
Abstract
Vascular dysfunction is a common finding in type 2 diabetes, although the response to urotensin II (UII), a potent vasoconstrictor peptide, remains unclear. We investigated whether a UII-induced contraction was increased in the aortas from type 2 diabetic Goto-Kakizaki (GK) rats at the chronic stage. At 36 or 37 weeks of age (older group), a UII-induced contraction was seen in GK rats and was reduced by a Rho kinase inhibitor or urotensin receptor (UT) antagonist, whereas UII failed to induce a contraction in aortas from age-matched Wistar rats. In UII-stimulated aortas, the expression of Rho kinases, Rho A, and phosphorylated myosin phosphatase target subunit 1 did not change between the two groups; however, phosphorylation of extracellular-regulated kinase 1/2 and p38 mitogen-activated protein kinase (MAPK) was greater in GK than in Wistar rats. Compared to intact aortas, UII-induced contractions were slightly, but not significantly, increased by endothelial denudation of the aortas of Wistar rats at 24 weeks of age. At 6 weeks of age (young group), the UII-induced contractions were seen in GK and Wistar groups. The total expression and the membrane-to-cytosol ratio of the UT protein slightly decreased in Wistar aortas with aging but not in GK aortas. These results demonstrate that the UII-induced contraction gradually decreased with aging in Wistar rats and was preserved in type 2 diabetes. Although alterations of UII-induced contractions during aging and type 2 diabetes may be associated with kinase activities (MAPKs or Rho kinase) or receptor profiles, further investigations are necessary to clarify the mechanisms.
Introduction
V
Many endogenous substances regulate vascular function, including vasoactive peptides that are of considerable interest because their production and/or signaling are altered in type 2 diabetic arteries. 20,21 Urotensin II (UII), a cyclic dodecapeptide, is systemically present and is especially expressed in the central nervous system; moreover, the peptide is also present in a variety of peripheral tissues, including cardiomyocyte, endothelial, and vascular smooth muscle cells, and it can work in a hormonal, autocrine, or paracrine manner. 22 –27 UII has a variety of vasoactive effects such as control of vascular tone, proliferation, migration, and proinflammation. 22 –31 Although these UII effects are mediated by various intracellular signaling pathways, including those involving calcium mobilization, phospholipase C, protein kinase C, tyrosine kinases, mitogen-activated protein kinases (MAPKs), and the Rho/Rho-kinase pathway, through activation of the G-protein-coupled receptor GPR14 [UII receptor (UT)], 26,31 –36 the exact signal transduction mechanisms in type 2 diabetic arteries have not been investigated. Some interesting reports suggest that circulating levels of UII are increased in diabetic conditions, 37 –41 although the circulating levels are very low and often not detectable. 42 Together, these observations suggest a key role for UII in the pathogenesis of diabetes-associated vasculopathies.
Among animal models of type 2 diabetes, 43,44 the Goto-Kakizaki (GK) rat was produced by selective inbreeding of glucose-intolerant Wistar rats by a Japanese group. 44 These rats have mild hyperglycemia, glucose intolerance, defective glucose-induced insulin secretion, and are not obese, allowing for the investigation of molecular mechanisms independent of the confounding effects of obesity. 45,46 We 9 –11,13,14,47,48 and others 7,49 –51 have reported vascular dysfunction in this model. For example, in aortas from GK rats at chronic stages of the disease (i.e., 32–38 weeks of age) versus age-matched control Wistar rats, we found (1) augmentation of endogenous angiotensin II (Ang II, a vasoconstrictor) signaling, including Ang II-induced contractions, expression of Ang II, angiotensin-converting enzyme, and Ang II type 1 receptor 48 and (2) relaxation induced by acetylcholine (ACh) 10 and insulin. 48 Moreover, we found that impaired norepinephrine-induced contraction and enhanced ACh-induced relaxation due to the overproduction of nitric oxide (NO) via endothelial NO synthase (eNOS) and upregulation of the α2D-adrenoceptor occurs in early-stage GK rats (at 12 weeks of age). Conversely, impaired ACh-induced relaxation in later-stage GK rats (at 36 weeks of age) was attributable to reduced NO bioavailability. 47 The potent vasocontractile peptide ET-1 induced contraction that also increased in arteries from GK rats at various stages of disease. 11,21,50 Although contractile responses induced by various endogenous vasoconstrictors are altered in arteries from GK rats, no experimental studies on UII reactivity and molecular signaling have been performed using GK rats in early or chronic stages of diabetes.
In the present study, we first designed experiments to assess the alterations in the UII-induced aortic contraction that might occur as a result of long-term diabetes. We isolated aortas from 36- or 37-week-old GK rats and compared them with those from age-matched control Wistar rats and tried to identify some of the molecular mechanisms associated with the alterations that we detected. We next investigated the effect of the endothelium on UII-induced contractions in aortas from 24-week-old Wistar rats. Finally, to investigate age-related changes of UII-induced contraction, we examined the UII-induced contractions in aortas from 6-week-old GK and Wistar rats. We found that the UII-induced aortic contractions were gradually decreased with aging in Wistar rats and was preserved in GK rats.
