
Abstract
As superoxide is a key molecule of inflammatory activation, superoxide-hydrogen peroxide (S-HP) imbalance genetically caused could alter immunosenescence patterns. To test this hypothesis, we collected and cultured peripheral blood mononuclear cells (PBMCs) carrier's different genotypes of a genetic polymorphism located in the superoxide dismutase manganese-dependent gene (Val16Ala-SOD2). We used an in vitro genetic model based on previous studies, which suggested an association between homozygous genotypes (AA and VV) and alterations in oxidative–inflammatory mediators. PBMCs collected from young healthy volunteers were cultured in the presence of phytohemagglutinin, as well as the following cell culture passages obtained from the 72-hour initial culture. Each follow passage started with the same cell concentration (1 × 105 cells). The general immunosenescence pattern was observed independent of SOD2 genotypes: cellular proliferation until the 15th passage, when cellular arrestment occurred in the G0/G1 phase. From the 10th passage, a higher proliferative state was observed, indicating inflammatory hyperactivation, with an increase in the levels of inflammatory cytokines (IL-1, IL-6, and TNFα), nitric oxide, superoxide, lipoperoxidation, protein carbonylation, reactive oxygen species, and DNA damage. The S-HP imbalance affected the intensity of some immunosenescence parameters. AA cells, which present basal high HP levels, were associated with higher DNA damage and lipoperoxidation levels, whereas VV, which present basal high S levels, was associated with higher proinflammatory cytokine levels. In summary, the results suggested that a basal S-HP imbalance could affect the intensity of some immunosenescence markers, and this influence could explain the potential association between an imbalance of genotypes (AA and VV) and the risk of developing some chronic diseases.
Introduction
T
Moreover, aging causes functional deficits in the T and B lymphocyte functions. 3,4 Therefore, these changes lead to the immune system becoming more inclined to elicit nonspecific inflammation, which accelerates degenerative morbidities such as cardiovascular and neurodegenerative diseases. 5 Some reactive molecules, such as nitric oxide (NO) and superoxide (S), which have an importance to trigger phagocytic processes, can be decreased in aged macrophages. 2
Therefore, alterations in the oxidative metabolism have been associated with immunosenescence. In this process, impairment of the antioxidant system, which controls the levels of oxidative molecules, can also occur. This is the case with superoxide dismutase manganese-dependent (SOD2 or MnSOD), which dismutates S in hydrogen peroxide (HP) into mitochondria. Investigations have reported decreases in the levels of SOD2 in aged macrophages. 6 Previous studies suggested that an S-HP imbalance caused by a single-nucleotide polymorphism (SNP) found in the SOD2 gene could alter or change cellular oxidative and inflammatory patterns. In this SNP, a change of valine (GTT) to alanine (GCT) on 16 codon causes a structural protein modification that affects the SOD2 transport into mitochondria and, consequently, the enzyme efficiency. 7 The SOD2 enzyme produced from A allele has an alpha-helix conformation and presents around 40% more efficient SOD2 than V allele, which presents a beta-sheet protein conformation. 8,9
The association between Val16Ala-SOD2 and diseases associated with a chronic inflammatory grade seems to be bidirectional, generally associated with homozygous genotypes that cause both the highest (AA) and lowest (VV) SOD2 efficiency. 10 While the AA genotype has been associated with immunosenescence markers 10 and some cancer types, and the worst infection response causing septic shock, 11 the VV genotype has been associated with cardiovascular dysfunctions, 12,13 metabolic morbidities such as hypercholesterolemia, obesity and diabetes complications, 14 –18 recurrent tonsillitis, 19 and bronchial asthma. 20 Additional investigations also described that VV cells present higher proinflammatory cytokine levels than AV and AA cells. 21 These cells also have a differential response in the inflammatory cytokine levels when exposed to drugs and bioactive molecules, such as methotrexate, 22 rosuvastatin, 23 and resveratrol. 24
It is, therefore, plausible to infer that basal S-HP imbalance Val16Ala-SOD2 SNP associated could have some influence on immunosenescence. Given that it is difficult to investigate this hypothesis in vivo, we evaluated the SOD2 polymorphism influence on immunosenescence indicators of peripheral blood mononuclear cells (PBMCs) chronically in vitro activated by phytohemagglutinin (PHA) exposure.
