
Abstract
The quantification of aspartic acid racemization in the proteins of nonmetabolically active tissues can be used as a measure of chronological aging in humans and other long-lived organisms. However, very few studies have been conducted in shorter-lived animals such as rodents, which are increasingly used as genetic and metabolic models of aging. An initial study had reported significant changes in the ratio of
Introduction
T
However, very little work has been done to optimize these racemization dating methods for smaller shorter-lived animals, such as rats. Only two studies have been done within rodents investigating the correlation of aspartic acid racemization in teeth with the age of the animal. In 1995, Ohtani et al. reported significant differences between
Human eye lens has also been analyzed for the accumulation of racemized aspartate derivatives. Like the mineralized collagens and associated proteins in teeth, the long-lived proteins of the eye lens, including α-, β- and γ-crystallin, can accumulate damaged residues. Masters et al. (1977) reported significant racemization rates within the human eye lens, with a linear correlation between age and the
The studies described above demonstrate the time-dependent accumulation of aspartic acid derivatives and its correlation with age, particularly in long-lived species. With the prevalent use of mice and rats in aging and disease research, the development and optimization of methods used to investigate aging markers in these organisms are important. The goal of the work reported here was to explore the use of two age-related markers of aging in rats, racemization in molars and isomerization in the lens. Our results show that significant increases in racemization of aspartate/asparagine residues with age cannot be detected in molars, but that increases in the isomerization of these residues with age can be readily quantified in the lens.
Experimental Procedures
Collection of eye lens and teeth from aged rats
Sprague-Dawley (SD) rats were raised in the INIBIOLP rat colony at the National University of La Plata, Argentina. Animals were housed in a temperature-controlled room (22°C ± 2°C) on a 12-hour light/12-hour dark cycle. Food and water are available ad libitum. In this rat colony, the average 50% survival time for females, studied in groups of 50–60 animals, is approximately 32 months, provided mammary tumors are systematically removed when detected by palpation. The 2-month-old, 18-month-old, and some of the 29-month-old animals received no treatment. However, some of the 29-month-old animals received, 3 months before sacrifice, an introcerebroventricular injection of a placebo adenovector expressing the reporter gene for the DsRed2 red fluorescent protein and were fixed as described below. Assessment of spatial memory by the Barnes Maze test 27 as well as immunohistochemical analysis of glial fibrillary acidic protein (for astrocytes) and doublecortin immunoreactive cells in the hippocampus (for neuroblasts) revealed no significant differences between these placebo control adenovector-treated and untreated animals. Furthermore, the incidence of cataracts in both groups at the time of sacrifice was comparable (unpublished data). Thus, it is unlikely that the placebo adenovector treatment had a significant effect on the rate of eye lens or tooth protein racemization and isomerization. One rat (44 months old, designated SO) was intravenously injected with human bone marrow-derived mesenchymal stem cells as described. 28 Although the mesenchymal stem cell treatment seems to have extended the life span of the 44-month-old rat, it did not prevent the development of strong bilateral cataracts, 28 which again makes it improbable that the treatment had a significant impact on eye lens and tooth protein metabolism.
All animals used here were virgin females. Animals were sacrificed by rapid decapitation, except for the group of 29-month-old animals that were fixed. These animals were anesthetized with ketamine hydrochloride (70 mg/kg, i.p.) and xylazine (15 mg/kg, i.m.) and perfused with phosphate-buffered 4% paraformaldehyde (pH 7.4). After removal of the brain, the heads were frozen at −80°C and shipped to UCLA packed in dry ice. All experiments with animals were performed in accordance with the Animal Welfare Guidelines of NIH (INIBIOLP's Animal Welfare Assurance No. A5647-01); IACUC approved on April 3, 2007, extended December 19, 2011.
At UCLA, heads were thawed at 4°C for 3 days in 300 mL of deionized water. The jaws were separated and molars were removed using a burlisher. Excess soft tissue was removed and the molars were then cleaned in an ultrasonic water bath for 20 min in deionized water, followed by 20 min in ethanol, and vacuum-dried. Eyes were excised using surgical scissors and lenses were extracted under a Zeiss Stemi 2000 light microscope. Lenses were washed gently in 500 μL of deionized water in 1.5-mL microcentrifuge tubes on a rotator for 20 minutes at 4°C. Rat heads for lens extraction were not subjected to any tissue fixation.
