
Abstract
Background/Aims:
The main purpose of the present study was to investigate the effects of vitamin C and E supplements on the antioxidant capacity of erythrocytes obtained from young and aged rats.
Methods:
Male Wistar rats aged 3 and 24 months were used. Vitamins C and E were injected at doses of 200 mg/kg (day) intraperitoneally in young and aged groups. The antioxidant capacity, oxidant stress parameters, and deformability of red blood cells collected from different age stages were evaluated. An in vitro oxidation system was constructed to explore the mechanisms of antioxidant capacity change in the vitamin treatment groups.
Results:
Treatment with vitamins C and E can effectively restore the antioxidant capacity and deformability of red blood cells (RBCs) in aged rats. Under in vitro oxidative conditions, an age-dependent decline in the influx rate of L-cysteine was observed. This was significantly improved following treatment with vitamins C and E.
Conclusion:
We present evidence of an improvement in the antioxidant capacity of RBCs by treatment with vitamins C and E in aged rats. These observations also suggest that treatment with vitamins C and E improves glutathione synthesis by enhancing the influx rate of L-cysteine through the modification of membrane proteins and lipids.
Introduction
A
The life span of human mature red blood cells (RBCs) is 120 days, whereas life span of rat erythrocytes was reported to be only 60 days. As the major component of the circulatory system, erythrocytes not only transport gas but also play a vital role in maintaining the balance of osmotic pressure and redox in vivo. 3 –5 Aging is characterized with disruption of balance between oxidants and antioxidants due to depletion of antioxidants and excess accumulation of ROS, leading to cellular dysfunction. 6,7 The RBC membrane consists of a lipid layer with phospholipids and cholesterol, the biological and mechanical properties of which were severely impaired during aging due to oxidative stress. Current studies suggest that aging may contribute to impaired erythrocyte insulin signaling, 8 reduced total antioxidant status, increased ROS production, and markedly decreased RBC membrane fluidity. 9 In addition, aging is also accompanied with changes in the plasma membrane redox system (PMRS) and decreases in the mechanical properties of RBCs. 10
Since the aging process is characterized with oxidative damage, antioxidant supplements might be practicable in protecting cells from oxidative injury. Among the potential compounds in foods, vitamins C and E are the principal dietary antioxidants that protect erythrocytes from damage caused by ROS. 11 –15 The current study tested the hypothesis that dietary supplementation with vitamins E and C would lessen oxidant activity and oxidative damage in tibialis anterior muscles of aged rats subjected to chronic repetitive loading. 16 Indeed, several studies have reported that antioxidant supplementation attenuates oxidative stress in many physiological and pathological conditions. 17,18 On the contrary, other studies reported that these vitamins induce a pro-oxidant effect or no effect to redox status. 19,20 Therefore, the extent of the influence of supplementation with these antioxidants regarding aging-induced free radical bursts remains to be explored.
Although previous studies have already presented that supplementation of several antioxidants may have preventive effects on age-related pathophysiological and clinical conditions, little research has been done on changes in the mechanical properties and antioxidant capacity of RBCs induced by aging. Hence, the aim of the present study was to investigate the effects of vitamin C and E supplements on aging-induced free radical burst as well as its effect on the function of aging RBCs. In addition, recent findings concerning the availability of L-cysteine as a rate-limiting step in glutathione (GSH) synthesis led us to explore the effects of treatment with vitamins C and E on the influx rate of L-cysteine to reveal the antioxidant mechanisms of vitamins in RBCs.
Major reagents
All chemicals and reagents were purchased from Sigma-Aldrich (St. Louis, MO). Antibodies to Band 3 were obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Electrophoresis equipment and related supplies were obtained from American Bio-Rad Company. The spectrophotometer was purchased from American Thermo Company-Scientific. Laser rotating cell deformation instrument, LORCA, was purchased from Precil Company (Beijing, China).
Animal care
Animals used in this study were maintained in accordance with the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH publication no. 85-23, revised 1996), and all procedures were approved by the Institutional Review Board of the Institute of Health Sciences, Chongqing Institutes for Biological Sciences, Chinese Academy of Sciences, and Binzhou Medical University (Shandong, China).
