
Abstract
Lysyl oxidase-like 1 (LOXL1) is an amino-oxidase involved in maturation of elastic fibers. Its downregulation has been associated with elastic fibers repair loss in aging aorta, lung, ligament, and skin. Several evidences of LOXL1 epigenetic silencing by promoter methylation were reported in cancer and cutis laxa syndrome. We hypothesized that this mechanism could be implicated in skin aging process, as far as elastic fibers are also concerned. Anti-DNMT3A chromatin immunoprecipitation was conducted with nuclear extracts from skin fibroblasts isolated from young and elderly individuals, and showed a higher level of DNMT3A protein binding to the LOXL1 promoter in older cells concomitantly to the decrease of LOXL1 mRNA expression and the increase of LOXL1 promoter methylation. Using luciferase reporter assay driven by LOXL1 promoter in HEK293 cells, we demonstrated that LOXL1 transcriptional activity was dramatically reduced when a recombinant DNMT3A was concomitantly overexpressed. LOXL1 promoter transcriptional activity was restored in the presence of a broad-spectrum inhibitor of DNMT activity, 5-aza-2′-deoxycytidine. Finally, to assess whether the interplay between DNMT3A and LOXL1 promoter could be targeted to increase LOXL1 mRNA expression level, an Origanum majorana extract was selected among 43 plant extracts as a new inhibitor of human DNMT3A activity to restore LOXL1 secretion without cytotoxicity in aged skin fibroblasts.
Introduction
L
Transgenic mice deficient for LOXL1 have provided evidences that this enzyme is necessary for the maintenance and the renewal of elastic fibers during life, 3 while the LOX isoform is mainly involved in the development of elastic fibers during embryogenesis. 4 In general, strong correlations were made between a defect of LOXL1 expression and various diseases attributed to a lack of elastogenesis such as the pelvic organ prolapse, 5,6 the pseudoexfoliation syndrome, 7 venous insufficiency, 8 aorta remodeling, 9 and the cutis laxa syndrome. 10 Finally, a downregulation of LOXL1 gene is observed during chronological aging in many organs in mice 11 and in human skin, 12,13 and is thus considered an attractive pharmacological target to restore elastogenesis at adulthood. However, little is known about the mechanisms involved in the repression of LOXL1 during aging.
In a previous study, we demonstrated that, in absence of a functional TATA box, the transcription factor specific protein 1 (SP1) was the main inducer to recruit the transcriptional machinery leading to the LOXL1 promoter activation in fibroblasts from healthy young individuals. 14 These SP1 binding motifs are located in a GC-rich proximal region (−323/−494 relative to start codon) prone to DNA methylation. As methylated DNA was shown to counteract SP1 binding to its recognition sequence, 15 we then compared the methylation status of LOXL1 promoter from healthy and cutis laxa cells, and thus assigned DNA methylation as the mechanism responsible for LOXL1 repression in pathologic fibroblasts. Interestingly, a same epigenetic mechanism was reported in bladder cancer cells, 16 and in the pseudoexfoliation syndrome 17 where LOXL1 expression was dramatically decreased. These two studies, both conducted in different pathological contexts, highlight the importance of epigenetic regulation in controlling LOXL1 expression level.
Methylation profiles evolve over time and are dependent on the cell type considered. During skin aging, dermal fibroblasts undergo a global DNA hypomethylation in conjunction with a strong local hypermethylation of CpG sites in specific loci. 18,19 DNA methylation is mainly associated with chromatin silencing and consists in the transfer of a methyl group from the methyl donor S-adenosylmethionine to a cytosine in the CpG dinucleotide of eukaryotes. The transfer is orchestrated by the DNA-methyltransferase (DNMT) family of enzymes comprising three active isoforms: DNMT1, DNMT3A, and DNMT3B. 20
Briefly, on the one hand, DNMT1 is considered as a maintenance enzyme in charge of copying the methylation pattern from the parent to the daughter strand during DNA replication. On the other hand, DNMT3A and DNMT3B are de novo methyltransferases able to methylate hemimethylated or unmethylated DNA. However, DNMT3A and DNMT3B are not redundant enzymes. DNMT3B methylates DNA sequences, primarily minor satellite repeats, during early development, whereas DNMT3A has a more important role in chromatin methylation during late development and after birth. 21
Focusing on human DNMT3A isoforms, two main variants were described as DNMT3A1 and DNMT3A2. DNMT3A1 is ubiquitously expressed at low level and is mostly associated to heterochromatin. Interestingly, the DNMT3A2 variant expression is restricted to several tissues and is associated to euchromatin where single-copy genes, that is, protein coding sequences, are localized. 22 However, its expression in human skin has never been investigated so far.
