
Abstract
Autologous bone marrow mesenchymal stem cell (BM-MSC) transplantation is a novel strategy for treating ischemic heart disease. However, limited benefits have been reported in aging patients. Rejuvenation of aged human BM-MSCs (hBM-MSCs) could be a means to improve the efficacy of stem cell transplantation in older patients. While it has been shown that sirtuin 6 (SIRT6) is an important antiaging factor in various cells, the role of SIRT6 in hBM-MSCs remains unknown. The hBM-MSCs from different ages were cultured for quantifying SIRT6 expression by mRNA and Western blotting. The cell proliferative and migration abilities were evaluated by BrdU staining, cell growth curves, and scratch assay. Senescence-associated β-galactosidase (SA-β-Gal) activity and aging-associated p16 (cyclin-dependent kinase inhibitor 2A) expression were also quantified. The knockdown of SIRT6 in hBM-MSCs was used to investigate its impact on aging. SIRT6 expression increased with age, while the proliferative and migration abilities of aged hBM-MSCs were decreased compared with young cells. Knockdown of SIRT6 impaired the proliferative, migration, and oxidative stress resistance potentials of BM-MSCs. SA-β-Gal activity and p16 expression were increased in aged cells compared with young ones and in siRNA SIRT6 knockdown cells compared with their controls. Aging results in compensatory overexpression of SIRT6 in hBM-MSCs. Downregulation of SIRT6 in these cells resulted in less cell proliferation and migration but increased SA-β-Gal activity and p16 expression. These results suggest that SIRT6 regulates the aging process in hBM-MSCs and could serve as a target for their rejuvenation.
Introduction
D
However, clinical trials of cardiac cell therapy showed limited benefits in aging patients 4 because stem cells from aged donors have limited proliferative, survival, and regenerative capacity. 5 Transplantation of human BM-MSCs (hBM-MSCs) from aged patients has a lower capacity to repair injured myocardium and restore cardiac function after a rat MI compared with that of young hBM-MSCs. 6 To improve the efficacy of cell therapy in aged patients, research has focused on identifying ways to rejuvenate senescent BM-MSCs.
The antiaging factor sirtuin 6 (SIRT6) has been shown to be a critical regulator of transcription, genome stability, telomere integrity, DNA repair, and metabolic homeostasis. 7 SIRT6 knockout mice display dramatic phenotypes of accelerated aging. 8 In contrast, overexpression of SIRT6 in Hutchinson–Gilford progeria syndrome cells impedes premature senescence and formation of dysmorphic nuclei. 9 Despite its importance to aging, SIRT6 expression in hBM-MSCs from different aged donors and its effect on cellular function have not been investigated.
In the current study, we investigated the expression of SIRT6 in hBM-MSCs derived from infant, young, and aged populations. We found increased SIRT6 expression during aging. Knockdown of SIRT6 in cells decreased cellular, proliferative, and migration abilities but increased cell death and senescence-associated β-galactosidase (SA-β-Gal) activity as well as aging-associated p16 (cyclin-dependent kinase inhibitor 2A) expression. Our results suggest that compensatory overexpression of SIRT6 plays an important role in regulating cellular aging processes. SIRT6 may thus be a potential target for the rejuvenation of aged stem cells.
Materials and Methods
Ethics, consent, and permissions
The use of human cells was approved by the Ethics Board Committee of the Shanxi Medical University and conformed to the principles of the Declaration of Helsinki. All patients provided written informed consent.
Harvest and culture of hBM-MSCs
Patient characteristics are summarized in Table 1. The hBM was harvested from the posterior superior iliac spine of patients. Briefly, 1 mL hBM was cultured in T-25 flasks with 5 mL Iscove's Modified Dulbecco's Medium (IMDM) with 10% fetal bovine serum (FBS). When the cultured cells reached a confluence of 80%–90%, cells were subcultured in a density of 5000 cells/cm2. The cells were used for all experiments at the fourth passage.
Quantification of SIRT6 and p16 mRNA and proteins in hBM-MSCs
SIRT6 and p16 mRNA in cultured cells were quantified using real-time polymerase chain reaction (RT-PCR) with the following primers (SIRT6: forward: 5′-GTCTTCCAgTgTggTgTTCCA-3′, reverse: 5′-CCCAgTCTAggATggTgTCC-3′; p16: forward: 5′-CTCACCATggATgATgATATCgC-3′, reverse: 5′-AggAATCCTTCTgACCCATgC-3′; and β-actin (ACTB): forward: 5′-CCTgAAgTACCCCATCgAgC-3′, reverse: 5′-CTCTTgCTCgAAgTCCAggg-3′). RT-PCR products were evaluated using ImageJ software (NIH), and relative levels of SIRT6 mRNA were presented as the ratio of SIRT6 to ACTB (n = 4/group).