Methods
General experimental design
The investigation analyzed the influence of a basal S-HP imbalance associated with Val16Ala-SOD2 on PHA (125 μg/mL). This antigen found in plants has carbohydrate binding specificity, and T cell proliferation induced by PHA requires the presence of monocytes. 25 The following immunosenescence indicators were assessed in this study: cellular proliferative rate; apoptosis induction; modulation of NO, superoxide, and cytokine levels (IL-1, IL-6, TNFα, and IL-10); and modulation of oxidative stress biomarkers (lipoperoxidation, protein carbonylation, reactive oxygen species [ROS], and DNA damage).
PBMCs were initially cultured for 72 hours, and the cells obtained were considered 0 passage. These cultures were initially compared among PBMCs carrier's different Val16Ala-SOD2 genotypes and used to obtain aliquots of cells to produce the next culture using the same cellular concentration used in the zero culture (1 × 105 cells), which was again PHA exposed. Every 72 hours, this procedure was repeated until the culture presented a consistent number of immunosenescent markers. Cellular proliferative response to PHA exposure was evaluated in all culture passages, whereas the other markers were sporadically analyzed. In this study, PHA was chosen as the PBMC mitogenic activator since, in contrast to lipopolysaccharide (LPS), which is a major outer surface membrane component of gram-negative bacteria, PHA (lectin) is found in several types of foods habitually consumed by human beings, including peas, peanuts, soybeans, dry beans, broad beans, dry peas, lentils, and others. 26 Therefore, we considered the use of an antigen to be more realistic, as PHA did not trigger acute inflammatory response associated with bacterial infection, which is the case with the LPS molecule.
Ethics issues, Val16Ala-SOD2 genotyping, and blood sample collection
This study is part of a research project previously approved by the Ethics Committee at the Universidade Federal de Santa Maria, Brazil, and all volunteers signed consent terms and agreed to donate blood samples to perform in vitro assays (process number: 0332.0243.000-11). To obtain cells with different Val16Ala-SOD2 genotypes, three subjects' carrier's different genotypes (n total = 12) were chosen from a databank of 120 genotyped healthy volunteers that was established in a previous study conducted by Barbisan et al. 22 The genotyping was performed using a Phusion High-Fidelity PCR kit (Thermo Scientific CO) to access genomic DNA, and the genotype frequencies found in the samples were AA, 26.7%; VV, 28.2%; and AV, 45.1% and were in Hardy–Weinberg equilibrium. The blood samples were obtained from nonsmokers and nonobese subjects with similar health, anthropometric, and lifestyle conditions, including no previous history of any chronic degenerative disease or dysfunction, and no intake of daily medicine. At the moment of blood collection, we certified that the volunteers did not present any acute inflammatory conditions, such as infectious diseases. The volunteers were advised to avoid eating/drinking antioxidant foods such as vegetables and fruits for 24 hours as well as multivitamin supplements to minimize dietary influence on PBMCs. Each volunteer donated 30 mL blood samples, to obtain cell cultures in triplicate.
General cell culture protocol
All reagents and flasks used in the culture were acquired from Vitrocell, Waldkirch and Sigma-Aldrich. To perform the PBMC cultures, the blood samples of 12 subjects (four for each genotype) were collected by venipuncture in heparin tubes (Labttest). PBMCs were obtained within 1 hour of collection using the Histopaque 1077 (Sigma-Aldrich) density gradient medium centrifuged for 15 minutes at 2500 rpm. The cells were counted in a Neubauer chamber using Trypan exclusion dye, and 1 × 105 PBMCs were cultured in the RPMI 1640 culture medium with 10% fetal bovine serum, 1% penicillin/streptomycin, and 1% amphotericin B at 37°C in a humidified 5% CO2 atmosphere for 24 hours. Cultures were also supplemented with PHA (20 μL/mL) (Vitrocell, Waldkirch) to activate PBMCs (macrophages and T lymphocytes). The cell populations obtained from blood samples were identified by flow cytometry analysis, taking into consideration the cell size and granulometry. A new culture passage was produced by centrifugation (20 minutes × 2000 rpm), discarding of supernatant, cell resuspension, counting, and transference by new flask cultures in the same mean culture conditions.
