MDA level, but decreased the activities of CAT, SOD, and GSH-Px in the striatum (p < 0.001, 0.001, 0.01, and 0.01, respectively; compared to vehicle+saline group). L-DOPA treatment significantly mitigated the elevation of MDA level and the reduction of CAT activity in the striatum (p < 0.05 all; compared to vehicle+6-OHDA group). Interestingly, medium dose of CP1 significantly mitigated the reduction of CAT activity in the striatum (p < 0.05; compared to vehicle+6-OHDA group).
Values are given as mean ± standard deviation (n = 6/group).
p < 0.05, ** p < 0.01, *** p < 0.001 compared with vehicle+saline group and # p < 0.05, ## p < 0.01, ### p < 0.001 compared with vehicle+6-OHDA group.
CAT, catalase; 6-OHDA, 6-hydroxydopamine hydrochloride; GSH-Px, glutathione peroxidase; MDA, malondialdehyde; SOD, superoxide dismutase.
Discussion
In this study, it had been clearly demonstrated that the combination of C. rotundus and Z. officinale (CP1) at a ratio used in this study showed higher antioxidant activities and AChEI and MAOI effects than Z. officinale or C. rotundus alone. It also attenuated the degeneration of neurons, and cholinergic neurons in the hippocampus. In addition, the degeneration of dopaminergic neurons in the substantia nigra was also observed. Moreover, the improved both motor and nonmotor impairments were also observed. It has been demonstrated that 6-OHDA induces nigrostriatal damage and the impairments of both motor and nonmotor symptoms such as cognitive deficit similar to that observed in patients with PD. 29 –33 Therefore, it has been used as a validated model of PD. The current results also confirm these changes. It was demonstrated that the administration of 6-OHDA into the substantia nigra produced the motor and memory deficit, the elevation of MAO-B, AChE, and oxidative stress in the hippocampus and striatum. Based on the connection between the substantia nigra and the limbic system, including the hippocampus, 6 the administration of 6-OHDA into the substantia nigra could induce the dopaminergic damage not only in the substantia nigra but also in striatum and hippocampus, which in turn induced the functional disturbances of the affected areas. 6,34
Accumulative lines of evidence have shown that spatial memory impairment is associated with the neurodegeneration in the hippocampus, 35,36 and the neurodegeneration, which occurs in this area, is under the influence of oxidative stress status in the mentioned area. 37 –39 It has been shown that all doses of CP1 used in this study enhance spatial memory and neuron density in the hippocampus. In addition, the cholinergic function and oxidative stress status in the hippocampus also increase in CP1-treated rats. Based on the crucial role of oxidative stress status on the neurodegeneration and the role of neurodegeneration on spatial memory impairment mentioned earlier, it has been suggested that CP1 improve oxidative stress status reflected by the decreased MDA level giving rise to the increased neuron density in both CA1 and CA3, the areas contributing the role on encoding and retrieval, 40 which in turn enhance spatial memory. Although CP1-treated rats at all doses used in this study decreased MDA level, no changes of SOD, CAT, and GSH-Px activity in hippocampus were observed in rats, which received CP1. Therefore, the reduction of MDA level might also involve other factors rather than the increase of enzymatic scavenging enzymes such as the nonenzymatic antioxidant system 41 and the decreased oxidative stress formation. 42 Rats that received CP1 at a high dose also showed the increased neuron density in CA1 and CA3 while no improved oxidative stress status in hippocampus was observed. Therefore, other factors such as the decreased apoptosis might also involve. 43 In addition, it has been reported that the suppression of AChE, which in turn increased cholinergic function in hippocampus, also plays the pivotal role on the increased learning and memory impairment in the Parkinson-like condition induced by 6-OHDA. 6 Therefore, the alteration of AChE in hippocampus of rats treated with CP1 was also investigated. The current data also revealed that all doses of CP1 also suppressed AChE in hippocampus. Therefore, the suppression of AChE in hippocampus might also play a role on the improved spatial memory impairment in 6-OHDA lesion rats in this study.
CP1 at doses of 200 and 300 mg/kg BW also improved motor symptom and dopaminergic neuron density in the substantia nigra of 6-OHDA lesion rat, an animal model of PD. It has been reported that the improved dopaminergic function 44 plays an important role on the improved motor behaviors disorder such as rotational behavior. Therefore, the improved rotational behaviors in CP1-treated rats might be due to the increased dopaminergic neurons in the substantia nigra, an important area involving the pathogenesis of PDs. On the basis of previous findings, which demonstrated the role of apoptotic pathway in the degeneration of dopaminergic neurons in the substantia nigra, 43 it has been suggested that the enhanced dopaminergic neuron density in substantia nigra in 6-OHDA lesion rats treated with CP1 at medium and high doses may be due to the decreased apoptosis in the substantia nigra. However, this required further investigation. Since oxidative stress and MAO-B suppression have been reported to be the targets for therapeutic strategy against PD, 45 the alterations of oxidative stress markers, and the activity of MAO-B in striatum, a key area in regulating the basal ganglia function was also investigated. However, only the rats that received CP1 at a high dose showed the suppression of MAO-B activity in the striatum. Although rats that received medium dose of CP1 showed the enhanced CAT activity in the striatum, no significant change of MDA level was observed. Therefore, the suppression of MAO-B in striatum induced by a high dose of CP1 might increase dopaminergic function, which in turn improved the function of basal ganglia and improved motor symptom in the animal model of PD. The improved oxidative stress was less likely to play a role in the improved motor symptom induced by CP1 both at medium and at high doses. No dose-dependent manner of CP1 was observed in this study because the relationships between the concentration of CP1 and the observed parameters were not the simple linear relationships. Numerous factors play roles on the learning and memory and neurodegeneration.