Young (3-month-old) and aged (24-month-old) male Wistar rats were used for the experiments. Animals were fed according to the national standard rodent feed, with free feeding and drinking water. All rats were kept in the condition where relative humidity was 45%–55%, room temperature was 5 + 22°C, and the light time was 8:00–20:00. Rats were assigned based on age to two groups: young(Y) and aged (A) groups after 1 week of adaptive feeding, and each group of 20 rats was further divided among four groups: (i) control (Yc and Ac), (ii) vitamin C supplement (Yvc and Avc), (iii) vitamin E supplement (Yve and Ave), and (iv) vitamin C and vitamin E supplements (Yve+vc and Ave+vc).
The drug delivery system was according to the method of Sutcu R. 21 Briefly, the vitamin treatment groups were treated with, respectively, soluble olive oil, vitamin C soluble olive oil, and equal amount of vitamin E and vitamin C dissolved olive oil of 200 mg/kg (day) and continued for 6 weeks, while the control group was injected with the same amount of olive oil in the abdominal cavity for 6 weeks. The aged groups follow the above approach.
Hematocrit (HCT), mean corpuscular volume (MCV), mean corpuscular hemoglobin concentration (MCHC), mean corpuscular hemoglobin (MCH), and red cell distribution width (RDW) were tested using LH 780 hematology analyzer (Beckman Coulter, Fullerton, CA).
Blood sampling and generation of RBC suspensions
In each group, blood was collected by the method of heart blood sampling. By washing four times with phosphate-buffered solution (PBS), and then centrifuging at 900 × g at 4°C for 4 minutes, RBCs can be obtained and frozen for later use.
Detection of RBC antioxidant index
The activities of two antioxidant enzymes, catalase (CAT) and superoxide dismutase (SOD), and the levels of GSH were measured. The activities of CAT and SOD were assessed using the methods of Aebi 22 and Misra, 23 respectively. GSH levels were determined using the method described by Dringen and Hamprecht. 24
Measurements of methemoglobin levels
Blood was centrifuged at 2,000 rpm at 4°C to remove plasma, passed through cotton to eliminate white cells, and washed three times with choline wash solution (180 mM choline, 1 mM MgCl2, 10 mM Tris-Mops, pH 7.4 at 4°C, 320–340 mOsm). Red cell MetHb levels were determined as described by Kohn et al. Measurement of methemoglobin concentration was based on the absorbance of methemoglobin at 630 nm, which is characterized by ɛmM 630 = 4.4/mM (cm).
Determination of SH radicals in erythrocyte membranes
SH radicals of erythrocyte membrane proteins were quantified according to Anderson 25 and Yamaguchi. 26 SH radical levels were estimated by measuring absorbance at 415 nm after incubation at 37°C in a water bath for 15 minutes, the contents of SH radicals were determined using GSH as a standard.
Membranous peroxide (thiobarbituric acid-reactive substances) level detection
The extent of lipid peroxidation of RBC membranes was estimated by measuring thiobarbituric acid-reactive substance (TBARS) levels according to the method of Stocks and Dormandy. 27 TBARS levels were estimated by measuring absorbance at 532 nm after a reaction with thiobarbituric acid. Trichloroacetic acid extracts of RBC samples were used to circumvent the interference of proteins with TBARS determinations. Results were expressed as nanomoles per gram of hemoglobin (Hb). 28
Immunofluorescence and image analyses
RBCs were fixed with 4% paraformaldehyde and 0.05% glutaraldehyde in PBS and permeabilized in the same solution containing 0.05% Triton X-100. After being blocked with 3% BSA and 0.1% Tween 20 in PBS to block nonspecific protein binding, cells were treated with primary antibodies (rabbit polyclonal to Band 3; Abcam) diluted in 10 mg/mL PBS for 1 hour at room temperature. Cells were washed at 3-minute intervals thrice with gentle shaking, then incubated for 1 hour with secondary antibodies (anti-rabbit IgG; Santa Cruz Biotechnology, Santa Cruz, CA) at 1:700 dilution in PBS, and washed thrice in PBS. Fluorescence was imaged using an Olympus IX71 microscope with a 63/1.25 oil immersion objective and equipped with a CCD camera (Olympus, Tokyo, Japan).