The SIRT6 protein levels were quantified using Western blot analysis with antibodies against SIRT6 (1:250; Abcam) and ACTB (1:2000; Abcam). The density of protein bands was analyzed with ImageJ software, and the ratios of SIRT6 to ACTB were then calculated. The p16 protein levels were quantified using immunocytofluorescence. Cultured cells were fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS) and then incubated with mouse antibodies against p16 (1:100 in PBS at 4°C overnight in humidified box). The slides were incubated with Cy3-conjugated secondary antibody (1:200 in PBS for 1 hour at room temperature in humidified box avoiding light). The slides were then counterstained with 4′,6-diamidino-2-phenylindole (DAPI; 100 ng/mL in PBS at 37°C for 10 minutes in dark). The percentage of p16-positive cells was quantified by counting cells with red color compared to total cells in the same field (n = 4/group).
Identification of SIRT6 in hBM-MSCs
Localization of SIRT6 in cultured hBM-MSCs was evaluated using immunohistochemistry. The cultured cells were fixed in 4% paraformaldehyde in PBS and then incubated with rabbit anti-SIRT6 antibody (1:200 in PBS overnight in humidified box at 4°C). The slides were incubated with FITC-conjugated secondary antibody (1:200 in PBS at 37°C for 1 hour in humidified box avoiding light). The slides were then counterstained with DAPI (100 ng/mL in PBS at 37°C for 10 minutes in dark). The SIRT6-positive cells were photographed.
Cell growth
To evaluate the proliferative ability of hBM-MSCs, we performed the BrdU labeling technique and cell growth curve assay. For the BrdU assay, cells were seeded on 24-well plates. When cells reached 40%–50% confluence, they were further cultured in the BrdU medium (IMDM with 10% FBS containing 3 μg/mL BrdU) for 3 days. Cells were then immunostained for BrdU. The BrdU-positive cells were counted by ImageJ software, and the percentage of BrdU-positive cells was calculated. For the cell growth curve counting, 7000 cells were seeded on 12-well plates (there were three replicative wells in each sample). The cell numbers were counted from 0 to 6 days every 2 days to establish the growth curve.
Cell migration assay
To measure the migration ability of hBM-MSCs, cells were cultured on six-well plates. When the cell cultures reached ∼100% confluence, a scratch wound was made through the middle of the wells using a 20-μL pipette tip. Images were taken using an inverted microscope (Olympus). The percentage of migration rate was measured and analyzed 24 hours after scratch by ImageJ software (n = 4/group).
Cell death and apoptosis assay
The hBM-MSCs were exposed to H2O2 (1000 μM) for 6 hours and harvested. Apoptosis was tested by flow cytometry (Beckman Coulter) staining for Annexin V and propidium iodide (PI) (Life Technologies). The percentage of dead (PI-positive and Annexin V-negative cells) and apoptotic (Annexin V-positive cells) cells was quantified by flow cytometry (n = 4/group).
Measurement of SA-β-Gal activity
SA-β-Gal activity was evaluated using a Senescence Detection Kit (Solaribio) according to the manufacturer's instructions. The percentage of positive cells was counted by counting cells with blue color compared to total cells in the same field (n = 4/group).
Knockdown of SIRT6 in hBM-MSCs
Only one type of siRNA was used to knock down SIRT6 expression in the hBM-MSCs according to the manufacturer's protocol. Briefly, 7000 cells were seeded on 24-well plates in IMDM (FBS and antibiotic free). Cells cultured for 24 hours were transfected with 3 μL 20 μM Oligo RNA-5 carboxyfluorescein (FAM) by 2 μL Lipofectamine 2000 (Invitrogen). The medium was replaced at 6 hours after transfection, and the identification of transfection was performed by a fluorescence microscope (Olympus); the transfection efficiency was quantified by flow cytometry (FCM). The SIRT6 knockdown efficiency in hBM-MSCs was evaluated by RT-PCR (24 hours after transfection) and Western blotting (72 hours after transfection) (n = 4/group).
Statistics
Data are presented as mean ± standard deviation. Statistical analyses were performed using GraphPad Prism 5 (GraphPad). A two-tailed Student's t-test was used for two-group comparisons. All other comparisons among three or more groups were analyzed by one-way ANOVA, followed by Tukey's post hoc tests. Differences were considered statistically significant at p < 0.05.
Results
Identification of hBM-MSCs
The cultured hBM-MSCs were identified using flow cytometry to examine the cell markers. We found that greater than 99% of cells were CD105 and CD44 positive, and there was no significant difference among the infant, young, and aged groups. The percentage of both CD34- and CD45-positive cells was lower than 1%, and there was also no significant difference among the three groups (Fig. 1A). These data suggest that the cells cultured and used in the subsequent experiments were hBM-MSCs.