Our study has clearly demonstrated that CP1 showed the improvement of both motor symptoms and nonmotor symptoms such as cognitive function in 6-OHDA rats as similar as those observed in 6-OHDA rats treated with either vitamin C or L-DOPA. It has been found that 6-OHDA improves motor symptoms by enhancing the available dopamine in the nigrostriatal pathway. 46 In addition, it can also exert antiparkinsonian action through its nondopaminergic effect. 47 Our findings also demonstrated that L-DOPA could improve both dopaminergic function and oxidative stress status in the striatum, which in turn improved motor symptom in 6-OHDA rat model of PD. Therefore, these findings were also in agreement with the previous data mentioned earlier. Although 6-OHDA could produce a great advantage for PD, the long-term treatment could produce dyskinesia. 48 When compared to vitamin C, it was found that the benefits of both vitamin C and CP1 were in the same magnitude. However, the daily recommended of vitamin C is 120 mg/day, 49 while the effective dose of this substance that can improve PD obtained from this study is 200 mg/(kg·day) or around 12,000 mg/day for a 60-kg person. Therefore, this appeared to be beyond the recommended dose and could produce undesired effects such as urinary stone. 50 –52 In addition, the pro-oxidant effect of vitamin C was also reported. 53 Since our pilot study demonstrated that LD50 through the oral route of CP1 was more than 5 g/kg BW, CP1 was practically safe for the consumption based on the Hodge and Sterner Scale. 54 Therefore, CP1, the novel food supplement, may be served as the potential alternative choice for the patients with PD.
It was previously reported that C. rotundus extract could improve neurodegeneration in 6-OHDA rats 55 and improved memory impairment induced by hypocholinergic function. 56 Z. officinale extract also improved oxidative stress status and cholinergic function in the hippocampus, giving rise to the decrease of neurodegeneration and the improved memory. 11,13 However, our in vitro data clearly demonstrated that the combination extract of both plants (CP1) exerted higher antioxidant and AChEI activities than C. rotundus or Z. officinale alone. Therefore, only CP1 were treated in 6-OHDA rat model of PD. However, the in vivo effect of either C. rotundus or Z. officinale in the animal model of PD should be further explored to confirm the ingredients that are responsible for the beneficial effects of CP1.
Based on the high content of gingerol in CP1 and the antiapoptotic effect 57 of gingerol and the antiparkinson effect of quercetin, a constituent content in CP1, it has been suggested that the improved both motor and nonmotor symptoms of PD induced by CP1 may be partly associated with gingerol and quercetin. The interaction effects of both substances are also possibly to contribute to the role on the antiparkinson effect of CP1. However, the effect of other ingredients such as zingerone, which could decrease dopamine depression in the striatum, 58 and the degradation product of gingerol still could not be omitted.
Conclusion
The possible mechanisms of CP1 is proposed in Figure 8. CP1 is the potential candidate to improve both motor and nonmotor symptoms such as memory impairment in PD. The memory enhancing effect might occur through the suppression of AChE, which in turn increased cholinergic function in the hippocampus and the decreased oxidative stress status, which in turn increased neuron density in CA1 and CA3 in the hippocampus leading to the improved memory impairment. The motor improvement induced by CP1 might occur through the enhanced dopaminergic neuron density in the substantia nigra, which in turn improved dopaminergic function and motor disorder. In addition to the enhanced density of dopaminergic neurons in the substantia nigra, the improved dopaminergic function in striatum induced by CP1 especially at a high dose also occurred through the suppression of MAO-B in the striatum. Therefore, CP1 might be served as the potential phytotherapy against PD or as an adjuvant therapy against PD. However, further researches concerning the active ingredient, subchronic toxicity, and the interaction between CP1 and other medicines are essential before moving to clinical trial.

Schematic diagram concerning the possible mechanisms to improve memory and motor disorders of CP1 in hemiparkinsonian rats induced by the unilateral injection of 6-OHDA into the right substantia nigra. Color images available online at
Footnotes
Acknowledgments
This study was supported by the Higher Education Research Promotion and National Research University Project of Thailand, Office of the Higher Education Commission, through the Food and Functional Food Research Cluster of Khon Kaen University, the National Research Council of Thailand, and Research Division of Faculty of Medicine and the Integrative Complementary Alternative Medicine Research and Development Center, Khon Kaen University, Thailand.
Author Disclosure Statement
No competing financial interests exist.