Detection of RBC deformation
RBC deformability was determined by laser diffraction analyses using an ektacytometer (LBY-BX; Precil, Beijing, China) according to the method of Kucukatay V.
29
Based on the geometry of the elliptical diffraction pattern, the elongation index (EI) was calculated under shear stresses 0.3 and 30 Pa as follows:
where L and W are the length and width of the diffraction pattern, respectively. An increased EI at the given shear stress indicates greater cell deformation and hence greater RBC deformability. For studies using pervanadate, cell suspensions were preincubated with pervanadate (2 Mm; Beyotime, Shanghai, China) or vehicle for 10 minutes at 25°C before testing RBCs for cell deformability or anion transport activity.
In vitro model of oxidative stress
A hypoxanthine/xanthine oxidase (HX/XO) system was used to generate steady gradable production of superoxide. RBCs were portioned into aliquots and incubated with 1.5 mM HX and escalating doses of XO (0.2 U/mL) in a shaking heater block (37°C) for 2, 5, 10, 20, or 45 minutes, at which time 1 mM allopurinol was added to the sample to arrest superoxide production. Following HX/XO exposure, RBCs were washed thrice with PBS.
Detection of L-cysteine influx in erythrocyte
The procedure for measuring L-cysteine influx was essentially the same as described earlier. 30 A total of 0.25 mL of washed packed erythrocytes were suspended in 1 mL of PBS containing 8 mM glucose and 10 mM L-cysteine and incubated for 1 hour at 37°C in a water bath. Influx rate was calculated by subtracting the control (erythrocytes incubated in PBS–glucose without L-cysteine for 1 hour at 37°C) free–SH concentration from free–SH concentration obtained following treatment with L-cysteine.
Statistical analyses
All data, from three or more than three groups of repeated experiments, are expressed in the form of mean ± standard deviation. Intergroup differences between young and aged groups were assessed by independent sample t-test. The intragroup comparisons between treated groups and their respective control values were performed using one-way ANOVA. Analyses were performed with SPSS PC version 19.0. p < 0.05 means the difference was significant.
Results
Effects of Vc and Ve treatment on the antioxidant capacity of RBCs in rats of different ages
To explore the effect of Vc and Ve treatment on RBC antioxidant status, levels of GSH, CAT, and SOD were analyzed and the results are presented in Figure 1A–C. Compared with the young groups, the activity of antioxidant enzymes CAT (YC group, 73.3 ± 7.9 k/g Hb vs. AC group, 95.13 ± 5.2 k/g Hb; p < 0.01) and SOD (C group, 6231 ± 196 U/g Hb vs. AC group, 9308 ± 351 U/g Hb; p < 0.01) significantly increased in aged groups. Meanwhile, the levels of GSH significantly decreased (YC group, 22.02 ± 2.15 μmol/g Hb vs. AC group, 18.11 ± 2.66 μmol/g Hb; p < 0.05) (Fig. 1A).
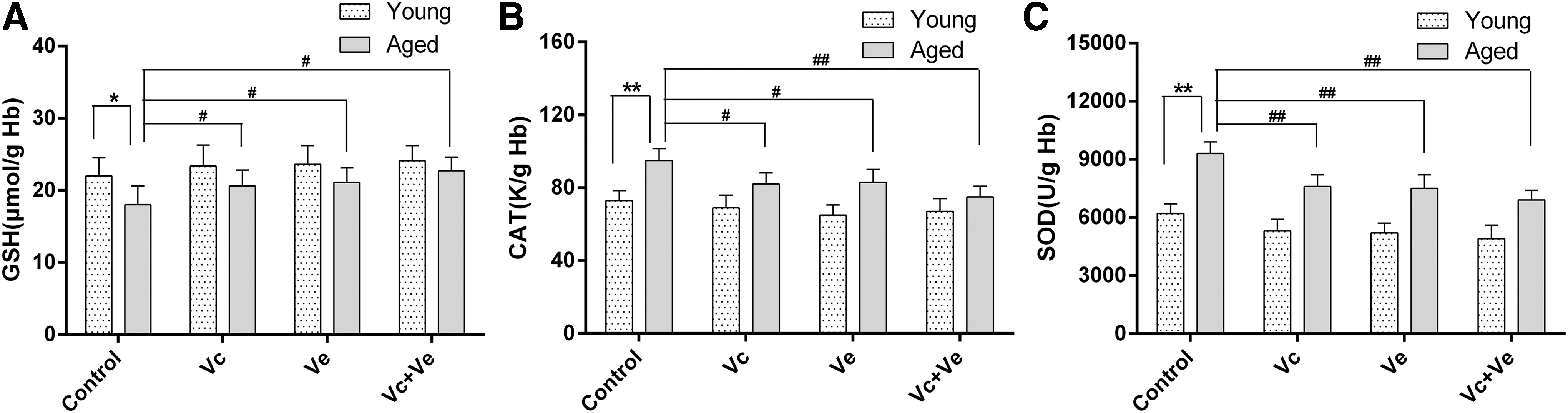
Erythrocyte antioxidant parameters in young and aged rats after Vc and Ve supplementation.