Determination of d/l -aspartate ratios in molar hydrolysates
After drying, intact individual molars were acid hydrolyzed in 500 μL of 6 N HCl at 110°C for 6 hours under vacuum in a Waters PicoTag apparatus. Samples were vacuum-dried, resuspended in 50 μL of deionized water, and any remaining particulate material was removed by centrifugation. Analysis of o-phthalaldehyde/N-acetyl-
Determination of l -isoaspartate levels in eye lens proteins
Lenses from unfixed rat heads were thawed as described above, weighed, and then 20 μL of T-PER Total Protein Extraction Reagent (Life Technologies 78510) was added per mg wet weight of lens at room temperature. Sodium EDTA, pH 7.5, and phenylmethanesulfonyl fluoride dissolved in isopropanol were added to final concentrations of 1 mM as protease inhibitors. Lenses were homogenized at room temperature in a 1.5-mL microcentrifuge tube with 200 strokes of a form-fitting plastic pestle (Kimble-Chase Kontes pellet pestle 7495150000; Fisher Scientific). Samples were centrifuged at 10,000 g for 5 minutes at room temperature to pellet unbroken cells and membrane debris. The supernatant was collected and stored at −20°C before further analyses.
The human recombinant protein
Rat eye lens endogenous protein l -isoaspartyl methyltransferase activity assays
In a final volume of 100 μL, 10 μL of lens extract protein was incubated for 2 hours at 37°C with final concentrations of 100 μM KASA(isoD)LAKY peptide, 125 mM Bis-Tris-HCl, pH 6.4, and 10 μM [3H]AdoMet, as prepared above. The reaction was stopped by adding 10 μL of 2 M sodium hydroxide, and 100 μL of the 110 μL mixture assayed for volatile radioactivity, as described in the section above. Background radioactivity was determined in a control lacking the lens extract and was subtracted from the value obtained in samples containing the lens extracts. Assays were performed in duplicate.
Results and Discussion
d -Aspartate accumulation in teeth cannot be used to determine rat age
Tooth enamel is 96% mineralized and contains only 4% protein. 32 Most of the protein that is present is trapped in the solid matrix of hydroxyapatite and lasts for years with little to no metabolic turnover. Dentin is similarly solid and is approximately 70% mineralized. Given that the entrapped proteins can accumulate spontaneous chemical modifications, enamel and dentin can be useful for age determination through the quantification of protein damage.
To optimize rodent age determination using reverse-phase liquid chromatography, attempts were made to replicate the removal of the dental crowns from the molars as described in Ohtani et al. 14 However, the very small size of the rat teeth and the distribution of the enamel prevented separation in our hands. Personal communication with Ohtani revealed that in fact whole molars were used for their analyses as they also found that the teeth were too small for the separation of enamel and dentin and that the description of the separation was incorrect in their article. Thus, we also analyzed whole molars in this study.