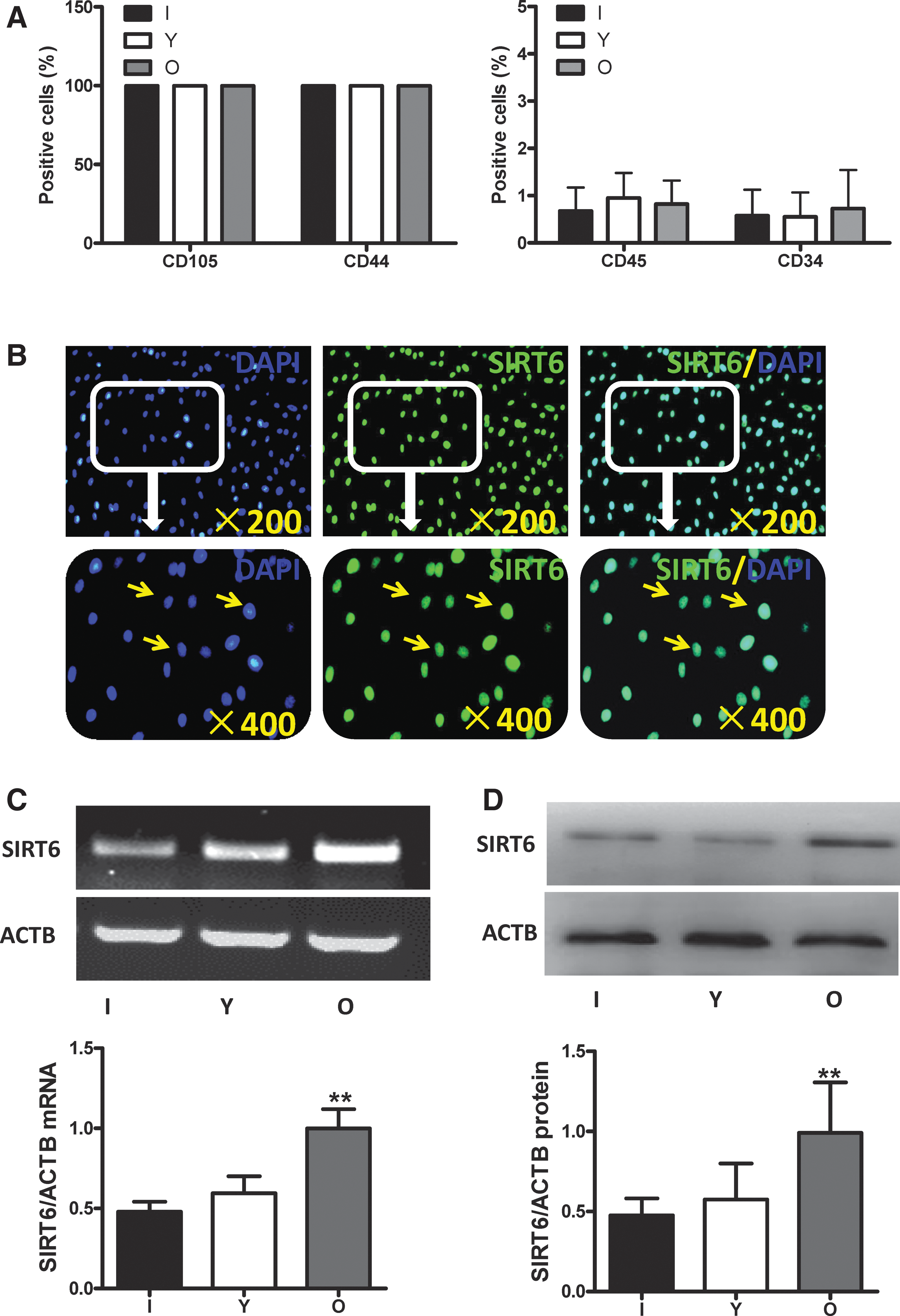
Identification of hBM-MSCs and the expression of SIRT6 in hBM-MSCs.
Age-dependent increase in SIRT6 expression
The expression of SIRT6 in hBM-MSCs among the different age groups was evaluated. SIRT6 is a predominantly nuclear protein as shown by immunocytofluorescent staining (Fig. 1B). The mRNA (Fig. 1C) and protein (Fig. 1D) expression of SIRT6 were significantly higher in the old group compared to the infant and young groups (p < 0.01).
siRNA knockdown of SIRT6 expression
The positivity of siRNA transfection was detected by immunofluorescent analysis for FAM at 6 hours posttransfection. The untransfected cells were negative for FAM staining, but cells in the siRNA control (NC) and siRNA SIRT6 groups were positive (Fig. 2A). When one type of siRNA was transfected into cells from both age groups, we found that the percentage of FAM-positive cells in the old group was significantly lower compared to the young group (Fig. 2B, p < 0.01). The siRNA was found to effectively knock down SIRT6 gene expression. We found that mRNA expression of SIRT6 was significantly decreased in the young and old siRNA groups (using the same siRNA) compared to their respective NC control groups (Fig. 2C, p < 0.01). The SIRT6 protein levels were also significantly decreased in the young and old siRNA groups compared to their respective NC groups (Fig. 2D, p < 0.01).