Materials and Methods
Drugs
ACh chloride was purchased from Daiichi-Sankyo Pharmaceuticals (Tokyo, Japan). Phenylephrine, bovine serum albumin, and monoclonal β-actin antibody were purchased from Sigma Chemical Co. (St. Louis, MO). UII (human) was purchased from the Peptide Institute (Osaka, Japan). Y27632 and SB657510 were obtained from Tocris Bioscience (Ellisville, MO). Horseradish peroxidase-linked secondary antibodies were purchased from Promega (Madison, WI). Antibodies for MYPT1, Rho A, ROCK1, ROCK2, and caveolin-1 were purchased from BD Bioscience (San Jose, CA). The antibody for phospho-MYPT1 (Thr853) was purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Antibodies for p38 MAPK, phospho-p38 MAPK (Thr180/Tyr182), ERK1/2, and phospho-ERK1/2 (Thr202/Tyr204) were purchased from Cell Signaling Technology (Beverly, CA). The antibody for UT and its control peptide were purchased from Enzo Life Sciences (Plymouth Meeting, PA). The antibody for GAPDH was purchased from GeneTex (Irvine, CA).
Animals and experimental design
Four-week-old male rats (GK and control Wistar) were supplied by CLEA (Tokyo, Japan). Food and water were provided ad libitum in a controlled environment (room temperature: 21–22°C, humidity: 50% ± 5%) until the rats were 6, 24, 36, or 37 weeks old. This study was approved by the Hoshi University Animal Care and Use Committee, and all studies were conducted in accordance with the Guide for the Care and Use of Laboratory Animals, published by the US National Institutes of Health and with the Guide for the Care and Use of Laboratory Animals adopted by the Committee on the Care and Use of Laboratory Animals of Hoshi University (accredited by the Ministry of Education, Culture, Sports, Science, and Technology, Japan). At the time of the assays, the nonfasted blood glucose levels were measured using a glucose meter (OneTouch Ultra; LifeScan, a Johnson & Johnson Company, Milpitas, CA).
Measurement of isometric force (vascular functional study)
The vascular isometric force in the aortic rings was recorded as described in our previous articles. 13,14,16,17,52 Each aorta was separated from the surrounding connective tissues and cut into 2 mm long rings. The rings were stretched until an optimal resting tension of 2.75 g was loaded and allowed to equilibrate for at least 45 minutes. Force generation was monitored using an isometric transducer (Nihon Kohden, Tokyo, Japan). A concentration–response curve for UII (10−10–10−7 M) was constructed. To investigate the effect of a UT antagonist and Rho kinase inhibitor on UII-induced contractions, we examined UII-induced contractions in the presence of 10−6 M SB657510 (UT antagonist) and 10−6 M Y27632 (Rho kinase inhibitor). Rings were incubated with the appropriate drugs for 30 minutes before administration of UII.
In a second series functional study, we performed a concentration–response curve for UII (10−10–10−7 M) in aortas in the presence and absence of endothelia isolated from 24-week-old Wistar rats. Endothelial denudation was prepared by gently rubbing the intimal surface, and a successful denudation was confirmed by assessing the inability of an ACh (10−5 M)-induced relaxation precontracted with phenylephrine (10−6 M).
In a third series functional study, we performed a concentration–response curve for UII (10−12–10−7 M) in endothelium-intact aortas isolated from 6-week-old GK and Wistar rats.
Western blotting
Western blotting and data analyses were conducted as described in our previous articles. 13 –15,53,54 To evaluate the activation of Rho kinase, ERK1/2, and p38 MAPK pathways, each aortic ring was placed in the Krebs–Henseleit solution at 37°C for 30 min and then treated with UII (3 × 10−9 M) for 20 min. To evaluate the expression of UT in the membrane and cytosolic fractions, the membrane and cytosolic proteins of aortic tissue were extracted using a Mem-PER™ Plus kit (#89842, ThermoScientific, Rockford, IL) following the manufacturer's protocol. The following primary antibodies were used: anti-UT (∼100 kDa; 1:1000), anti-Rho A (∼21 kDa; 1:1000), anti-ROCK1 (∼160 kDa; 1:1000), anti-ROCK2 (∼160 kDa; 1:1000), anti-phospho-MYPT1 (∼130 kDa; 1:200), anti-MYPT1 (∼130 kDa; 1:1000), anti-phospho-ERK1/2 (42 and 44 kDa; 1:1000), anti-ERK1/2 (42 and 44 kDa; 1:1000), anti-phospho-p38 MAPK (43 kDa; 1:1000), anti-p38 MAPK (43 kDa; 1:1000), anti-GAPDH (36 kDa; 1:5000), anti-caveolin-1 (24 kDa; 1:1000), and anti-β-actin (42 kDa; 1:5000). The specificity of the UT antibody was evaluated using blocking peptides according to the manufacturer's instructions. The bands were quantified and analyzed using CS Analyzer 3.0 software (ATTO, Tokyo, Japan). The phosphorylation levels of MYPT1, ERK1/2, and p38 MAPK were normalized by total MYPT1, total ERK1/2, and total p38 MAPK, respectively, and then presented as a fold increase (relative to Wistar). The protein expression values for UT, RhoA, ROCK1, and ROCK2 were normalized by β-actin and then presented as a fold increase (relative to Wistar). To assess the successful isolation of the membrane and cytosol fractions, caveolin-1 and GAPDH proteins were detected as markers of the membrane and cytosol, respectively. To evaluate the subcellular localization of the UT receptor, we performed densitometry and calculated the membrane-to-cytosolic ratio of the UT receptor using the absolute values of the intensity of the UT receptor band (100 kDa) of each fraction from the aorta of the same rat.
Statistical analyses
The contractile force exerted by the aortic rings is expressed as a percentage of the 80 mM KCl-induced contraction, and such contractions were not significantly altered among groups at 36 or 37 weeks of age (data not shown). Data are presented as mean ± standard error. For the aortic functional studies, individual concentration–response curves were assessed by a nonlinear regression curve fitted using the variable slope model (namely, a four-parameter dose–response curve) (version 5.0; GraphPad Software, Inc., San Diego, CA). Statistical analyses were performed using a Student's t-test to compare two groups and a one-way analysis of variance (ANOVA) followed by Bonferroni's test to compare more than three groups. Statistical evaluations of the concentration–response curves were conducted by a two-way ANOVA with repeated measures followed by a Bonferroni post hoc test. A p value <0.05 was considered statistically significant.