After extraction and acid hydrolysis of the whole molar, OPA-NAC derivatives of amino acids were detected and quantified through RP-HPLC. Representative chromatographs can be seen in Figure 1, with

Determination of the
Respective levels of
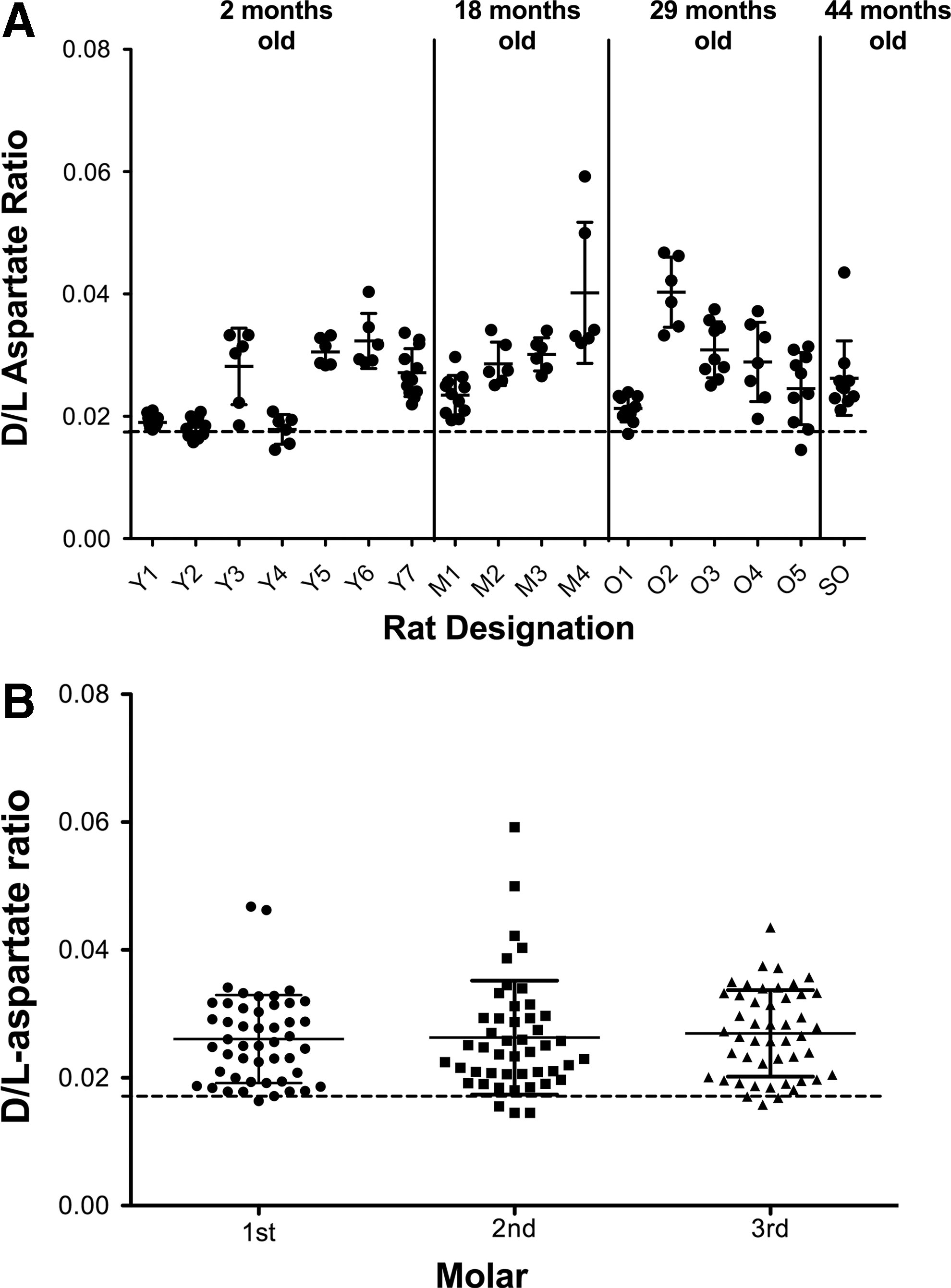
Rat molars are known to develop in a predictable manner, with the front molar erupting at approximately 19 days after birth, the second set on the 21st day, and the last set 35–40 days after birth. 36 Thus, in addition to exploring a relationship to age, we investigated the link between racemization and eruption date. Data for individual teeth were plotted according to the particular molar, shown in Figure 2B, and demonstrated that aspartate racemization does not significantly correspond with molar age relative to other molars in the mouth, contrary to the results reported by Ohtani et al. 14
The discrepancies seen between our results and those in Ohtani et al. 14 may have several origins. Our sampling included multiple rats for each age group excepting the 44-month-old rat, including seven 2-month-old rats, four 18-month-old rats, and five 29-month-old rats. The Ohtani group analyzed five individual rats of different ages ranging from about 2 months to 8 months. The increase in sample size may accurately represent the variability between rats of the same age group. Our age groups also spanned a much longer length of time.