Cell proliferation and viability assays
The biochemical reagents for performing the spectrophotometry and microscopy assays were obtained from Sigma-Aldrich. Kits and reagents for performing flow cytometry were obtained from BD Bioscience. The rate of cell proliferation was evaluated by two assays. The Trypan blue dye exclusion assay was determined by cell microscopic evaluation and colorimetric MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay. The MTT assay quantifies the level of NAD(P)-H-dependent cellular oxidoreductase enzymes, which reflect the number of viable cells. Absorbance was measured at 570 nm. Data were used to compare the proliferative response among cells carrier's different Val16Ala-SOD2 genotypes during each cell culture passage.
To confirm the proliferative response of PBMCs to PHA exposure, the frequency of cells in different cell cycle phases was determined (G0/G1, S, and G2/mitosis phases) by flow cytometry. 27 Cell cycle analysis was similar to that described in Azzolin et al. 28 Briefly, the PI reagent binds to the DNA by intercalating between the bases with little or no sequence preference. PI also binds to RNA, necessitating treatment with nucleases to distinguish between RNA and DNA staining. Following incubation, the cells were trypsinized, washed with PBS, and resuspended in 70% ethanol. Furthermore, cells were stored at −20°C overnight. The cells were centrifuged and washed once with PBS, before being resuspended in 500 μL propidium iodide (PI) solution in PBS: 50 μg/mL PI from 50 × stock solution (2.5 mg/mL), 0.1 mg/mL RNase A, and 0.05% Triton X-100, and incubated for 40 minutes at 37°C. Finally, 1 mL of PBS was added for washing and was resuspended in 500 μL PBS for flow cytometry analysis.
Since the different cell culture passages could trigger apoptosis, this event was analyzed by flow cytometry using an Anexin-V-FITC and PI staining kit (BD Pharmingen™). Anexin-V-FITC can identify an early apoptotic process, whereas PI enables late or necrotic apoptotic cells to be identified. The apoptosis detection is determined through the bond the cell receptors form with the fluorescent antibodies Anexin-V and PI. The assay was performed following the manufacturer's instructions.
Quantification of oxidative markers
Six oxidative markers were quantified in different cell culture passages: NO levels were determined spectrophotometrically by modified Griess method 29 and S levels. 30 Both assays were read at 550 nm. The Levine method 31 read at 370 nm was used to quantify protein carbonylation levels. Lipoperoxidation was estimated by the formation of thiobarbituric acid reactive substances measured by spectrophotometry and read at 532 nm. 32 ROS level was quantified by dichlorofluorescein acetate fluorimetric assay read at an excitation of 485 nm and an emission of 520 nm. 33 DNA damage was determined by 8-deoxiguanosine using an ELISA immunoassay kit obtained from Abcam according to the manufacturer's instructions. All levels of these variables were corrected by mg/protein.
Quantification of cytokines
Cytokine levels were quantified using kit immune assays from a Quantikine Elisa kit obtained from R&D Systems, which is able to quantify cytokines in cell culture supernates. The assays were performed according to the manufacturer's instructions. The sensitivity and detection range of each cytokine were, respectively, as follows: IL-1 (1 pg/mL, 3.1–300 pg/mL); IL-6 (0.7 pg/mL, 3.1–300 pg/mL); TNFα (5.5 pg/mL, 15.6–1000 pg/mL); and IL-10 (3.9 pg/mL, 7.8–500 pg/mL).
Statistical analysis
All measurements of variables analyzed in the study were performed in triplicate. As variance of results could be produced from individual variation or experimental errors, all evaluations were performed using an aliquot sample obtained from cultures from each volunteer with a similar SOD2 genotype. The results were expressed as mean ± standard deviation (SD). One-way analysis of variance or repeated-measures analysis of variance followed by Tukey's post hoc test were used to perform univariate analysis without considering Val16Ala-SOD2 polymorphism. The effect of this polymorphism on immunosenescence patterns was determined by two-way analysis of variance followed by post hoc Bonferroni test. The statistical tests were performed using GraphPad Prism software, and significant differences were considered when p ≤ 0.05.
Results
Flow cytometry analysis confirmed that cell cultures produced from PBMC samples carrier's different Val16Ala-SOD2 genotype consisted of two mononuclear cell populations: macrophages (P8) and lymphocytes (P5) with the presence of a few lymphocyte precursors (P0) in very low concentrations (Fig. 1A). As expected, exposure to PHA antigen caused immune activation observed by an increase in cellular proliferation in 72-hour cell culture (zero passage). A significantly higher cell proliferation was observed in AA than in V cell carriers (VV and AV) (Fig. 1B), indicating that the highly efficient SOD2 enzyme found in AA cells interferes with the speed and intensity of inflammatory response.