Results
Body weight and blood glucose
As shown in Table 1, body weight was slightly and significantly lower in GK than in Wistar rats at 6 weeks of age and 36 or 37 weeks of age, respectively. Blood glucose levels (nonfasted) were significantly greater in GK than in Wistar rats at both 6 weeks of age and 36 or 37 weeks of age.
Values are mean ± SE. Number of animals is shown within parentheses.
p < 0.05 vs. 6-week Wistar group.
p < 0.05 vs. age-matched Wistar group.
p < 0.05 vs. 6-week GK group.
GK, Goto-Kakizaki; SE, standard error.
UII-induced contraction in aortas from 36- or 37-week-old GK and Wistar rats
To determine whether UII-induced contractions of aortas from type 2 diabetes at the chronic stage would be greater than those from age-matched controls, we compared the contractions of aortas from 36- or 37-week-old GK and Wistar rats.
As shown in Figure 1 and Table 2, exposure of the aortic rings to UII (10−10–10−7 M) led to a concentration-dependent rise in tension in GK rats. Interestingly, the contractile response of UII (10−10–10−7 M) was not seen in control aortas [absolute contractile force: 0.24 ± 0.05 mN/mg (n = 7)].
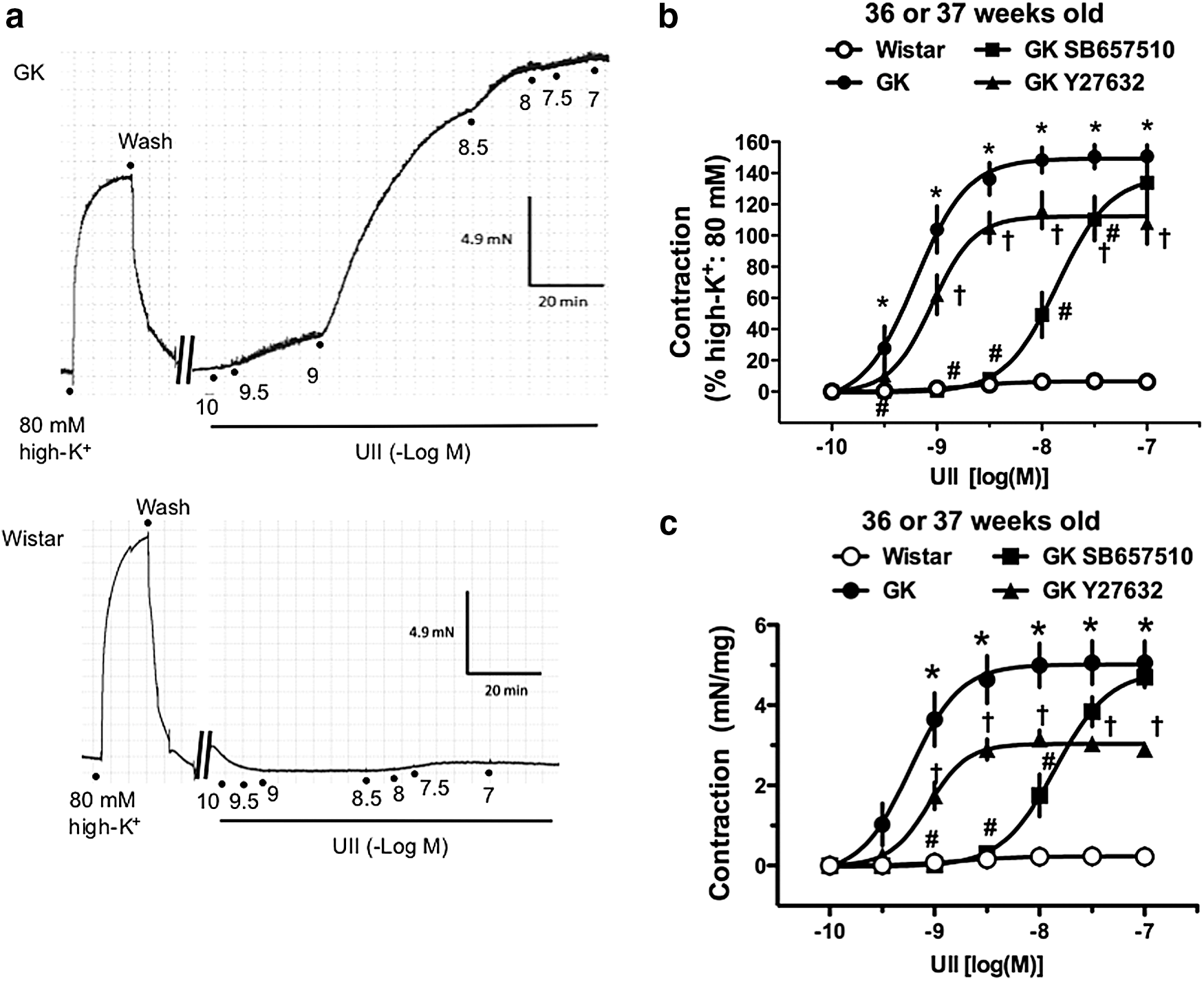
Urotensin II (UII)-induced contractions are seen in aortas from GK rats but not from control Wistar rats at 36 or 37 weeks of age.