We hypothesized that age-associated downregulation of LOXL1 could be orchestrated by epigenetic changes. The aims of this study were to evidence DNA methylation-mediated silencing of LOXL1, and to restore its expression by targeting such a mechanism in skin fibroblasts from elderly individuals.
Materials and Methods
Cell culture
Dermal fibroblasts were isolated from skin surgical explants of healthy individuals at different ages: five young females (1, 2.5, 4, 4, and 10 years) and five older females (59, 61, 62, 63, and 68 years). The cells were purchased from the Biological Resource Centre CBC-Biotec (Lyon, France), anonymity of donors was respected. Cells culture medium consisted in a ratio 1:1 of Dulbecco's Modified Eagle's Medium with Glutamax and F-12 medium (Invitrogen, Cergy Pontoise, France) supplemented with 10% fetal calf serum (Sigma-Aldrich, Saint-Quentin-Fallavier, France), 100 IU/mL penicillin, and 0.1 mg/mL streptomycin (Sigma-Aldrich). Cells were used below 20 population doubling level.
Human embryonic kidney (HEK) 293 EBNA cells were purchased from CelluloNet, Biological Resources Centre (UMS3444; Lyon, France). Cells culture medium consisted in Dulbecco's Modified Eagle's Medium with Glutamax (Invitrogen) supplemented with 10% fetal calf serum (Sigma-Aldrich), 100 IU/mL penicillin, and 0.1 mg/mL streptomycin (Sigma-Aldrich).
Origanum majorana extraction
Minced dry Origanum majorana was obtained from the BASF-BCS library. Five grams of solids were mixed with 100 mL of distilled water and placed in maceration at 4°C with stirring overnight. The macerate was centrifuged 15 min/8000 g at 4°C. The supernatant was then filtered sequentially 3, 1.2, 0.8, 0.45, and 0.22 μm until a clear aqueous solution was obtained and stored at −20°C. The extract was thawed extemporaneously and diluted at 1% (v/v) in the culture medium.
Real-time reverse transcription-polymerase chain reaction
Total RNA was extracted using RNeasy kit (Qiagen, Courtaboeuf, France). One microgram of total RNA was reverse transcribed using RevertAid H-Minus First Strand cDNA Synthesis kit (Fermentas, St. Rémy Les Chevreuse, France). Real-time PCR was performed on a cDNA mixture, equivalent to 25 ng of total RNA, with FastStart Universal SYBRGreen Master (Roche, Meylan, France) and Rotor-Gene-Q thermocycler (Qiagen). The primers were specific for human GAPDH (Forward, acacccactcctccaccttt; Reverse, agccaaattcgttgtcataccag), LOXL1 (Forward, gcacctctcatacccagggc; Reverse, tggcagtcgatgtccgcat) and DNMT3A (Forward, catctgcatctcctgtggg; Reverse, gcagttttggcacattcctc). Data analyses were performed using the comparative Ct method.
Chromatin immunoprecipitation
Cells plated in 175 cm2 flasks were fixed with 1% formaldehyde for 10 minutes, neutralized with glycine for 5 minutes to a final concentration of 125 mM, rinsed twice with Tris-buffered saline (TBS) (100 mM NaCl, 50 mM Tris-HCl pH 8.1), and then scraped and syringed with TBS containing 0.5% w/v SDS. The cell pellet was resuspended in the same buffer completed with 2% v/v Triton X-100.
The cell pellet was sonicated to yield DNA fragments of ∼1000 bp in average, immunoprecipitated with 2 μg of an anti-DNMT3A monoclonal antibody (MAB0080, Abnova; Tebu-bio, Le Perray en Yvelines, France), or 2 μg of normal IgG as negative control. Unprecipitated chromatin was used as positive control (input) and conserved at 4°C until DNA isolation step. For the two other tubes, after overnight incubation at 4°C with continuous mixing, aggregates were removed by centrifugation and protein-A beads were added to supernatant.