Identification of siRNA transfection efficiency and SIRT6 expression following siRNA transfection.
Decreased proliferation of hBM-MSCs after SIRT6 knockdown
To determine the impact of age and SIRT6 on the proliferative ability of hBM-MSCs, BrdU staining and cell growth curve counting methods were used. BrdU-labeled cells were observed in the hBM-MSCs of all three age groups (Fig. 3A, arrows). The percentages of BrdU-positive cells were twofold higher in the infant and young groups compared to the aged group (Fig. 3B, p < 0.01). We also found that the growth rates in the infant and young groups were significantly higher compared to the aged group (Fig. 3C, p < 0.01). These data indicate that proliferative ability decreased with aging.
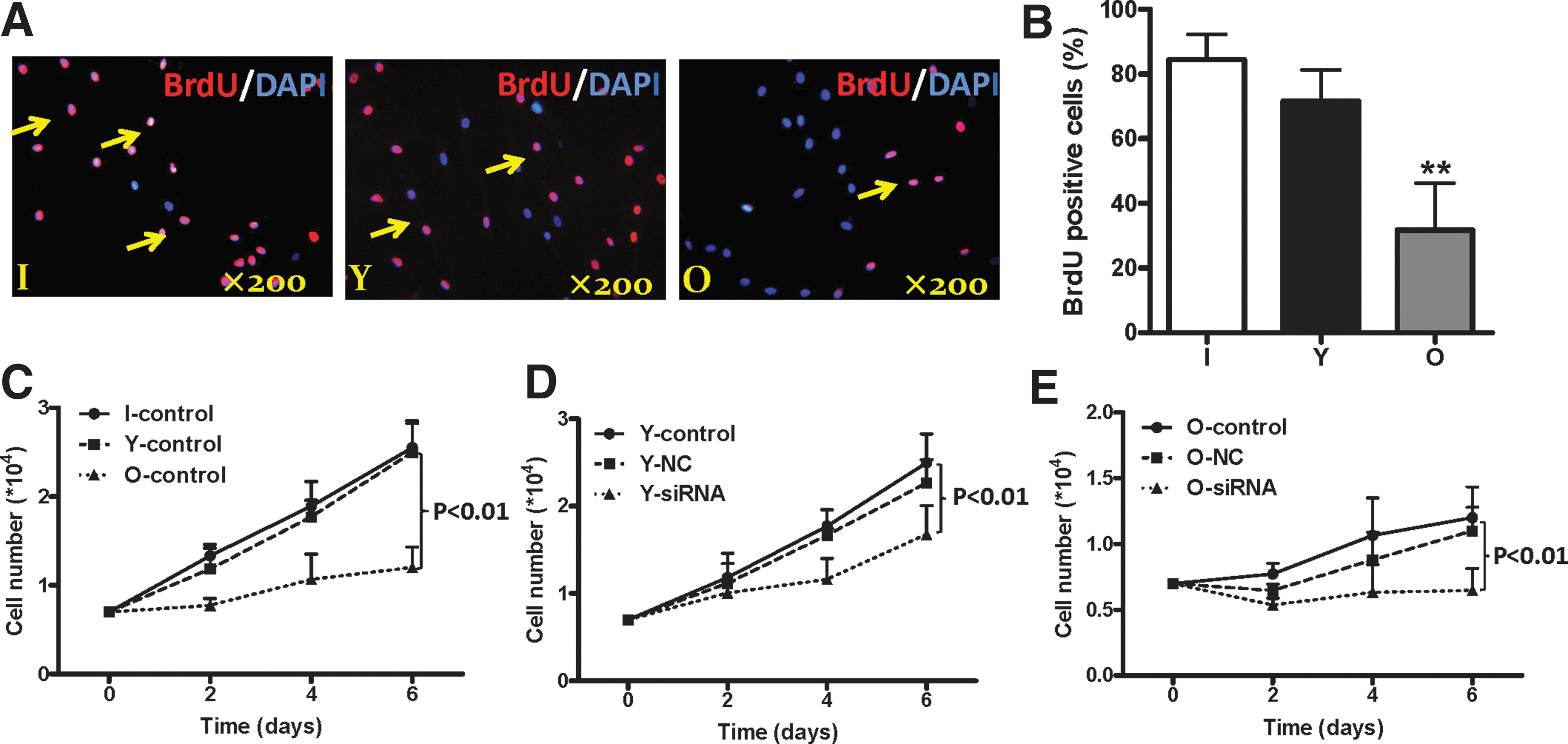
Impact of aging and SIRT6 in the proliferation of the hBM-MSCs.
The SIRT6 levels in young and aged hBM-MSCs decreased after SIRT6 siRNA transfection (Fig. 2). The growth rates in the siRNA groups of young and aged hBM-MSCs were significantly lower compared with the NC control groups (Fig. 3D, E, p < 0.01). Although the proliferative rate in aged cells was significantly lower than that in young cells, we observed almost no cellular growth in aged siRNA knockdown cells (Fig. 3E). These results suggest that SIRT6 plays a significant role in cell growth and proliferation.