Values are mean ± SE. Number of animals is shown within parentheses. Variables were determined by curve fitting data points from cumulative drug concentration–response relationships to a four-parameter logistic function (variable slope model).
Significant difference from the untreated GK group (p < 0.05 by Bonferroni's test).
UII, urotensin II.
Effects of a UT antagonist and Rho kinase inhibitor on UII-induced contraction in aortas from 36- or 37-week-old GK and Wistar rats
As shown in Figure 1b, c, and Table 2, the UT antagonist SB657510 (10−6 M) markedly shifted the UII-induced contractions in aortas from GK rats rightward. Moreover, the specific Rho kinase inhibitor Y27632 (10−6 M) significantly reduced the UII-induced aortic contraction in GK rats. The hill slope values were not altered among any of the groups (Table 2).
Evaluation of Rho kinase-associated protein expression
Using pharmacological approaches, the UII-induced aortic contraction in GK rats was shown to be partly mediated by the Rho kinase pathway (Fig. 1b, c). To investigate the possible mechanisms underlying the alterations in the UII-induced contraction in aortas from GK rats, we investigated the expression of Rho kinase-related proteins such as Rho A (Fig. 2a, b), ROCK1 (Fig. 2c, d), ROCK2 (Fig. 2c, e), and phosphorylated MYPT1 (a substrate for Rho kinase related to vascular contraction 4,55,56 ) (Fig. 2f, g) in aortas stimulated with UII (3 × 10−9 M for 20 min) isolated from rats at 36 or 37 weeks of age. Unexpectedly, the expression of all of these proteins was not significantly altered between the GK and Wistar groups.

Immunoblots for RhoA/Rho kinase components in UII-stimulated aortas from GK and Wistar rats at 36 or 37 weeks of age. Details are described in the Materials and Methods Section.
Effects of the endothelium on UII-induced contractions in aortas from 24-week-old Wistar rats
Reportedly, endothelium substances suppressed a UII-induced contraction. 22,57,58 To investigate whether endothelium-derived factors could be modulated to a UII-induced contraction, we compared contractile responses to UII in the presence and absence of endothelia in aortas from 24-week-old Wistar rats (Fig. 3). The UII-induced contraction was slightly but not significantly increased in endothelium-denuded (vs. endothelium-intact) preparations. Interestingly, when the UII-induced contraction in aortas from Wistar rats was compared between 24 weeks and 36 or 37 weeks of age, the contractile force was greater in the younger rats [absolute contractile force: 0.70 ± 0.15 mN/mg (n = 7)]. Therefore, we further investigated whether the UII-induced contraction would be greater in younger rats and would be different between GK and Wistar rats.
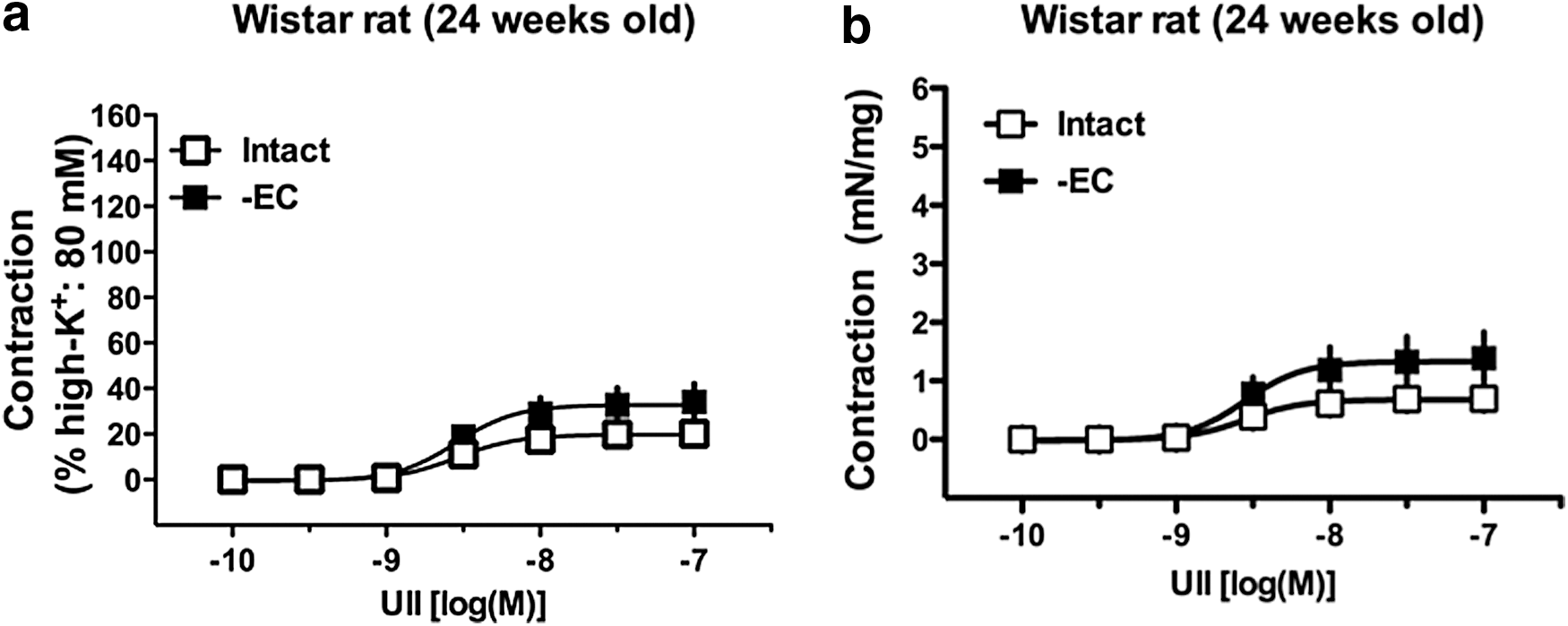
Effects of endothelial denudation on UII-induced contractions in aortas from 24-week-old Wistar rats. Concentration–response curves for UII (10−10–10−7 M) in endothelium-intact or endothelium-denuded (-EC) aortas from Wistar rats. Data are mean ± SE. n = 7 (intact) or n = 6 (-EC). The y-axis shows the contraction as a percentage of high-K+ (80 mM)-induced contraction
UII-induced contraction in aortas from 6-week-old GK and Wistar rats
To determine whether a UII-induced contraction would be present in aortas from younger Wistar rats and whether the contraction would be altered between GK and Wistar rats, we performed concentration–response curves for UII (10−12–10−7 M) in endothelium-intact aortas from 6-week-old GK and Wistar rats. As shown in Figure 4 and Table 3, UII led to an increased aorta tension in both GK and Wistar groups and the UII-induced contraction shifted leftward in GK versus Wistar groups. As expected, the contractile force was greater in younger Wistar rats [absolute contractile force: 5.72 ± 0.53 mN/mg (n = 5)] (Table 4).