After 2 hours at 4°C, immune complexes were collected by centrifugation at 4°C and washed three times with wash buffer containing 150 mM NaCl, 20 mM Tris (pH 8.1), 5 mM EDTA (pH 8), 6% w/v sucrose, 0.2% NaN3, 1% v/v Triton X-100, 0.2% w/v SDS, and twice with solution containing 1 mM EDTA, 50 mM N-[2-hydroxyethyl]piperazine-N′-[2-ethanesulphonic acid], pH 7.5, 500 mM NaCl, 1% v/v triton X-100, 0.2% w/v NaN3, 0.1% w/v deoxycholic acid, and twice with LiCl detergent solution (10 mM Tris pH 8, 1 mM EDTA pH 8, 250 mM LiCl, 0.2% w/v NaN3, 0.5% v/v Nonidet P-40, and 0.5% w/v deoxycholic acid), and finally in Tris–EDTA. All washes were performed for 5 minutes at 4°C with mixing.
To eluate immune complexes from beads, incubation was carried out for 45 minutes at 65°C in reversal buffer (1% w/v SDS and 100 mM NaHCO3). Eluate was then incubated overnight at 65°C in the same buffer to reverse cross-links. DNA was purified with phenol–chloroform and precipitated with ethanol using glycogen as a carrier. The purified DNA isolated by immunoprecipitation was analyzed by PCR using primers specific for the proximal LOXL1 gene promoter (Forward, tcctcagaggcaggtctgtg; Reverse, ataagcaacaggctgggagc). PCR products were resolved by a 2% agarose gel with ethidium bromide and optical density of bands was analyzed with Image J software.
DNMT3A-flag expression vector cloning
cDNA (NM_153759.2) coding for DNMT3A was purchased from Origene (ref: SC109167; CliniSciences, Nanterre, France). cDNA was amplified with primers containing NheI and SalI sites in 5′ and 3′ positions respectively. PCR was performed using the high fidelity Authentik DNA polymerase (Eurogentec, Angers, France). The 3′ primer also contained the sequence coding for the Flag motif (DYKDDDDK). PCR product was digested by NheI and SalI (Promega, Charbonnières, France), in parallel, the pCep4 vector (Invitrogen) was digested with NheI and XhoI (Promega). Then, insert was ligated into pCep4 plasmid to form the pCep4-DNMT3A-Flag expression vector.
Luciferase assays
HEK 293 EBNA cells were plated in 24-well plates at 200 × 103 cells/well. The day after, subconfluent cells have been transfected with 1 μg total DNA and 3.5 μg Lipofectamine 2000 (Invitrogen). DNA was a mix of three different plasmids that have been co-transfected. pRL-TK plasmid was used to normalize transfection efficiency. PrLOXL1 constructions cloned into basic pGL3 plasmid (Promega) are as follows (see Debret et al. 14 for details): pLL28 (−2172 bp), pLL32 (−712 bp), and pLL34 (−81 bp). Promoter constructions were co-transfected with pCep4 (Mock) or pCep4-DNMT3A-Flag (DNMT3A) expression vectors. DNMT activity was inhibited using 5-aza-2′-deoxycytidine (Sigma-Aldrich) at 5 μM or with an O. majorana exract (patent n°FR2999926, BASF—Beauty Care Solutions, Lyon, France) at 1% (v/v) for 24 hours.
Transfected cells were lysed with passive lysis buffer (Promega), and then Renilla luciferase (480 nm) and Firefly luciferase (560 nm) activities were assessed using Dual-Luciferase Reporter Assay System (Promega). Results were normalized with pRL-TK as transfection efficiency reference and with pGL3 basic as background reference.
Western blotting
For DNMT3A-Flag immunodetection, 20 μg of protein extract were separated by 10% SDS-PAGE. Electrophoretically resolved bands were then transferred to polyvinylidene difluoride membranes (Immobilon; Millipore SAS, Molsheim, France) by electroblotting. The blots were incubated with a monoclonal antibody raised against Flag (ref: F3165; Sigma-Aldrich) in a dilution of 1:5000. Blots were then incubated with peroxidase conjugated anti-mouse IgG (Bio-Rad) before detection using an enhanced chemiluminescent kit (Perkin Elmer, Courtaboeuf, France).
Methyl-sensitive high resolution melting PCR
LOXL1 promoter methylation status was assessed in the different cell strains or in 62 years old fibroblasts treated or not with 1% O. majorana and gDNA was extracted using QIAamp DNA mini kit (Qiagen).