It is also possible that there was some confusion in the Ohtani et al. publication
14
since the techniques reported for the separation of molar enamel and dentin are not possible under the methods given. However, we demonstrate that the variability between rats of the same age prevents this method from reliably identifying rat age or from using
These results indicate that the rate of racemization of rat tooth proteins is slow enough that we cannot distinguish the age-dependent accumulation above the background of spontaneous racemization resulting from the acid hydrolysis procedure. The acid hydrolysis conditions have been optimized to ensure near complete hydrolysis of peptide bonds linked to aspartic acid and asparagine residues while minimizing the background racemization. 23
l -isoaspartate levels in rat eye lens correspond with increased age
The degradation of
We took advantage of the availability of an enzyme that specifically methylates both
Previous reports have indicated that the rat eye lens grows throughout life, but at a declining rate over the lifetime of the organism, 41,42 a situation distinct from that seen in the human eye lens where most of the development occurs before one year of age and only increases very slowly thereafter. 43 In fact, lens wet weight has been used as a marker of rat age. 42,44 –46 From the results seen in Figures 3A and 4A, we also demonstrated an age-dependent increase in eye lens wet weight. However, it seems that as the lens grows, the amount of soluble protein decreases, suggesting the formation of damaged and aggregated proteins, dropping down from about 1000 μg in a 2-month-old rat to just below 500 μg in the 44-month-old rat (Figs. 3B and 4B).

L-isoaspartate accumulates in the rat eye lens after 2 months, but plateaus after 18 months of age. Rat eye lenses were extracted from seven young rats (2 months), four middle-aged rats (18 months), five old rats (29 months), and one exceptionally old rat (44 months), including several of the animals whose teeth were analyzed in Figure 2. Rats were identified as in Figure 2; R and L represent the left and right lenses. Cytosolic extracts were made and L-isoaspartate content measured as described in the Experimental Procedures section.

The data from Figure 3 are plotted as a function of the age of the individual rats. In
Figures 3C, 3D, 4C, and 4D show the quantification of the levels of
The plateau/decrease in total