Within the aged groups, Vc and Ve treatment groups displayed lower CAT (AC group, 95.13 ± 5.2 k/g Hb; Avc group, 82.11 ± 4.2 k/g Hb; Ave group, 83.21 ± 5.5 k/g Hb; Avc+ve group, 75.69 ± 5.8 k/g Hb; p < 0.05, 0.05, and 0.01, respectively) (Fig. 1B, C) and SOD (AC group, 9308 ± 351 U/g Hb; Avc group, 7612 ± 313 U/g Hb; Ave group, 7507 ± 332 U/g Hb; Avc+ve group, 6931 ± 532 U/g Hb; p < 0.01, 0.01, and 0.01, respectively) activity compared with rats in control group. The GSH content increased significantly in Vc and Ve treatment groups (AC group, 18.11 ± 2.66 μmol/g Hb; Avc group, 20.60 ± 2.86 μmol/g Hb; Ave group, 21.15 ± 2.06 μmol/g Hb; Avc+ve group, 22.71 ± 1.96 μmol/g Hb; p < 0.05, 0.05, and 0.05, respectively) (Fig. 1A)
Effects of Vc and Ve treatment on the oxidative damage parameters of rats RBCs at different ages
To explore the impact of Vc and Ve treatment on RBC oxidant stress parameters, the levels of SH radical, TBARS, and MetHb were analyzed and the results are presented in Figure 2. Compared with the young groups, the free thiol groups in erythrocyte membranes significantly decreased (YC group, 81.2 ± 4.9 nmol/mg protein vs. AC group, 74.13 ± 5.2 nmol/mg protein; p < 0.05) and membrane lipid TBARS levels significantly increased (YC group, 12.8 ± 1.5 nmol/g Hb vs. AC group, 19.6 ± 1.6 nmol/g Hb; p < 0.01) in the aged group, while there is no significant change in the content of high iron hemoglobin (Fig. 2).

Erythrocyte oxidant stress parameters in young and aged rats after Vc and Ve supplementation.
Within the aged groups, Vc and Ve treatment groups displayed higher free thiol group levels of membrane protein (AC group, 74.13 ± 5.2 nmol/mg protein; Avc group, 80.12 ± 4.2 nmol/mg protein; Ave group, 78.61 ± 4.5 nmol/mg protein; Avc+ve group, 79.94 ± 4.8 nmol/mg protein; p < 0.05, 0.05, and 0.05, respectively) and significantly decreased membrane lipid TBARS levels (AC group, 19.6 ± 1.6 nmol/g Hb; Avc group, 15.6 ± 1.2 nmol/g Hb; Ave group, 15.9 ± 1.0 nmol/g Hb; Avc+ve group, 14.3 ± 1.9 nmol/g Hb; p < 0.05, 0.05, and 0.05, respectively) compared with rats in old control group (Fig. 2A, C).