Concentration–response curves for UII (10−12–10−7 M) in aortas from 6-week-old GK and Wistar rats. Data are mean ± SE. n = 5. The y-axis shows the contraction as a percentage of high-K+ (80 mM)-induced contraction
Values are mean ± SE. Number of animals is shown within parenthesis. Variables were determined by curve fitting data points from cumulative drug concentration–response relationships to a four-parameter logistic function (variable slope model).
Significant difference from the Wistar group (p < 0.05 by Student's t-test).
Values are maximal contractile response expressed as (mN/mg tissue) and are mean ± SE. Number of animals is shown within parentheses.
NA, not assessed.
UT expression in aortas from GK and Wistar rats
To investigate whether an alteration of the UT receptor expression was involved in the UII-induced aortic contraction, we measured the expression of UT in aortas isolated from young (6 weeks old) and old (36 or 37 weeks old) rats (Fig. 5). To determine the specificity of the antibody against UT, we conducted immunoblotting using the UT primary antibody with and without incubation with the appropriate antigenic peptide (Fig. 5a). As shown in Figure 5a, only the band at ∼100 kDa disappeared when the primary antibody was preincubated with the blocking peptides, although multiple bands were detected in the aorta. Therefore, we identified the band (at 100 kDa) as a putative UT receptor. The UT protein expression (at 100 kDa) did not change in aortic tissues among the four groups (Fig. 5b, c).
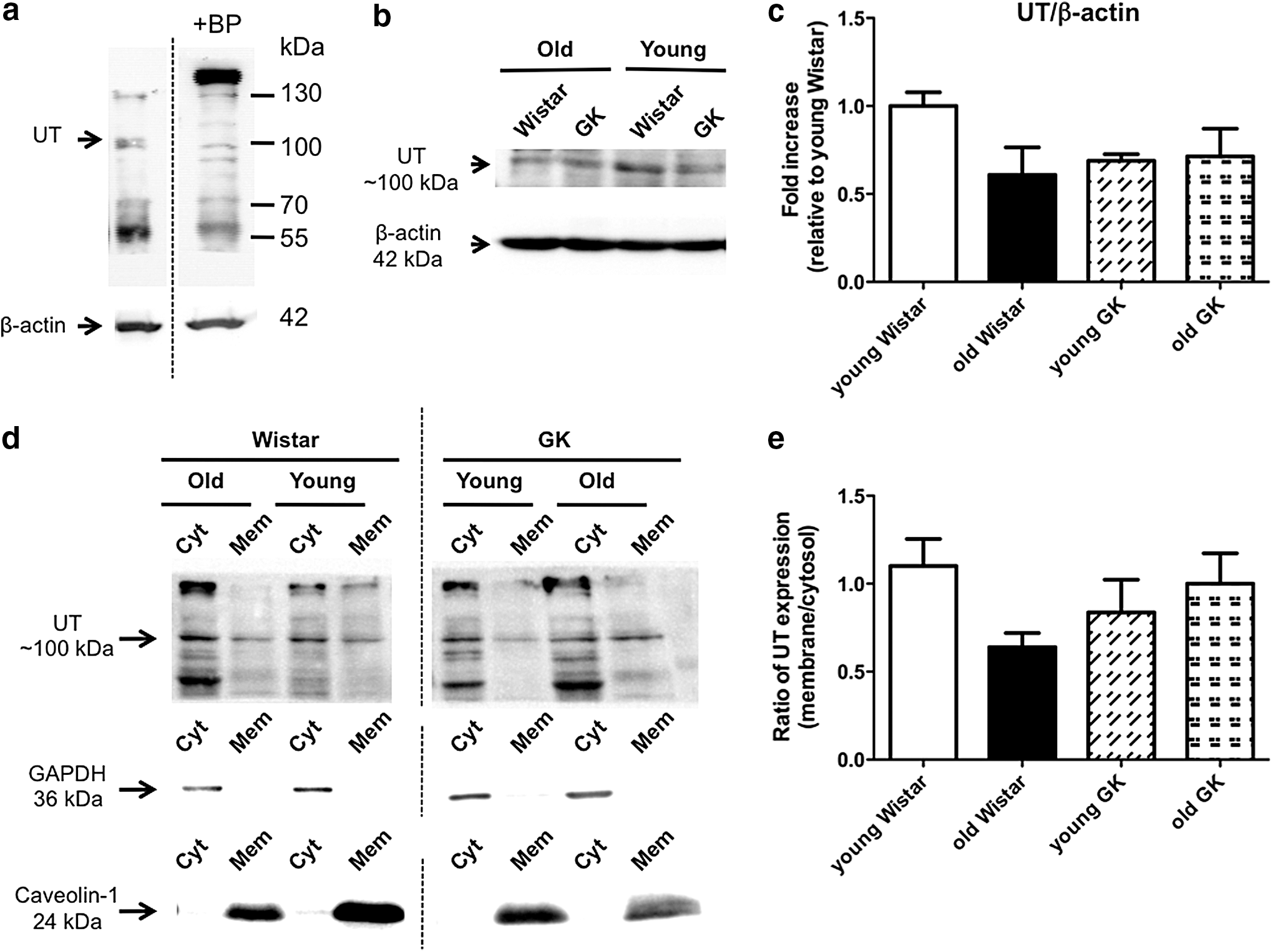
Immunoblots for the urotensin II receptor (UT) in aortas from GK and Wistar rats.