One microgram of gDNA was subjected to sodium bisulfite conversion using EpiTect Plus DNA Bisulfite kit (Qiagen). Converted DNA concentration was determined with Nanodrop spectrophotometer and diluted with water at 10 ng/μL. HRM-PCR run were performed in 25 μL mix containing 10 ng of single strand converted-gDNA, 12.5 μL EpiTect HRM PCR mix (Qiagen), 1.9 μL of 10 μM primers (Forward, gttygatttttttttagaggtag; Reverse, actttatcctatctaacctt), and water. Rotor-Gene-Q thermocycler (Qiagen) program was as follows: 95°C 5 minutes, then 45–50 repeated cycles with 95°C 10 seconds denaturation, 60°C 30 seconds hybridization, 72°C 32 seconds elongation, and a melt curve was performed from 75°C to 85°C with a 0.1°C step.
Immunofluorescence and microscopy
For immunofluorescence, cells were cultured at postconfluence on labteks and then fixed in cold (−20°C) methanol for 10 minutes, washed three times in PBS, and preincubated 60 minutes at room temperature with BSA 1% in PBS. Then, cells were incubated 1 hour at room temperature with a rabbit polyclonal anti-LOXL1 antibody diluted at 1:500 in PBS-BSA 1%, followed by three washes with PBS and by a second incubation with donkey anti-rabbit alexa 488 (Life Technologies) 1:1000 in PBS-BSA 1% at room temperature for 40 minutes. After three washes in PBS, Labteks were mounted with ProLong reagent (Life Technologies) containing 4′,6-diamidino-2-phenylindole (DAPI) for nuclear stain. Immunofluorescence pictures were acquired with a Zeiss LSM 700 confocal microscope.
Statistical analyses
The results are expressed as mean values ± standard error of the mean from triplicate assays or independent experiments as indicated in figure legends. The most appropriate test for comparison of data was selected from unpaired-Student's t test, paired-Student's t test, and one-way ANOVA test.
Results
DNMT3A binding on LOXL1 promoter in skin fibroblasts
Experiments were carried out using normal skin fibroblasts from healthy female donors divided in two groups of different ages: five strains below 10 years (“young”) and five strains over 59 years (“elderly”). Cells were cultured for 2 days after confluence, and then total RNA was extracted. Real-time reverse transcription-polymerase chain reaction (RT-PCR) assessments with primer specific for LOXL1 mRNA showed a significant lower level of expression in older cells when compared with young cells (Fig. 1a). This drop in LOXL1 expression validates our cellular model.
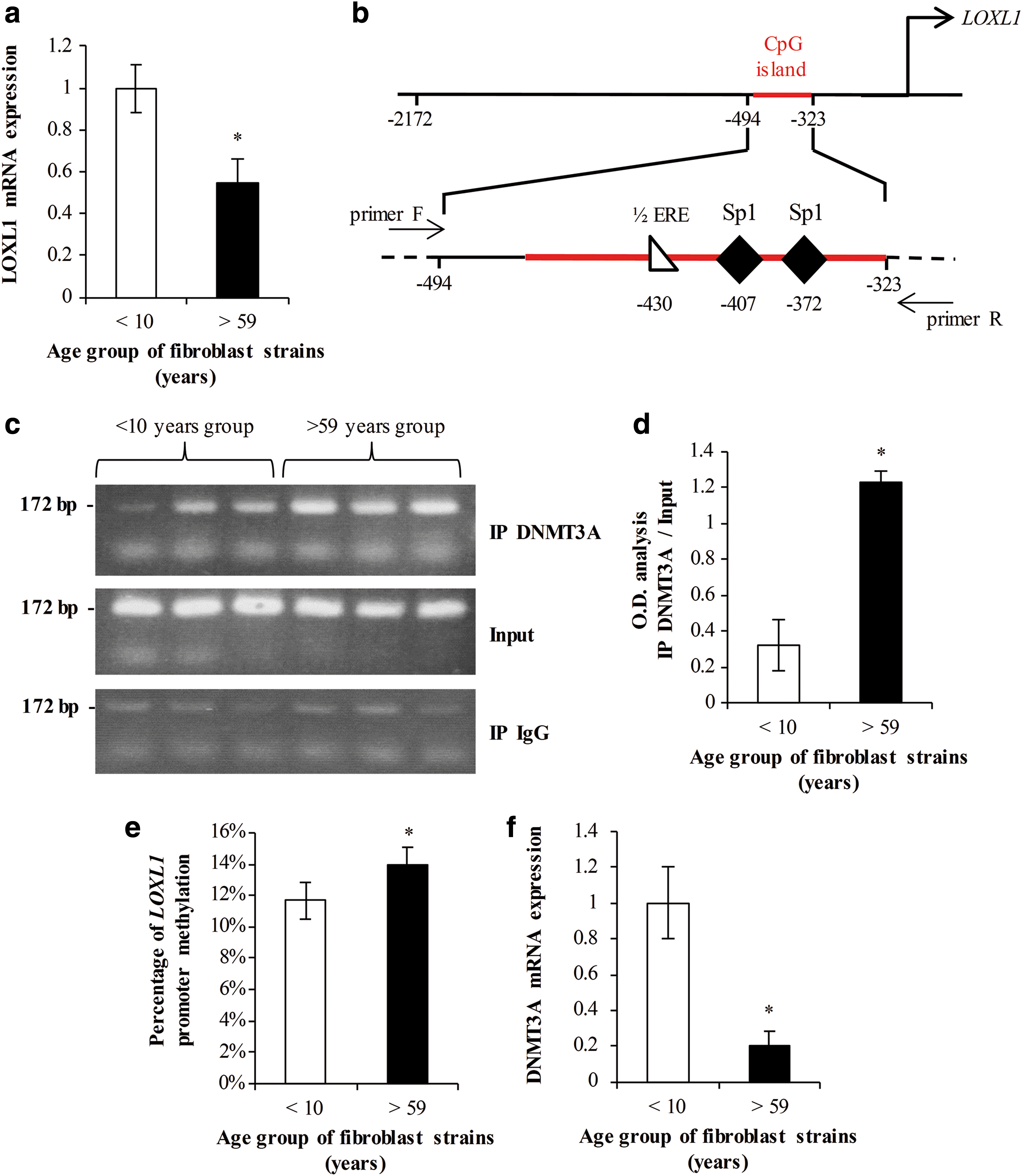