Effects of Vc and Ve on rat RBC deformability at different age stages
RBC deformability (i.e., the EI) of the Y groups and the A groups was measured at shear stresses 3 and 30 Pa (Fig. 3). As expected, EI significantly declined in the AC group compared with the YC group under shear stresses (YC group, 0.42 ± 0.01 at 3 Pa and 0.571 ± 0.008 at 30 Pa; AC group, 0.38 ± 0.013 at 3 Pa and 0.534 ± 0.009 at 30 Pa; p < 0.05 and p < 0.01, respectively).In addition, the EI at 3 Pa shear stress increased significantly after Vc and Ve treatment.

RBC deformability as reflected by the elongation index in young and aged rats after Vc and Ve supplementation.
Table 1 demonstrates hematological parameter of young and aged rats as well as the effect of Vc and Ve treatment on these parameters. The MCV and RDW of aged rats were significantly higher compared with young rats. Vc and Ve treatment resulted in a significant decrement in MCV and RDW value and a significant increment in MCHC and MCH value of aged rats.
Values are mean ± SD. * p < 0.05 and ** p < 0.01, young vs. aged; †p < 0.05 and ††p < 0.01, control vs. vitamin supplement.
HCT, hematocrit; MCH, mean corpuscular hemoglobin; MCHC, mean corpuscular hemoglobin concentration; MCV, mean corpuscular volume; RDW, red cell distribution width; SD, standard deviation.
Oxidative damage parameters of rat RBCs in vitro
As shown in Figure 4, the oxidant stress parameters of erythrocytes collected from young and aged rats were tested under in vitro oxidant condition. There were significantly decreased free thiol group levels (81.10 ± 4.9 nmol/mg protein vs. 53.6 ± 4.9 nmol/mg protein in YC group; p < 0.05, and 74.13 ± 5.2 nmol/mg protein vs. 47.5 ± 4.2 nmol/mg protein in AC group; p < 0.05, respectively) and significantly increased TBARS levels (12.8 ± 1.5 nmol/g Hb vs. 23.6 ± 