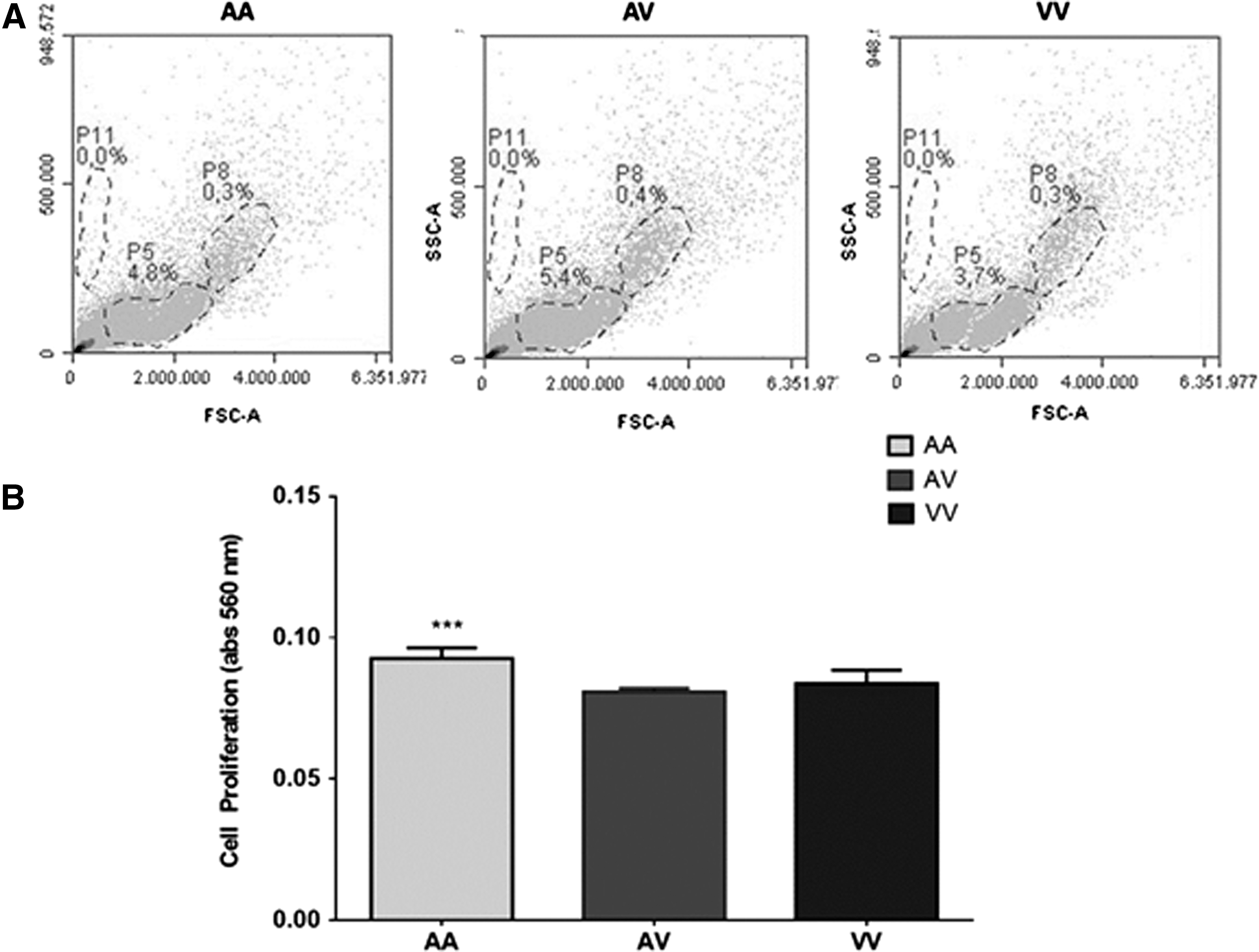
General characteristics of cell cultures produced from healthy adult subjects' carrier's different Val16Ala-SOD2 genotypes, which cause basal S-HP imbalance.