Impaired hBM-MSC function after SIRT6 knockdown
To determine the impact of age and SIRT6 on the migration ability of hBM-MSCs, the scratch assay was performed. The cellular migration rate was significantly decreased in aged cells. The migration rates were threefold higher in infant and young cells compared to aged cells (Fig. 4A, p < 0.01). The migration rate of SIRT6 knockdown cells was also significantly decreased relative to control cells but independent of age (Fig. 4B, C). The migration rate of young siRNA cells was significantly lower than their respective NC control cells (Fig. 4B, p < 0.05). Similarly, the migration rate of old siRNA cells was also significantly lower than their respective NC control cells (Fig. 4C, *p < 0.05). These data indicate that aged hBM-MSCs have lower migration ability and that SIRT6 is involved in cell migration capacity.
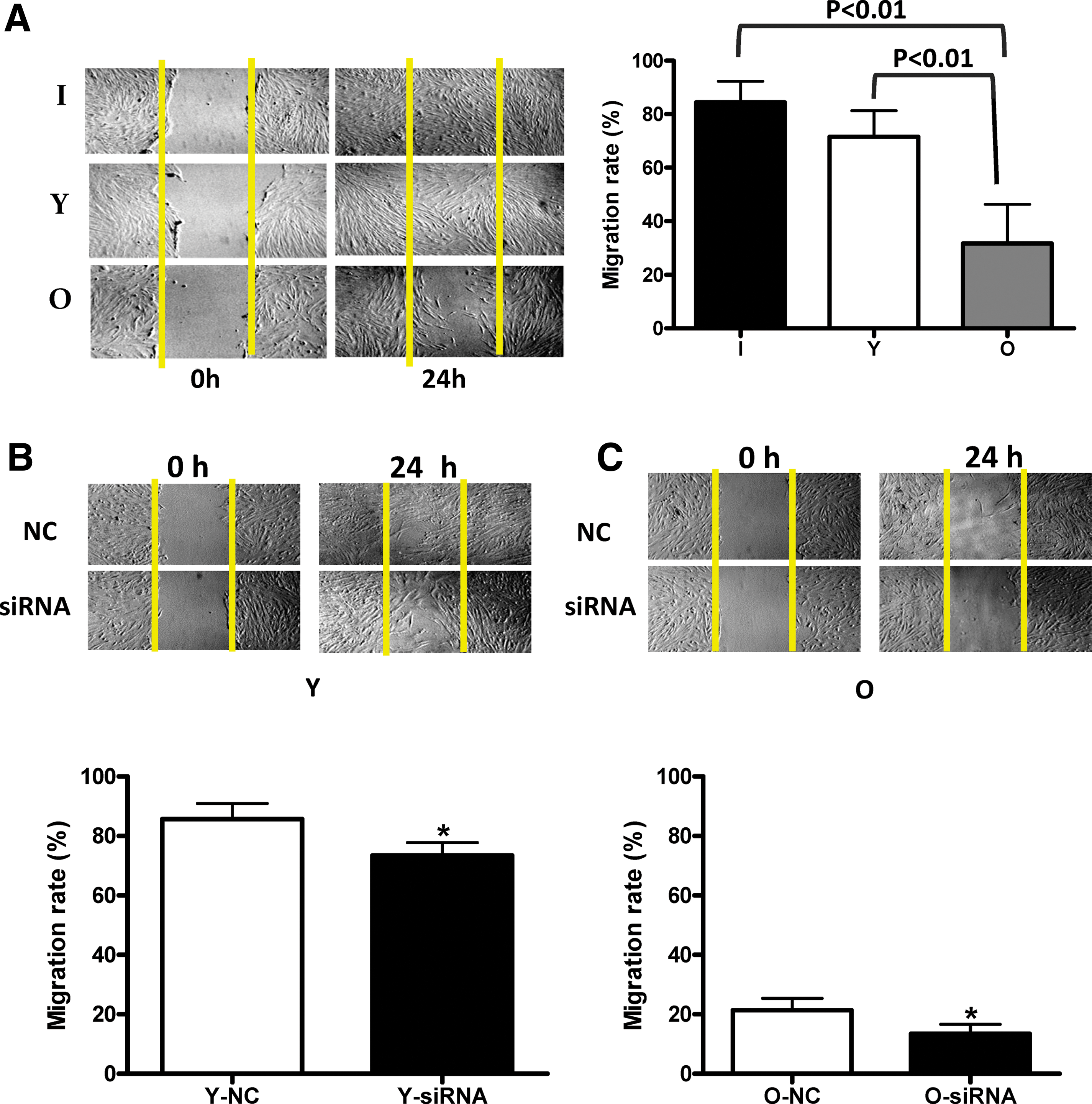
Impact of aging and SIRT6 on the migration of hBM-MSCs.