1.6 nmol/g Hb in YC group; p < 0.05, and 19.6 ± 1.7 nmol/g Hb vs. 25.7 ± 1.6 nmol/g Hb in AC group; p < 0.05, respectively). In both young and aged groups, Vc and Ve treatment led to significantly increased levels of free thiol groups and significantly decreased levels of TBARS.
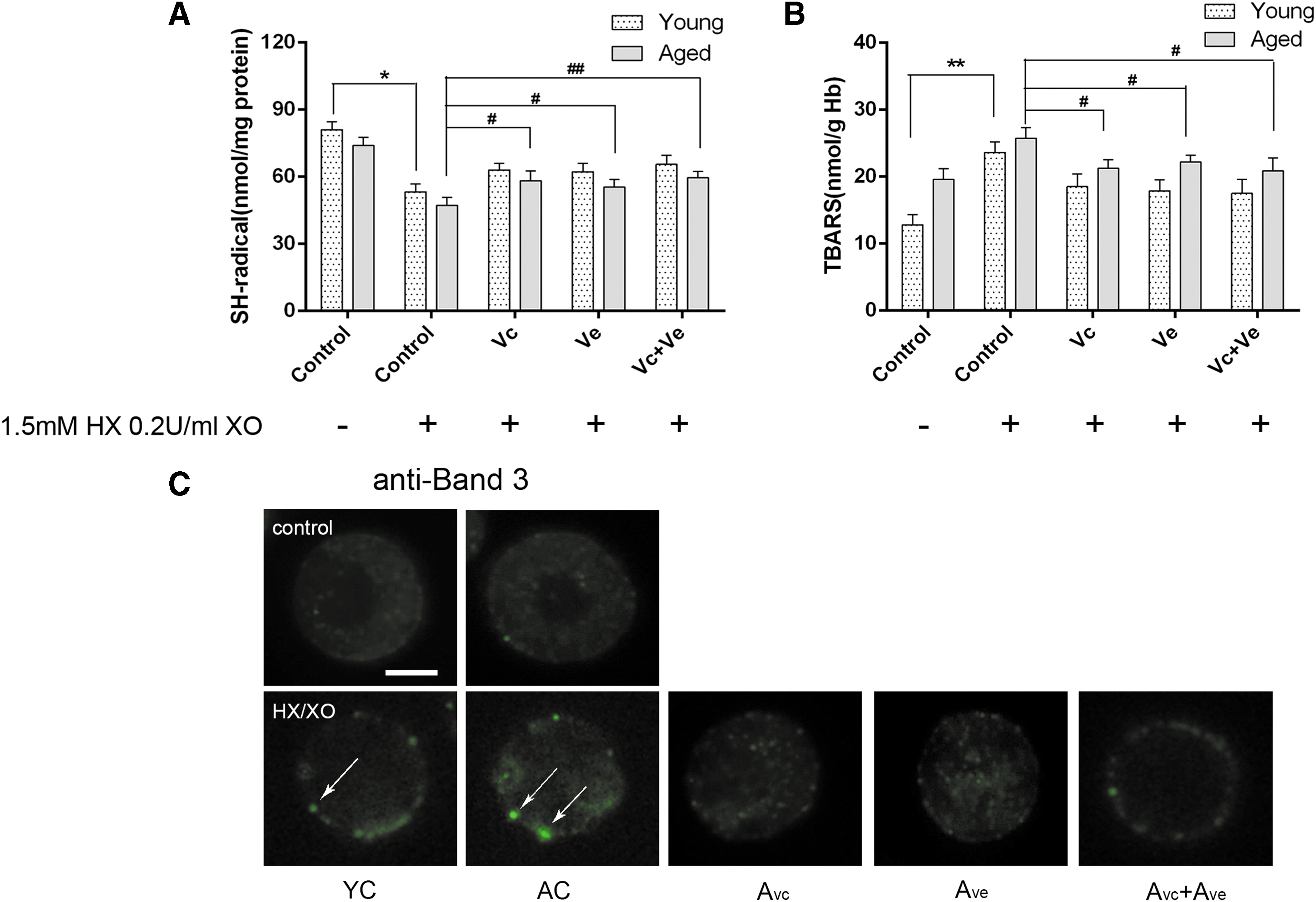
Erythrocyte oxidant stress parameters in young and aged rats after Vc and Ve treatment under in vitro oxidant condition.
Erythrocytes lack nuclei and other organelles and do not exhibit protein synthesis. Hence, oxidative stress-induced alterations in pre-existing proteins could result in recognizable changes at the membrane surface. Subsequent analyses of Band 3 by microscopy methods (Fig. 4C) verified the presence of Band 3 aggregates in a small proportion of cells formed after in vitro oxidant incubation in all tested groups; the Vc and Ve treatment resulted in significant repair of oxidative-induced protein cross-linking.
L-cysteine influx rate
To explore the mechanism of improving antioxidant capacity induced by Vc and Ve treatment, L-cysteine influx rate was tested under in vitro oxidant condition. As shown in Figure 5, L-cysteine influx rate was significantly decreased in control groups after the treatment of HX 0.2 U/mL XO 1.5 mM. L-cysteine influx rate significantly increased after Vc and Ve treatment (AC group, 1.11 ± 0.56 μmol/h (mL) Hb; Avc group, 1.80 ± 0.36 μmol/h (mL) Hb; Ave group, 1.75 ± 0.26 μmol/h (mL) Hb; Avc+ve group, 2.11 ± 0.31 μmol/h (mL) Hb; p < 0.05, 0.05, and 0.05, respectively) (Fig. 5A). Meanwhile, GSH content significantly increased in Vc and Ve treatment aged groups (AC group, 8.51 ± 2.66 μmol/g Hb; Avc group, 11.30 ± 2.86 μmol/g Hb; Ave group, 12.45 ± 2.06 μmol/g Hb; Avc+ve group, 14.51 ± 1.96 μmol/g Hb; p < 0.05, 0.05, and 0.05, respectively) (Fig. 5B).