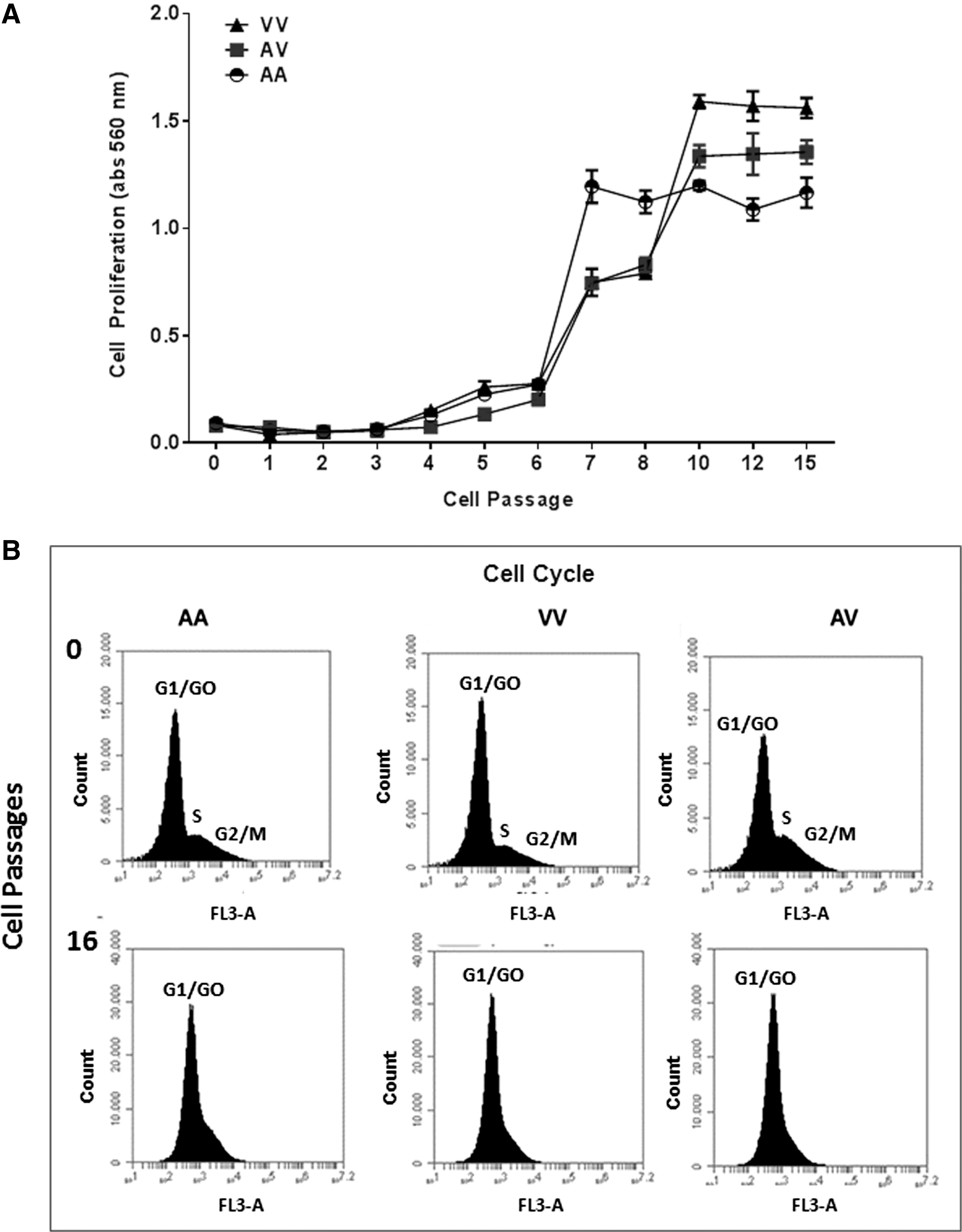
A cell proliferation pattern among different culture passages was initially evaluated without considering the Val16Ala-SOD2 genotype (Fig. 1C). The results showed a similar proliferative rate from zero to the third passage. In culture passages 4–6, the proliferation rate increased substantially in comparison with initial cultures. However, an unexpected finding was observed from 7th cell culture passage, which presented a hyperactivation of cellular proliferation. However, between the 10th and 15th passage, the cell concentrations tend to stabilize despite the cellular concentration being more than 10 times higher than the initial culture passages.