DNMT3A binding and DNA methylation are increased in LOXL1 promoter from “elderly” fibroblasts.
The direct link between DNMT3A and LOXL1 promoter was then assessed by chromatin immunoprecipitation in 3 cell strains of each group. After precipitation of the chromatin with an antibody raised against the DNMT3A protein, PCR were conducted with primers specific for the GC-rich proximal region of the LOXL1 promoter (Fig. 1b). An aliquot of preimmunoprecipitated chromatin was used as control (“input”). PCR amplicons resolved by agarose-ethidium bromide gels are shown in Figure 1c. The bands with higher intensities indicate a higher binding of DNMT3A within LOXL1 promoter. Optical density analysis of the bands showed a significant increase in the older group (Fig. 1d). This experiment clearly demonstrated that DNMT3A isoform can bind the promoter of LOXL1 gene. Moreover, this binding is observed at a higher extent in older cells.
The methylation status of LOXL1 promoter in these cell strains was then estimated by methyl sensitive high resolution melting (MS-HRM) PCR with primers specific for the proximal region after conversion of the gDNA by bisulfite treatment as detailed in the Materials and Methods section. Using this method, a slight (3%) but significant increase of LOXL1 promoter methylation was observed in cells from older individuals (Fig. 1e).
The local increase of DNMT3A, and its subsequent effect upon LOXL1 promoter methylation, possibly arises from an overall overexpression of this isoform in the cell. DNMT3A gene expression was thus followed in all cell lines by real-time PCR (Fig. 1f). The result shows a fivefold decrease in expression of the DNMT3A gene in the older group. This reinforces the hypothesis of a DNMT activity targeting the promoter region of LOXL1 in these cells. Taken together, these results showed that the drop of LOXL1 expression level in skin fibroblasts isolated from older subjects was accompanied by a local increase of DNMT3A binding correlated to an increase of DNA methylation within the LOXL1 proximal promoter region.
Effect of DNMT3A-overexpression on LOXL1 promoter activity in HEK293 cells
To further assess the direct effect of DNMT3A on LOXL1 expression, we analyzed LOXL1 promoter activity by measuring relative activity of luciferase reporter gene fused to different constructs of the LOXL1 promoter (Fig. 2a): long promoter pLL28 (−2172 bp relative to ATG), minimal promoter pLL32 (−712 bp relative to ATG), and null promoter pPLL34 lacking the GC-rich region (−81 bp relative to ATG).

DNMT3A inhibits LOXL1 promoter activity.