Reportedly, plasma membrane UT receptors are associated with a contractile response to UII in rat aortas. 59 To investigate whether the localization of UT would change between the membrane and cytosol in aortas with aging and diabetes, we detected the UT of aortas under a basal condition (i.e., without UII stimulation) in each membrane and cytosolic protein fraction (Fig. 5d, e). The UT expression was seen in each membrane and cytosolic fraction, and the purities were evaluated with representative markers, such as caveolin-1 (membrane) and GAPDH (cytosol), in young and old diabetic/control rats (Fig. 5d). The UT membrane-to-cytosol ratio was slightly decreased (but not significantly) in control, but not diabetic, aortas with age (Fig. 5e).
Effects of UII on ERK1/2 and p38 MAPK activation
Reportedly, UII affects vascular function through the activation of MAPK pathways, including ERK1/2 and p38 MAPK.
29
–31,60,61
To investigate whether MAPK activation would increase in diabetic aortas stimulated with UII, we assessed the expression of phosphorylated ERK1/2 and p38 MAPK, which are the activated forms.
11,51,62
As shown in the aortas from 36- or 37-week-old rats (Fig. 6), the phosphorylation of ERK1/2 (Fig. 6a) and p38 MAPK (Fig. 6b) was significantly greater in GK rat aortas stimulated with UII (3 × 10−9 M for 20 min) compared with the controls. In vehicle-treated aortas (i.e., no stimulation with UII) from 36–37-week-old rats, the expression levels of phosphorylated ERK1/2, total ERK1/2, phosphorylated p38 MAPK, and total p38 MAPK did not differ between GK and Wistar groups (Supplementary Fig. S1; Supplementary Data are available online at
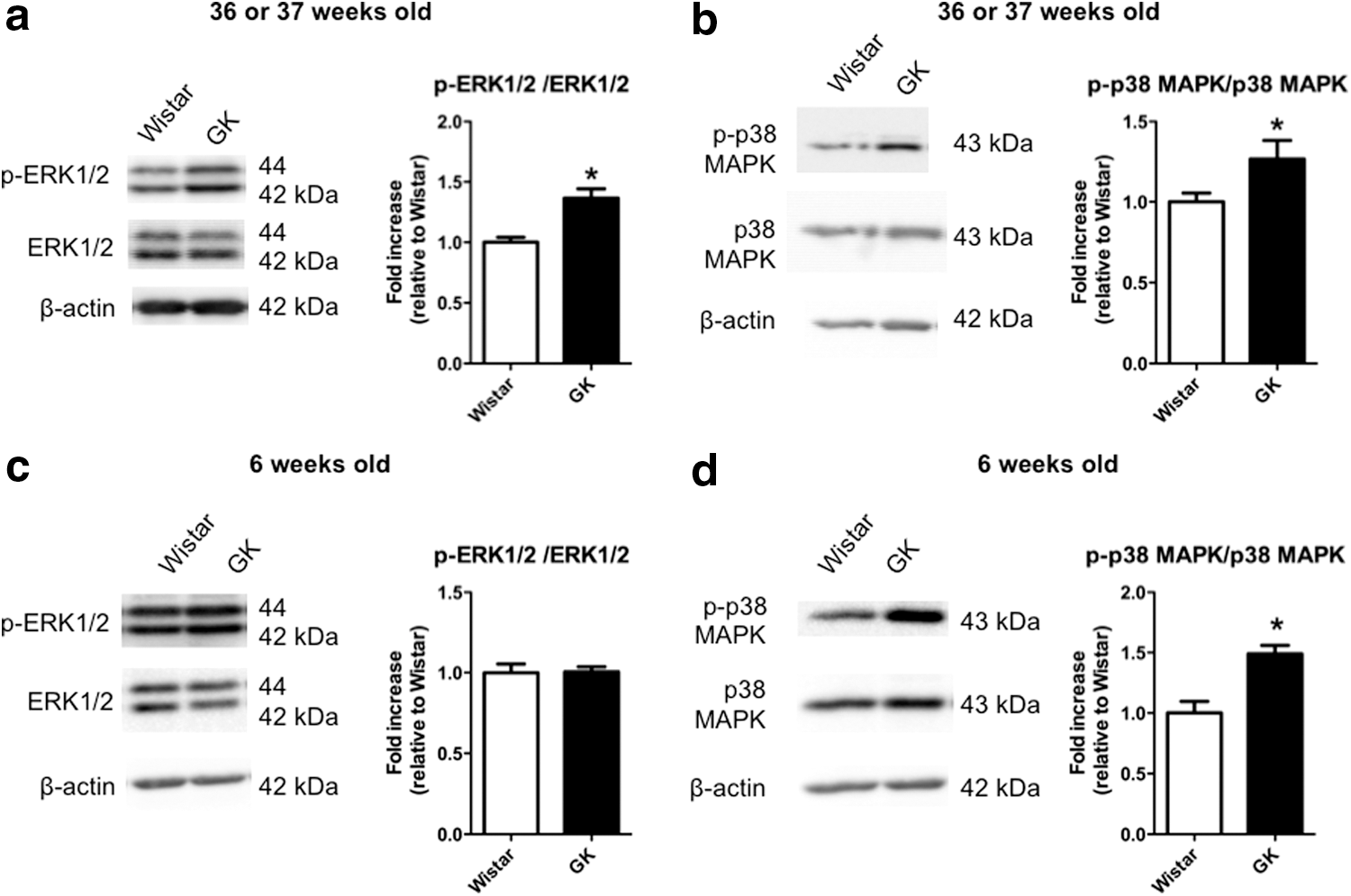
Immunoblots for phosphorylated and total extracellular-regulated kinase 1/2 (ERK1/2) and p38 mitogen-activated protein kinase (MAPK) in UII-stimulated aortas from GK and Wistar rats at 36 or 37 weeks of age
Discussion
The role of UII in the cardiovascular system and in diseases remains largely unknown. Recent evidence has revealed an increased expression of UII and the UT receptor in cardiovascular diseases, including atherosclerosis, hypertension, and diabetes, suggesting a potential role of UII in the development of such diseases. 24,25,27,37,63 –65 The GK rat is a unique, nonobese model of type 2 diabetes. 45,46 Although there are numerous reports of vascular dysfunction in this diabetic model, 7,9,10,11,13,14,47 –51 no previous study has investigated the responsiveness and mechanisms involving UII in this model. Moreover, few reports have investigated the age-related changes of a UII-mediated response in rat aortas. In the present study, for the first time, we demonstrated that the UII-induced contraction in isolated aortas was preserved with aging in GK rats, but an age-related decrease of a UII-induced contraction was seen in the aortas of control Wistar rats. The mechanisms underlying these different responses to UII between type 2 diabetic GK rats and Wistar rats may be partly attributable to the activities of Rho kinase and/or MAPKs (namely, ERK1/2 and p38 MAPK) or to the alteration of UT profiles.