These results were compared among cells carrier's different Val16Ala-SOD2 genotypes (Fig. 2A). Similar proliferative patterns found in zero cell cultures were observed until the 4th passage. However, in the 5th–6th cell culture passages, this pattern changed with a clear increase in cellular proliferation occurring. At this moment, cellular proliferative rates were higher in homozygous (AA and VV) than in heterozygous (AV) cells (p = 0.001).
Comparison of some immunosenescence indicators of cell cultures produced from PBMCs carrier's different Val16Ala-SOD2 genotypes (AA, VV, and AV).
The hyperactivation process was observed in all cultures, but again, AA cells presented a higher proliferative rate than V allele cells. However, in the next passage, AA cells tended to maintain a similar proliferative pattern until the 15th passage, whereas VV and AV cells continued growing until the 10th passage. From this passage, V cells also showed a stabilization in cell proliferation rate until the 15th passage.
From the 10th to 15th culture passages we observed production of synergic lymphocyte-macrophage clusters (rosettes) (Fig. 2B) which made it difficult to continue the experiments. As cell cycle analysis by flow cytometry showed that most cells were in the G0/G1 phase indicating some proliferative senescence state, we opted to finish the analysis in the 15th cell culture passage. All cells presented a similar pattern independent of Val16Ala-SOD2 (Fig. 2C).
During the different cell culture passages analyzed in this study, apoptosis events were not significant. In fact, as can be seen in Figure 3, necrotic and apoptotic events were higher in the initial culture (zero) than in subsequent cell culture passages, probably due to cell manipulation at the moment of collection and processing of blood samples. Although apoptosis events cannot be found in the cultures, the results showed a modification in the cell size and granularity in the last passage evaluated in this study when compared with initial cultures. These results were also observed in all samples indicating no influence of basal S-HP imbalance associated with Val16Ala-SOD2 SNP.
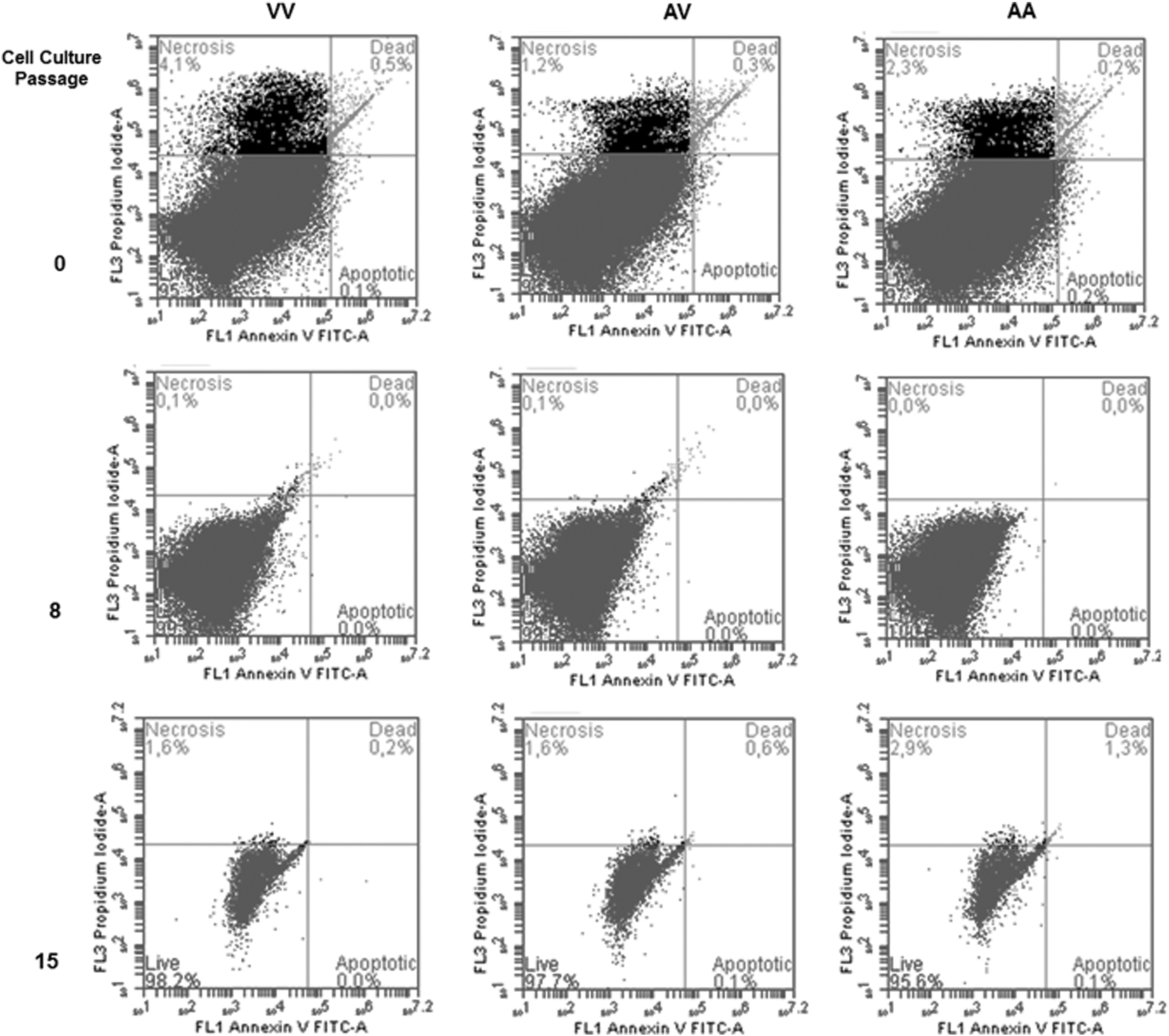
Representative Annexin-PI flow cytometry of apoptosis events in different cell culture passages (0, 8, and 15th) of PBMCs carrier's different Val16Ala-SOD2 genotypes (AA, VV, and AV). Apoptosis events were more frequent in initial culture (0 passage), and changes in cell morphology determined by size and granulometry were observed in 15th cultures. These results were independent of Val16Ala-SOD2 genotypes.
Inflammatory activation by PHA exposure among different cell culture passages was also analyzed by modulation of NO, superoxide, and cytokine (IL-1, IL-6, TNFα, and IL-10) levels.