Effect of SIRT6 on cellular apoptosis
Apoptosis of hBM-MSCs was quantified by flow cytometry to stain for Annexin V and PI. We found that the percentage of apoptotic cells (Annexin V-positive cells) was more than eightfold higher in aged cells compared to infant and young cells (Fig. 5A, right panel, p < 0.05). Despite the higher percentage of dead cells in aged cells compared to infant and young cells, there was no statistical difference among the three groups (Fig. 5A, left panel).

Influences of aging and SIRT6 on antiapoptosis and oxidative stress resistance.
To determine the antiapoptosis effect of SIRT6, hBM-MSCs were knocked down with siRNA. We observed no significant difference in cell apoptosis levels between young siRNA and NC control cells (Fig. 5B, right panel). Similarly, there was also no significant difference in the percentage of dead cells between siRNA and NC control cells (Fig. 5B, left panel). These results indicate that the level of apoptosis in hBM-MSCs increased with age; however, knockdown of intracellular SIRT6 did not affect cellular apoptosis.
To investigate the effect of age on cellular oxidative stress resistance, hBM-MSCs from different age groups were treated with H2O2 for 6 hours, and cell viability was examined by flow cytometry staining with Annexin V and PI. The percentage of dead cells was significantly increased in old cells compared to infant and young cells (Fig. 5C, left panel, p < 0.05), and the percentage of apoptotic cells was eight times greater in aged cells compared to infant and young cells (Fig. 5C, right panel, p < 0.01).
To determine the effect of intracellular SIRT6 on cellular oxidative stress resistance, the young hBM-MSCs were first transfected with SIRT6 siRNA or NC siRNA for 72 hours, which was followed by 6 hours of H2O2 treatment. The percentage of dead cells was more than two times higher (Fig. 5D, left panel, p < 0.05) in the siRNA group compared to the NC control group. However, there was no significant difference in the percentage of apoptotic cells between the NC and siRNA groups (Fig. 5D, right panel). These data suggest that aging increased cellular sensitivity to oxidative stress; however, intracellular SIRT6 levels did not alter cellular sensitivities.
Increased activity of SA-β-Gal in aged and SIRT6 knockdown hBM-MSCs
SA-β-Gal activity is a characteristic feature of cell senescence. Compared to aged hBM-MSCs in the aged group, the percentage of SA-β-Gal-positive cells in the infant and young groups was 16 times lower (Fig. 6A, p < 0.01). After knocking down SIRT6 in young hBM-MSCs, the percentage of SA-β-Gal-positive cells in the siRNA group was more than two times higher than the control group (Fig. 6B, p < 0.05). These results indicate that aging upregulates hBM-MSC senescence and that SIRT6 downregulates cell senescence.
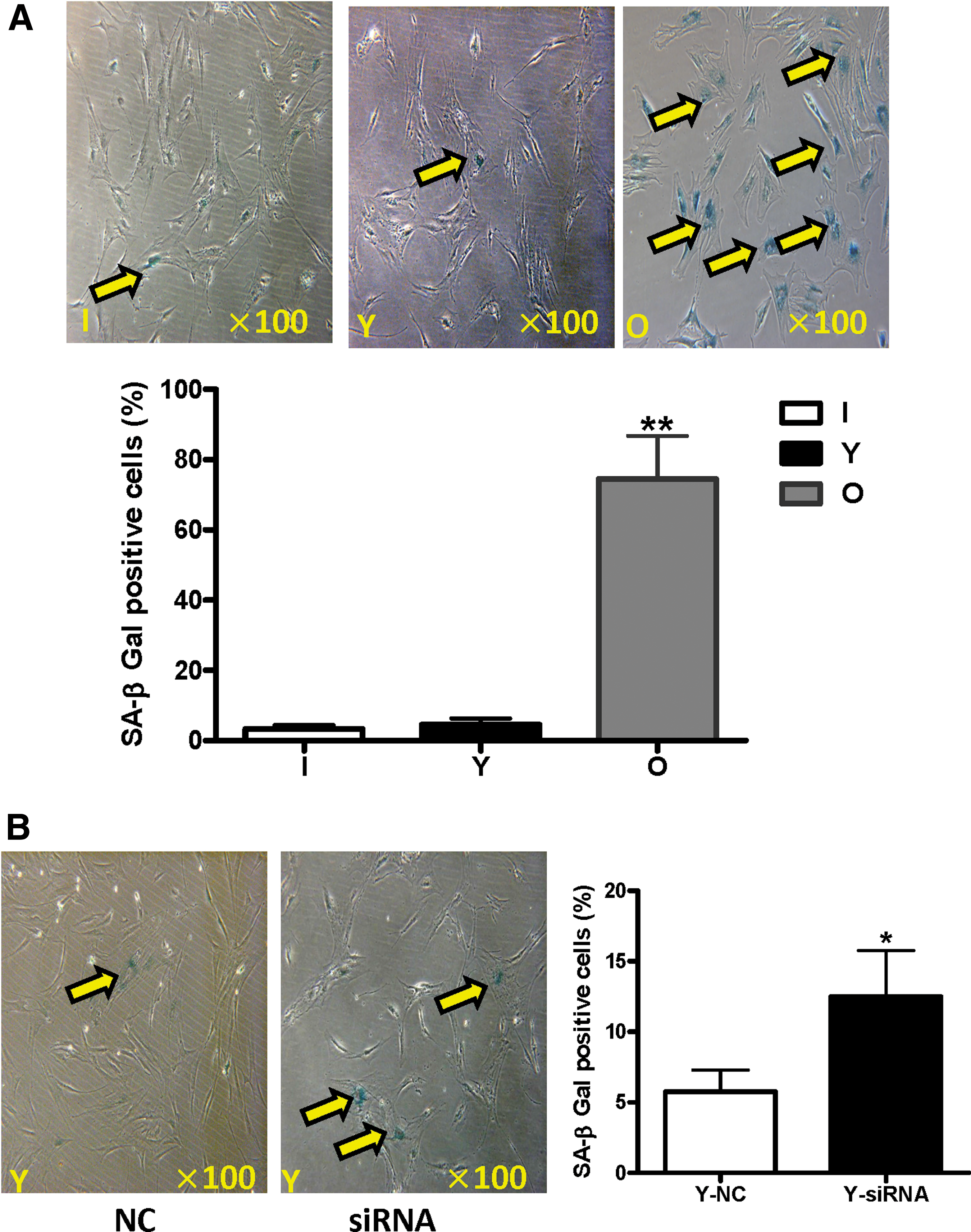
Influences of aging and SIRT6 on SA-β-Gal positivity in hBM-MSCs.
Increased p16 expression in aged and SIRT6 knockdown hBM-MSCs