Each promoter construction was co-transfected in Human Embryonic Kidney 293 EBNA cells (HEK 293 EBNA) with empty pcep4 plasmid (Mock) or with pcep-DNMT3A plasmid (DNMT3A) coding for DNMT3A. DNMT3A cDNA (NM_153759.2) was cloned into pCep4 plasmid with Flag tag in C-terminus. pGL3 basic vector was used as background control and co-transfections have been completed with pRL-TK plasmid as transfection efficiency control. Luciferase activities were analyzed 24h post-transfection (Fig. 2b). In mock conditions (white bars), pLL32 activity was higher than pLL28 activity, while pLL34 activity was almost not detected as already described. 14
In the presence of the coding vector pcep-DNMT3A (black bars), pLL28 and pLL32 luciferase activities were dramatically decreased while no significant change was observed with pLL34 promoter. Correct expression of the DNMT3A protein has been assessed by anti-Flag western blotting for each condition (Fig. 2c). A decrease in luciferase activity indicated that pLL28 and pLL32 promoter constructions were transcriptionally repressed in presence of DNMT3A protein. Absence of effect on pLL34 construction indicated that DNMT3A affects transcription with regards to the promoter region but not the luciferase coding sequence. To further investigate whether LOXL1 promoter repression was dependent of DNMT3A enzymatic activity, same experiments were conducted using 5-aza-2′-deoxycitidin (5azadC) as broad-spectrum inhibitor of DNMT activity (Fig. 2b, right panel).
In presence of 5azadC, no change in luciferase activity was observed between mock and pcep-DNMT3A conditions. In conclusion, the LOXL1 promoter transcriptional repression was assigned to DNMT3A enzymatic activity in this model.
Selection of O. majorana as potential DNMT3A inhibitor
Because 5azadC is highly cytotoxic and not compatible with long periods of culture, we screened a bank of plant extracts using the co-transfected HEK293 model to select a noncytotoxic active herbal extract capable of limiting LOXL1 promoter repression by DNMT3A. About 43 plant extracts were tested and the result obtained for an O. majorana extract is shown (Fig. 3a). The luciferase activity driven by the promoter pLL32 was not affected in presence of 1% O. majorana (Mock condition). When DNMT3A was co-transfected, the inhibition of luciferase activity remained significant but less strong in the presence of the plant extract compared to untreated conditions (hash). This illustrates a partial inhibition of DNMT3A activity by O. majorana. Although the inhibition of DNMT3A activity appeared low after 24 hours, the potential use of this plant extract over several days in culture made us consider this effect as efficient enough to be tested as a LOXL1 inducer in a normal cell strain where DNMT3A is not overexpressed.