It is known that the UT is a G-protein-coupled receptor that is coupled to the Gq-mediated signaling pathways to liberate inositol phosphates and mobilize intracellular Ca2+. 23 –26 Under normal physiological states, the UT expression is low in human and rodent arteries. 58,66 The contractile force of the thoracic aorta is much more than that of the abdominal aorta in rats because of the greater density of UT receptors in rat thoracic aortas. 65,67 The coronary artery in humans has a lower density of UT receptors compared with rat aortas, which may account for the greater contraction induced by UII in the rat aorta (vs. the human coronary artery). 58 In contrast, upregulation of the UT was seen in disease conditions, including atherosclerosis, hypertension, end-stage heart failure, renal disease, and diabetes. 20,41,65,68 Lim et al. 64 demonstrated that UII leads to differential responses in skin microcirculation between normal subjects (i.e., dose-dependent vasodilation) and chronic heart failure patients (i.e., dose-dependent vasoconstriction). Thus, the difference in contractile responses to UII may be attributable to a differential expression of functional UT receptors. Indeed, in the present study, the UII-induced contraction in the diabetic aorta was greatly attenuated by a selective antagonist of the UT receptor. When we detected the UT receptor using western blotting, multiple bands were detected in the aorta. Multiple bands for the UT receptor have been reported previously 69,70 and could result from the homo- or hetero-oligomerization of the receptor or its post-transcriptional modification, including glycosylation. 69 –71 Notably, when the primary antibody was preincubated with the appropriate antigenic peptide, only the band at 100 kDa disappeared. Therefore, we further investigated this band as the UT receptor in the present study. Under a basal condition, the UT expression in whole aortic tissues was not significantly altered between diabetic and control rats, and aging did not influence the expression, although it was slightly decreased in aortas from old Wistar rats compared to young ones. These results suggest that the UII-induced contraction may not only be associated with the UT receptor expression level (i.e., quantity) but also the nature of the UT receptor (i.e., quality), as discussed below.
Activation of the Rho kinase pathway in arterial smooth muscle is one of the key steps leading to contraction. 4,55,56,72,73 Indeed, several reports suggested that UII affects vascular function through the activation of such a pathway. 34,72 Moreover, there are several reports by us and others that demonstrated enhanced contractions induced by various constrictors that were associated with increased Rho kinase signaling in diabetic conditions. 8,12,14 In the present study, we found that the UII-induced aortic contraction in GK rats was partly inhibited by the Rho kinase inhibitor Y27632. These results suggest that in type 2 diabetic aortas, the Rho kinase pathway may be partly responsible for the UII-induced contraction. It was noteworthy that although the UII-induced contraction was partly reduced by the Rho kinase inhibitor in aortas, some contraction remained in diabetic aortas under such conditions. Moreover, the expression of Rho kinase-related components (namely, Rho A, ROCK1, ROCK2, and phosphorylated MYPT1) was not changed in UII-stimulated aortas. Judging from these results, the UII-induced aortic contraction seen in GK rats was partly attributable to the activation of not only the Rho kinase pathway but also other signaling pathways.