P16 expression is an important parameter of cell aging. Compared to infant and young hBM-MSCs, the mRNA expression of p16 in the aged group was more than two times higher (Fig. 7A, p < 0.01). After knocking down SIRT6 in young hBM-MSCs, the mRNA expression of p16 in the siRNA group was also more than two times higher than the control group (Fig. 7B, p < 0.05). Similarly, the percentage of p16-positive hBM-MSCs in the aged group was significantly higher compared with the infant and young groups (Fig. 7D, p < 0.01) and significantly higher in SIRT6 knockdown young hBM-MSCs compared with control cells (Fig. 7E, p < 0.01). These results again suggest that aging upregulates hBM-MSC senescence and that SIRT6 downregulates cell senescence.

Influences of aging and SIRT6 on aging-associated p16 mRNA expression and cell positivity in hBM-MSCs.
Discussion
It was found that the proliferative ability, differentiation potential, antiapoptotic capacity, and homing ability of aged BM-MSCs were impaired compared to young BM-MSCs, which thereby reduced the effectiveness of cell therapy. 6,10 –13 SIRT6 has been reported to be a critical regulator of transcription, genome stability, telomere integrity, DNA repair, and metabolic homeostasis. 14 SIRT6-deficient mice display premature aging syndromes and die at about 4 weeks of age. 8 Conversely, overexpression of SIRT6 extended the life span of mice. 15 These previous functional studies suggest that SIRT6 is an age-antagonizing factor as its expression is strongly correlated with aging. However, the effect of SIRT6 on stem cell aging has not been previously investigated. Thus, the current study set out to examine SIRT6 expression levels in human stem cells derived from infant, young, and aged donors and its effect on cellular function.
Interestingly, we found that SIRT6 expression increased with age in hBM-MSCs. This result is in agreement with a previous study by Koltai et al. demonstrating that the expression of SIRT6 was increased in aged skeletal muscle cells compared with young cells 16 and a recent finding of a significant age-related increase in SIRT1 protein (longevity protein similar to SIRT6) expression in human blood serum against oxidative stress in the elderly. 17 However, it was also reported that the expression of SIRT6 was downregulated in human dermal fibroblasts from aged donors. 7
The mechanism responsible for these dissimilar results in SIRT6 expression in aged cells remains unknown. A possible explanation is that increased SIRT6 expression with age is a stem cell-specific phenomenon. In our unpublished preliminary data, we found that in human monocytes isolated from young and old individuals, SIRT6 expression declined with age. However, further evidence is needed to support this postulation. Another potential mechanism for this discrepancy is a compensatory effect in aged individuals. It is well known that the “free radical theory of aging” is the aging mechanism, which hypothesizes that advanced aging leads to an imbalance between the concentrations of free radicals and antioxidants. 18
In our current study, we demonstrated that aged cells had a much lower antioxidant capacity to resist oxidative stress generated by H2O2 than young cells. To offset this aging effect, the SIRT6 protein (acting to promote resistance to oxidative stress) could accordingly increase its expression to compensate for the decrease in antioxidant capacity of aged cells. When we knocked down SIRT6 expression by siRNA, the stem cells became senescent. However, further research is needed to confirm this hypothesis.