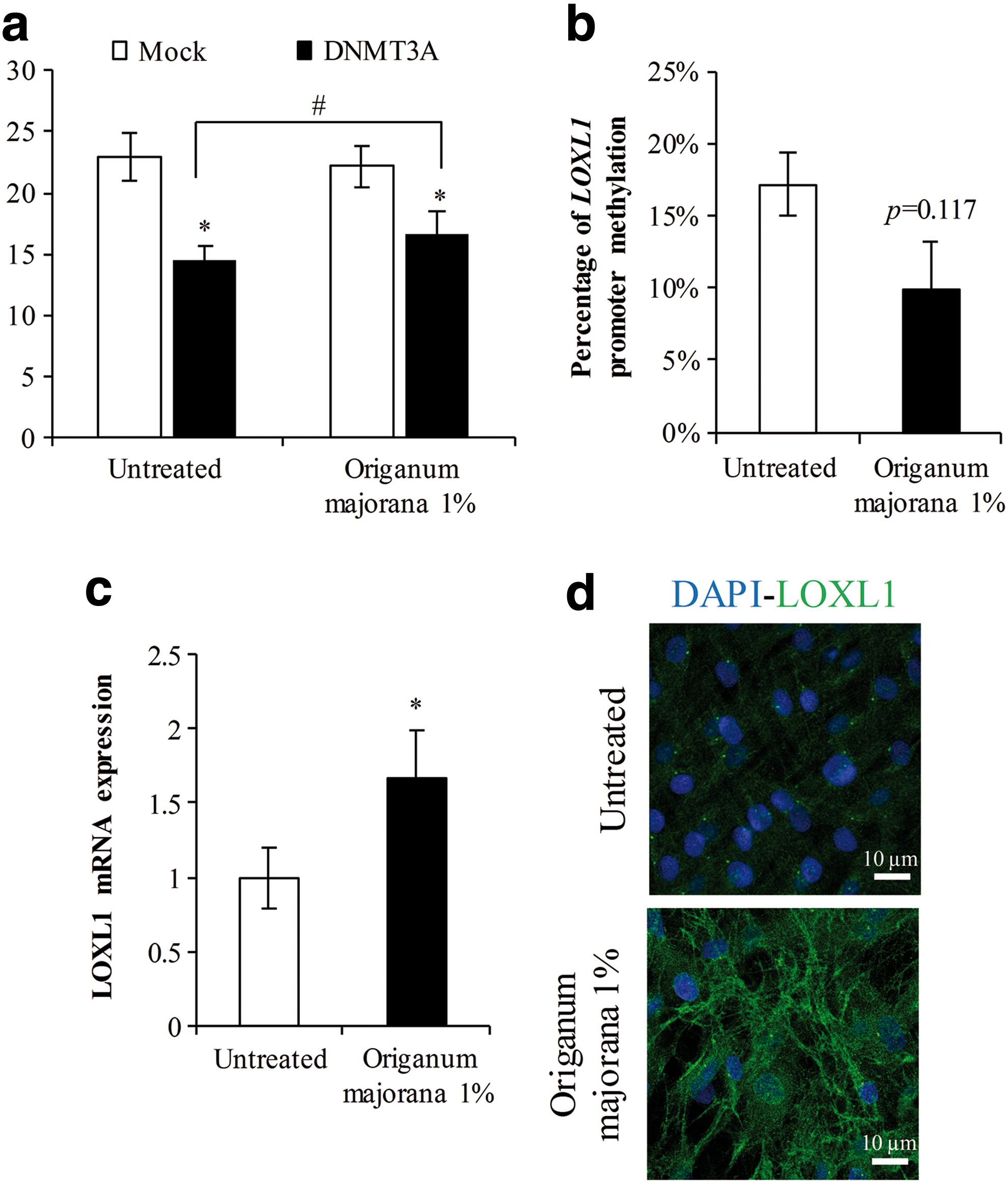
Origanum majorana extract decreases LOXL1 promoter methylation and increases LOXL1 mRNA expression and protein deposition in “elderly” fibroblasts.
O. majorana effect on LOXL1 expression in an elderly cell strain
To validate O. majorana extract as LOXL1 inducer, skin fibroblasts from a 62 years individual were used. Cells were seeded at 80% of confluence and medium containing or not the plant extract was refreshed every 2 days. Total RNA and gDNA were extracted 24 hours after confluence. The methylation status of LOXL1 promoter was estimated by MS-HRM-PCR, and LOXL1 expression was assessed by real-time RT-PCR. We observed a significant decrease in the melting temperature of amplified converted gDNA from cells treated with 1% O. majorana (Fig. 3b). This decrease indicated that methylation marks were present at a lesser extend within the LOXL1 promoter from treated cells. Moreover, this was correlated with a higher LOXL1 expression level compared to untreated conditions (Fig. 3c).
Deposition of LOXL1 protein in the extracellular compartment was then investigated by immunofluorescent staining and confocal microscopy in cultures at 8 days postconfluence (Fig. 3d). In untreated cells, LOXL1 was sparsely detected within the cytoplasm whereas LOXL1 deposition appeared more intense and organized in fibrillar structures in presence of O. majorana leaf extract. This observation demonstrated a higher level of LOXL1 production consistent with the above upregulation of LOXL1 gene under treatment. Taken as a whole, these last results demonstrated that the O. majorana leaf extract was effective in restoring LOXL1 expression and production in long period of culture of an elderly cell strain by counteracting methylation within the LOXL1 promoter.
Discussion
In the first part of this study, we investigated whether age-related LOXL1 downregulation could be attributed to a DNMT-dependent epigenetic remodeling as previously observed in a cutis laxa case. 14
We conducted our experimentations in fibroblasts from “young” versus “elderly” individuals at early passages. Because postconfluent cultures are required for LOXL1 maturation during early elastic fibers assembly process, 23 LOXL1 mRNA expression was investigated 2 days after confluence. In agreement with previous studies, 9,12,13 we observed a downregulation of LOXL1 mRNA expression level in postconfluent “elderly” fibroblast cultures thus validating our research model and the selected cell strains.