NO concentrations were significantly lower in AV than in AA and VV cells in the zero passage (p ≤ 0.001). From the first to the 10th passage a slight increase in the levels of this oxidative marker was observed. Between the 10th and 15th passage, a significant increase in NO concentrations occurred (p ≤ 0.001). These results were independent of Val16Ala-SOD2 SNP. S concentrations tended to increase linearly in relation to cell culture passages. From the first cell culture passage, the levels of all proinflammatory cytokines were significantly increased and continuously rose until the 15th passage. The only difference was that VV cells presented higher levels, AV- intermediary levels, and AA lower levels of proinflammatory cytokines. The concentration of IL-10, an anti-inflammatory cytokine, increased slightly among successive cell culture passages in all cells. However, higher IL-10 levels were observed in AA cells, whereas VV cells presented lower levels of this anti-inflammatory molecule. Again, AV cells presented intermediary levels in relation to homozygous cells (Fig. 4).

Comparison of inflammatory indicators among cell culture passages obtained from PBMCs carrier's different Val16Ala-SOD2 genotypes. All variables were corrected by mg/protein. Statistical comparisons were performed by two-way analysis of variance followed by Bonferroni post hoc test and are commented in the text. Different letters (a, b, c, and d) were used to identify statistical differences (p ≤ 0.05) among different cellular passages of each cell culture genotype. IL-1, interleukin 1; IL-6, interleukin 6; TNFα, tumor necrosis factor alpha; IL-10, interleukin 10.
Oxidative markers related to inflammatory response and oxidative stress were evaluated among cell culture passages obtained from PBMCs carrier's different SOD2 genotypes (Fig. 5). From the first culture passage, the ROS levels were increased and continuously rose in subsequent cultures. Similar lipoperoxidation concentrations were observed from zero to the 5th cell culture passage. From this passage, the levels of this oxidative stress marker started to rise. In the last cell culture passage analyzed in this study, AA cells presented higher lipoperoxidation levels than V allele cells (AV and VV). Protein carbonylation also presented variation in its concentration mainly between the 5th and 10th cell culture passages. However, VV cells showed higher protein carbonylation levels than others. DNA damage also changed the levels in relation to cell culture passage. The 8-deoxyguanosine levels began to rise from the first cell culture passage and tended to maintain the same concentration in further passages. DNA damage was higher in AA than in VV cells. AV cells presented intermediary values when compared with homozygous cells for all oxidative stress markers analyzed in this study.