In this study, we also investigated the function of SIRT6 in human stem cells. First, in cell growth, it has been reported that the effect of SIRT6 on cell proliferation varies depending on cell type. SIRT6 acts as a suppressor to inhibit the growth of cancer cells. 19 –24 However, in nonmalignant cells, the effect of SIRT6 is to promote cell proliferation, as has been shown in endothelial cells and chondrocytes. 25 –27 In our study, we used cell growth curves to evaluate the function of SIRT6 on BM-MSC proliferation. After knocking down SIRT6 expression using SIRT6 siRNA, hBM-MSC growth was significantly impaired. These data suggest that SIRT6 is important for cellular proliferative capacity and, most importantly, overexpression of SIRT6 in aged stem cells is likely maintaining their growth.
Second, cell migration is an ability of stem cell homing as the repairing potential of stem cells depends on its homing capacity. While a few studies have reported the effect of SIRT6 on cell migration in cancer and inflammatory cells, 28,29 no study has illustrated the effect of SIRT6 on stem cell migration function. We demonstrated that knockdown of SIRT6 impaired the migration ability of hBM-MSCs. Therefore, SIRT6 may serve as a target to improve the homing capacity of stem cells.
Third, the sensitivity of stem cells to apoptotic markers is an important factor for tissue repair efficacy. Van Meter et al. reported that SIRT6 accelerates apoptosis in cancer cells but not in nonmalignant cells. 30 Sundaresan et al. showed that the expression of apoptotic markers increased in SIRT6 knockout hearts. 31 However, Kawahara et al. showed elevated apoptotic resistance in SIRT6 knockdown human primary keratinocytes. 32 In the current study, we showed no significant effect of SIRT6 knockdown on hBM-MSC apoptosis. The mechanism by which SIRT6 regulates cellular apoptosis remains unknown. Further research is needed to understand the inconsistent findings regarding the effect of SIRT6 on cellular apoptosis.
Fourth, knocking down SIRT6 expression increased cell death in cultured hBM-MSCs after exposure to oxidative stress. Reactive oxygen species result in DNA strand breaks, base lesions, genomic instability, and cell death. SIRT6 was reported to be involved in DNA damage repair and genomic stability. 8,14,27,33,34 Our observation of SIRT6 protecting hBM-MSCs from oxidative stress-induced cell death might be due to a DNA repair mechanism.
Fifth, it has been reported that SIRT6 could protect cells from senescent processes. 25,35 Our results are in agreement with previous findings as we detected significantly increased SA-β-Gal activity, a characteristic feature of cell senescence, and significantly increased aging-associated p16 expression in SIRT6 knockdown hBM-MSCs compared with controls. In other words, SIRT6 downregulates cell senescence.
There are many mechanisms of cell aging, including telomere shortening, DNA damage and mutations, genomic instability, and increased reactive oxygen species. 36,37 Cellular aging can be described as the loss of cell proliferative capacity due to telomere shortening. This occurs when the DNA–protein cap structure that protects the linear chromosomes shorten with each cell division and diminish when the replicative life span of cells is finished. SIRT6 acts to maintain the telomeric chromatin, repair DNA damage, and support genomic stability. Hence, SIRT6 may be an important antiaging target for stem cells.
We investigated the proliferative and migration abilities as well as the apoptosis and survival rates of aged hBM-MSCs and compared them with young cells. These factors have a major impact on the repairing ability of stem cells. It is well known that the functional capacity of stem cells declines with age, resulting in a progressive decrease in homeostatic and regenerative capacity. 11 –13,36 Our study showed that cell growth, proliferative and migration abilities, as well as survival rate decreased in aged hBM-MSCs, while the rate of apoptosis increased. These results support the view that aging is an important factor in suppressing stem cell functional capacities. Therefore, rejuvenation of senescent BM-MSCs in aged individuals is urgently needed for stem cell therapy.
In conclusion, we confirmed that aging impairs the proliferative and migration abilities of stem cells while reducing cell survival and increasing cell senescence via enhanced SA-β-Gal activity. Knockdown of SIRT6 also impaired the proliferative, migration, and oxidative stress resistance potentials of hBM-MSCs while increasing SA-β-Gal activity and aging-associated p16 expression. Our results suggest that SIRT6 regulates the aging process, which could serve as a target for the rejuvenation of aged stem cells.
Footnotes
Acknowledgments
We thank Dr. Leigh Botly for help with article preparation and editing. This study was funded by the 331 Early Career Researcher Grant of Shanxi Medical University (No. 201409) and a Canadian Foundation for Innovation Grant (No. RMF111623). R.-K.L. holds a Canada Research Chair in Cardiac Regeneration.
Author Disclosure Statement
No competing financial interests exist.