In this context, we assigned the DNMT3A as a potential negative regulator of LOXL1 promoter activity with age since its binding on LOXL1 promoter was increased in “elderly” cells. However, we also show that the expression of DNMT3A is greatly reduced in these cells. This is in line with the accepted concept that aging is associated with an overall decrease in DNA methylation in conjunction with the local hypermethylation of certain genes, in particular in their promoter region. Specifically, with regard to DNMT3A, it turns out that its expression entirely follows this scheme through various tissues from the central nervous system to the liver. 24,25
At the global skin level, the expression of DNMT3A has been shown to be greatly reduced with age, 19 in similar proportions to the result we obtained in cultured skin fibroblasts. In this study, the authors also compiled a list of hypermethylated genes including LOXL1. The exact mechanism to explain how DNMTs jointly lose expression and gain in specificity remains unclear. However, an assumption has been proposed by Jung and Pfeifer, 26 pointing out some unmethylated DNA regions initially protected by the polycomb repressor complex 1 (or 2), which undergoes degradation with age to leave access to de novo DNMTs to locally produce a new pattern of hypermethylated regions. Hence, counteracting this mechanism appears mandatory for longevity.
We then screened a bank of noncytotoxic plant extracts, and their ability to counteract LOXL1 promoter inactivation by DNMT3A was assessed. An O. majorana leaf extract was selected for its ability to restore partially the LOXL1 promoter activity in the presence of recombinant DNMT3A. The last decades have seen the emergence of many DNMTs inhibitors primarily used in the fields of carcinogenesis.
Among these compounds, two derivatives of the nucleoside cytidine, azacytidine, and 5azadC, have been extensively studied and used in phase-II clinical trials as chemotherapeutic agents. 27 More recently, a phase-III clinical trial was conducted with orol azacytidine in the treatment of acute myeloid leukemia. 28 5-azadC displays strong cytotoxicity mediated by the de novo DNMT3A and DNMT3B isoforms in embryonic stem cells, 29 and induces premature senescence of fetal lung fibroblasts. 30 The use of cytotoxic compounds is thus not compatible with long-term cultures or treatments required to restore extracellular matrix (ECM) homeostasis in aging.
The O. majorana extract is an interesting alternative since the inhibition of DNMT3A was moderate and did not display any toxicity toward dermal fibroblasts survival in our hands. Several studies reported beneficial effects of this plant extract, used as aqueous extract or essential oil, to protect from gastric mucosal injury, 31 to inhibit advanced glycation end-products formation in rats, 32 and to induce apoptosis in a leukemia cell line. 33 No cellular mechanism has been put forward; however, it is tempting to speculate that the high antioxidant content of the O. majorana could mediate most of these effects. 34,35
Although we failed in determining a direct inhibition of DNMT3A by O. majorana due to an incompatibility of our extract with in vitro DNMT assays, we assume that an indirect mechanism involving antioxidant molecules would be more convincing. This hypothesis is supported by recent investigations showing the role of diet polyphenols in cancer chemoprevention by targeting the epigenetic machinery. 36
Finally, our results showed an increase of LOXL1 expression and deposition in the extracellular space, in correlation with a decrease of LOXL1 promoter methylation in “elderly” fibroblasts in response to 1% O. majorana treatments. These results demonstrate that a gene encoding for a protein from the ECM can be inactivated by epigenetic regulation during aging. Hypermethylation of several ECM-related genes was observed in correlation with age in different organs such as COL1A1 in the periodontal ligament, 37 RELN in the neocortical region, 38 and SPARC in intervertebral discs. 39 Although gene regulation through epigenetic mechanisms is extensively documented regarding cancer and oncogenes, very little is known about such a regulation of genes encoding proteins of the ECM. A better understanding of these mechanisms paves new ways to counteract ECM defects in aging and fibrotic diseases.
In conclusion, we demonstrated that LOXL1 can be silenced in aging fibroblasts by a DNMT-dependent epigenetic mechanism that could be reversed by the use of an O. majorana extract. This work offers new therapeutic approach and contributes to better understand the mechanisms involved in connective tissues repair loss during aging and age-related diseases.
Footnotes
Acknowledgments
This work was supported by the CNRS—French National Centre for Scientific Research, the Claude Bernard University Lyon 1 and the SRD—French Society of Dermatological Research. Léa Moulin was a recipient of a studentship from the CNRS and BASF–BCS France SAS partnership.
Author Disclosure Statement
No competing financial interests exist.