To investigate this possibility, the specific activity of the endogenous PCMT1 enzyme was assayed in the eye lens extracts using saturating amounts of an
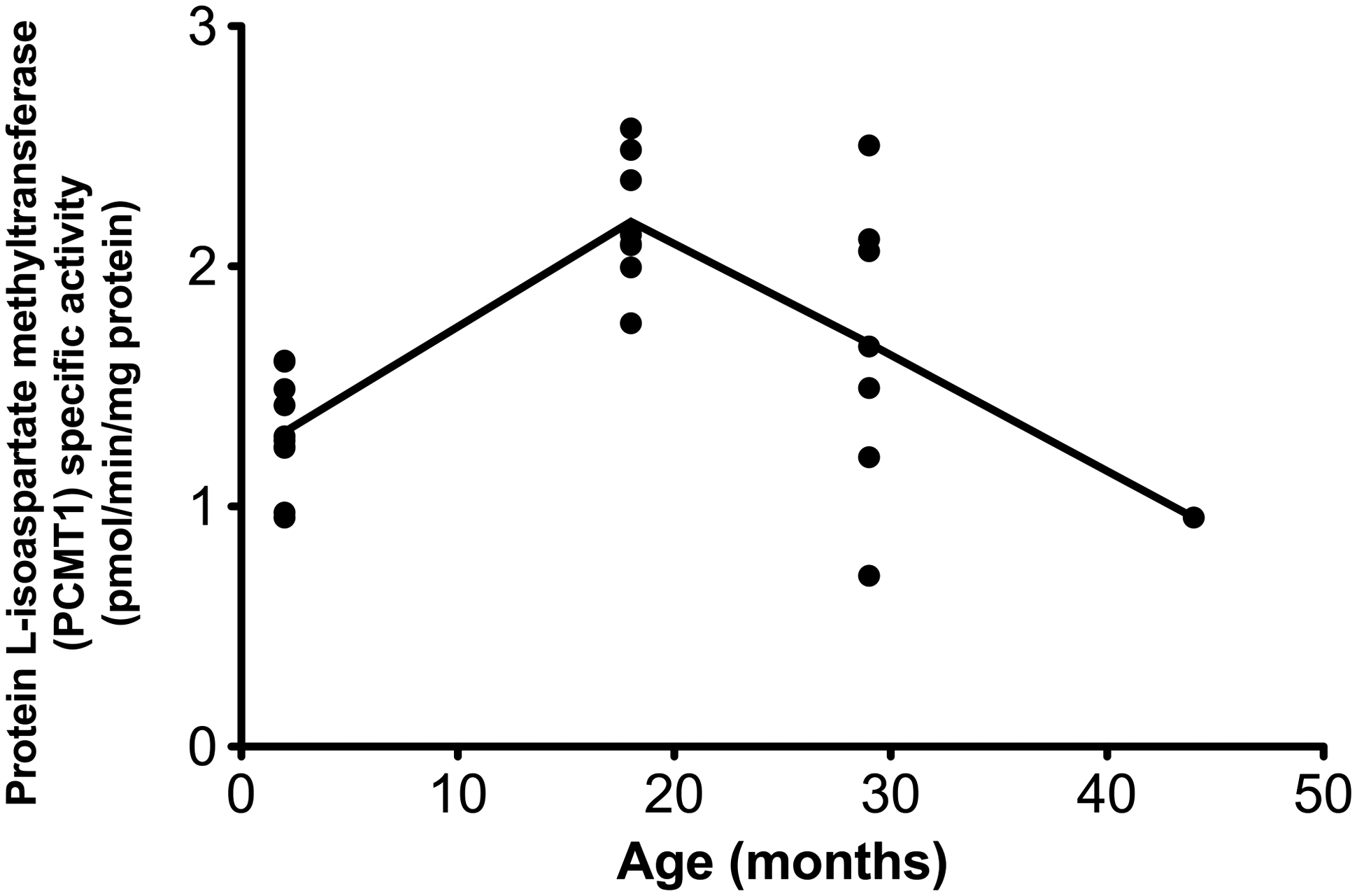
Age dependence of the specific activity of endogenous protein
Since we observed that the levels of total soluble protein decreased over the lifetime of the rats, a second possibility is that the formation of aggregates may prevent the accumulation of
In mice, it was similarly observed that in heart and testis extracts the amount of isoaspartate initially accumulates rapidly, and then levels off, suggesting again possible compensation mechanisms. 40 It is possible that the number of proteins with labile aspartate/asparagine residues is limited and that the plateau is due to all possible sites being racemized or isomerized. However, within an mg of protein, there are approximately 800 nmols of asparagine and aspartate residues, which may racemize or isomerize. The maximum level of damage that was observed in our study was about 12 nmol isoasparate/mg protein (Fig. 3D), suggesting that we are only observing approximately 1.5% of sites forming aberrant derivatives.
As stated previously, whole tooth analysis for
Footnotes
Acknowledgments
The authors thank Dr. Aubrey D. N. J. de Grey of the SENS Research Foundation (Mountain View, CA) for initiating this work and for providing guidance throughout. The authors thank Dr. Shane White of the UCLA School of Dentistry for his help in preparing tooth samples and Drs. Rajendra Gangalum and Suraj Bhat of the UCLA Jules Stein Eye Institute for assistance and advice on the preparation of lens samples. They thank Dr. Jonathan Lowenson for his guidance in this work and for his help in preparing the manuscript. This work was supported by a grant from the Life Extension Foundation, Inc., and funds from the Elizabeth and Thomas Plott Chair in Gerontology of the UCLA Longevity Center. R.A.W. was supported by the Ruth L. Kirschstein National Research Service Award GM007185.
R.G.G. and E.M. provided the animal material; E.M. took responsibility for monitoring the health of the 44-month-old rat; R.G.G., R.A.W., and S.G.C. planned the study; R.A.W. performed the analytical experiments; R.A.W. and S.G.C. wrote the article and analyzed the data; and R.A.W., E.M., R.G.G., and S.G.C. reviewed and revised the article.
Author Disclosure Statement
No competing financial interests exist.