The idea that MAPKs, such as ERK1/2 and p38 MAPK, play an important role in the control of vascular function in (patho)physiological conditions has received a great deal of attention. 11,14,17,51,53,62 Moreover, several reports suggested that UII affects vascular function via activation of ERK1/2 and p38 MAPK. 29 –31,34,60,61 Although the importance of ERK1/2 and p38 MAPK in vascular dysfunction has been recognized, a complete assessment of the possible relationship between these MAPKs and vasoconstriction induced by UII in diabetes had not been made; thus, we investigated this issue using UII-stimulated aortas from GK rats. In the present study, we found that increased phosphorylation of ERK1/2 and p38 MAPK in UII-stimulated GK aortas (vs. controls) was seen in aortas from older animals, although basal levels (i.e., unstimulated conditions) of phosphorylated ERK1/2, ERK1/2, phosphorylated p38 MAPK, and p38 MAPK were similar between the GK and Wistar groups, and increased phosphorylation of p38 MAPK was seen in GK aortas (vs. controls) in younger animals. These results suggest that a sustained UII-induced aortic contraction in aged GK rats may be partly attributable to activation of these MAPKs. However, the mechanisms underlying the development of ERK1/2 phosphorylation (activation) and a sustained increment of p38 MAPK phosphorylation in diabetic aortas with aging remain unclear. In the present study, we investigated the levels of intracellular molecules associated with UII-induced contraction on UII stimulation for 20 min, as several reports suggest that this time is adequate to obtain adequate responses to stimulation by UII in various tissues and cells. 31,35,74 Moreover, long-lasting effects of UII were also reported. 32,35,74 Therefore, further investigations will be required to understand the time course of UII stimulation, both in terms of contractile response and the activity/expression of related molecules in diabetic aortas.
In the present study, we demonstrated that the UII-induced contraction was preserved in diabetic but not in control aortas and that such a contraction decreased in Wistar rats as they aged. It is known that endothelial dysfunction is exhibited in various arteries of type 2 diabetes, including GK rats. 11,21,50 Indeed, the endothelium-dependent relaxation was impaired in aortas from GK rats at the chronic stage of disease. 10,48 Although endothelium-derived substances are able to modulate the contraction induced by endogenous vasoconstrictors, 11,57 the contribution of the endothelium in the UII-induced contraction may be minor at the chronic stage of diabetes, even though endothelial dysfunction is present. This is supported by Ishihata et al., 57 who demonstrated that a UII-induced contraction was not modulated by endothelium denudation, NOS inhibitor, or a cyclooxygenase inhibitor in aortas from aged (25 to 27 months) male Fischer 344 rats. Moreover, in the present study, a UII-induced contraction was not enhanced by endothelial denudation in aortas from 24 weeks of age. Our present data in the series of time course observations of UII-induced responses suggest that functional coupling between UT and the vasocontractile components in the aorta has already developed and been maintained in long-term diabetic states. At present, the causal factors and mechanisms that differ between GK and controls remain unknown, although the quality and quantity of functional UT receptors may vary in these conditions. Several reports have suggested that UII has a tachyphylactic effect in vascular cells. 75,76 The contractile force induced by UII may be defined not only by the UT receptor density but also by UT receptor trafficking, including desensitization, internalization, and recycling. For example, Giebing et al. 59 suggested that the long-lasting UII-induced aortic contraction was attributable to continuous externalization (recycling) of UT receptors. Moreover, Lecointre et al. 77 demonstrated that a signaling switch of UT coupling with a G-protein occurred that involved UT/G13/Rho/Rho kinase or UT/Gi/o/phosphatidylinositol-3 kinase components to distinguish cellular processes such as migration and adhesions in glioma cells. In the present study, we found in control but not GK aortas that not only the total aortic homogenate but also the ratio of the UT receptor in membrane-to-cytosol tended to decrease with aging as well as the contractile response to UII. Moreover, the concentration–contractile relationship with UII appeared to have a Hill constant larger than 1.0 in all groups. These results suggested that the mechanisms of the ligand and receptor interaction are complicated and that some kind of positive cooperation may be involved in the UII-induced contraction. How and to what extent the coupling with the UT receptors and contractile machinery and nature of the UII-induced receptor activation, including its trafficking in type 2 diabetes, remain unclear, necessitating further investigation on this point.
An accumulating body of evidence suggests that UII signaling is involved in the development of diabetes and diabetes-associated complications. For example, local and systemic levels of UII are increased in diabetic patients and animal models of the disease. 22,37 –41,68,78 Clozel et al. 79 demonstrated that the orally active selective UT antagonist palosuran could improve pancreatic and renal function in diabetic rats. Watson et al. 78 demonstrated that the selective UT antagonist SB657510 could reduce diabetes-associated atherosclerosis in diabetic Apoe-knockout mice. Considering the evidence above and the present results, we suggest that the manipulation of arterial UII/UT signaling could be a therapeutic target in diabetes-associated vascular dysfunction. However, further investigations will be required to comprehensively understand UII/UT signaling by time, region, and condition (e.g., diseases).
In conclusion, we have for the first time demonstrated that a UII-induced contraction was specifically preserved in type 2 diabetic GK aortas and that this may be partly attributable to increased kinase activities (MAPKs or Rho kinase) or altered receptor profiles. We believe that our findings should stimulate further interest in the control of UII/UT signaling in the continuing efforts to reduce diabetes-associated vasculopathies.
Footnotes
Acknowledgments
We thank M. Iguchi, M. Nagata, M. Takahashi, A. Suwa, J. Nomoto, S. Hotozuka, M. Majima, M. Takeuchi, and H. Sashikubi for the excellent technical help. This work was supported, in part, by JSPS KAKENHI Grant Numbers JP26460107, JP15K21419, and JP15K07975. The authors thank Enago (
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.