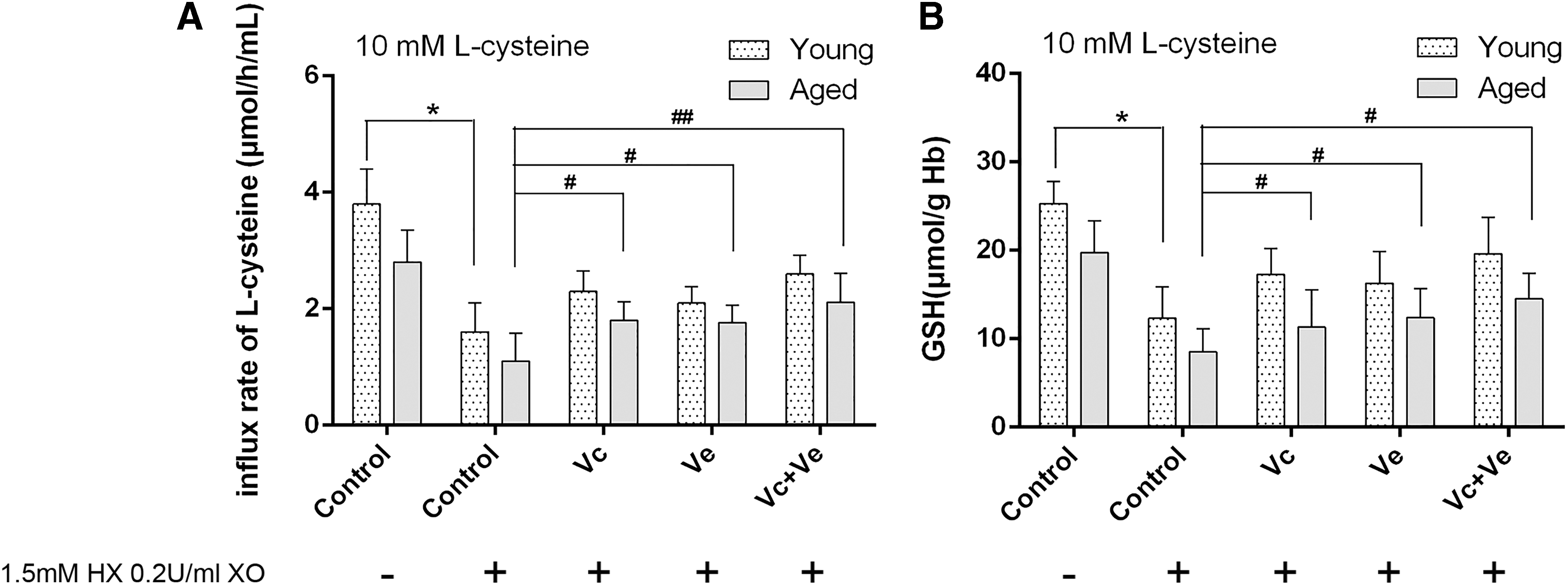
Erythrocyte L-cysteine influx rate and GSH levels in young and aged rats after Vc and Ve treatment under in vitro oxidant condition.
Discussion
Oxidative damage has long been implicated in the aging process. RBCs were continuously exposed to a large number of oxidants derived from various endogenous as well as exogenous sources during the aging process. Dietary antioxidant supplementation may be an effective treatment for the correction of impaired PMRSs, reducing the incidence of cardiovascular disease in the elderly. In this study, we found that supplementation with vitamins C and E could improve the antioxidant capacity and mechanical property of RBCs in aged rats. Based on in vitro observations, the influx rate of L-cysteine has been implicated in improving GSH levels after treatment with vitamins C and E.
As the oxygen transporter, mammalian erythrocyte is permanently in danger of oxidative damage across its lifetime due to the high cell concentration of oxygen and hemoglobin, a powerful promoter of the oxidative process. The antioxidant system of RBCs comprises enzymatic and nonenzymatic antioxidants, mainly comprising CAT, SOD, and GSH. 31 Under normal conditions, the synergistic effects of these molecules could remove most free radicals, including H2O2, O·2−, and OH·, maintaining the cellular redox balance. However, in a variety of physiological and pathological conditions, this redox balance was broken, resulting in oxidative damage to cell components, including hemoglobin and membrane proteins and lipids. 32 –35 In the present study, we observed a significant reduction of GSH in RBCs of aged rats, and supplementations with vitamins C and E effectively restored the level of GSH. There are two possible reasons that contribute to this phenomenon: (i) the group of aged rats produced more free radicals and the balance of GSH/GSSH shifts in the direction of oxidation and (ii) the supplementary rate of glutathione decreased in aged rats. Furthermore, the antioxidant enzyme test results show that the activity of SOD and CAT increased significantly in the aged rats, which was effectively attenuated by vitamin C and E treatment. These data suggest that the aged rats suffer more severe attacks by free radicals compared with younger rats and that supplementation with vitamins C and E can scavenge those free radicals from the plasma directly.
The functional role of RBCs is to transit through the capillaries to supply tissues with oxygen. Decreased deformability prevents the RBCs from responding to changing demands by easing the passage through the vessel, thereby decreasing the oxygen supply during metabolism and leading to age-related diseases. Recently, a study by Kucukatay et al. found that aging is associated with decreased RBC deformability and MCHC and an increase in erythrocyte MCV. 36 Aging of RBCs is closely related to a progressive decrease in the ADP/ATP ratio and finally leads to changes in the regulation of RBC deformability. 37 In the present study, we observed significant repair effects on the deformability of RBCs in aged rats. This result was consistent with the detection results of RBCs in each group. The improvement in antioxidant capacity by vitamin treatment reduced membrane protein cross-linking and lipid peroxidation damage induced by oxidative stress, eventually leading to improvement of the deformability of RBCs in aged rats. In addition, MCV and RDW of aged rats were found to be significantly higher compared with that of young rats. Meanwhile, vitamin C and E treatment resulted in a significant decrement in MCV and RDW. A decrease in cell volume (i.e., MCV) creates a more biconcave-disk shape with an increased surface area-to-volume ratio, which leads to increased ability of the cells to undergo deformation in aged rats.
Given that young and old rats were under different redox status in the in vivo environment, this may further affect the activity of certain antioxidant enzymes. Herein, to investigate the effects of vitamin C and E treatment on the antioxidant capacity of RBCs at different age stages, the oxidant stress parameters of erythrocytes collected from young and old rats were tested using in vitro oxidant conditions. Compared with young rats, there was a significantly higher level of membrane protein and lipid peroxidation in aged rats in vivo. Within aged groups, Vc and Ve treatment led to significantly increased levels of free thiol groups and significantly decreased levels of TBARS under in vitro oxidant conditions. In addition, the degree of cross-linking of the main membrane skeleton protein Band 3 was observed to increase. This oxidative cross-linking degree of Band 3 decreased considerably after treatment with vitamins C and E. This means that (i) the antioxidant capacity of RBCs in aged rats was lower than young rats and (ii) vitamin C and E treatment may play an effective role in improving the antioxidant capacity of RBCs in aged rats.
Previously, it was shown that L-cysteine is the only amino acid that has a functional free SH group. Given the presence of this thiol group, L-cysteine plays an important role in the elimination of free radicals and maintenance of a proper intracellular redox status in RBCs. 38,39 In addition, despite the fact that three amino acids were required for GSH synthesis, L-cysteine availability was the rate-limiting step of GSH synthesis. 40 A recent study found that the influx rate of L-cysteine was closely related to the membrane protein thiol content and the status of membrane lipids. 38 In this study, we observed an age-dependent decline in the influx rate of L-cysteine and a significant improvement after treatment with vitamins C and E. These results provide a potential molecular pathway by which vitamins C and E act on membrane proteins and lipids to improve the influx rate of L-cysteine, thus in turn enhancing the synthesis of GSH and ultimately leading to the improvement of the antioxidant capacity of RBCs. The resulting level of GSH detection is also consistent with our inference.
Conclusion
In summary, we present evidence of an improvement in the antioxidant capacity of RBCs by treatment with vitamins C and E in aged rats. These observations also suggest that treatment with vitamins C and E improves the synthesis of GSH by enhancing the influx rate of L-cysteine through the modification of membrane proteins and lipids. Our findings provide supporting evidence that treatment with vitamins C and E, at least partially, can improve the oxidative stress-induced age-related decrease in antioxidant capacity of RBCs.
Footnotes
Acknowledgment
Dr. Yanlian Xiong was supported by grants from the Initial Scientific Research Fund of Binzhou Medical University (BY2014KYQD09).
Author Disclosure Statement
No competing financial interests exist.