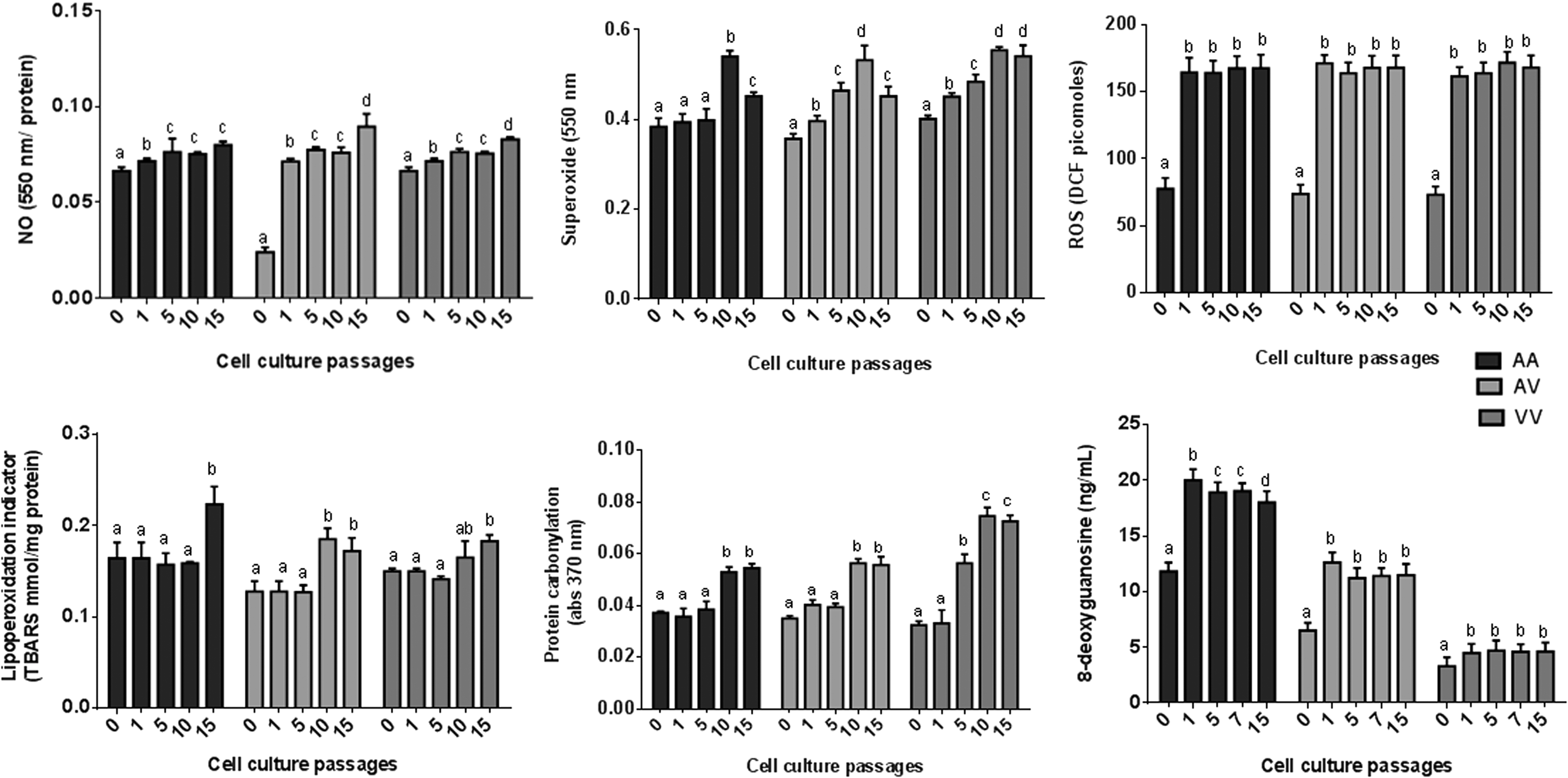
Comparison oxidative markers among cell culture passages obtained from PBMCs carrier's different Val16Ala-SOD2 genotypes. Statistical comparisons were performed by two-way analysis of variance followed by Bonferroni post hoc test and are commented in the text. Different letters (a, b, c, and d) were used to identify statistical differences (p ≤ 0.05) among different cellular passages of each cell culture genotype. NO, nitric oxide; S, superoxide; ROS, reactive oxygen.
Discussion
This study evaluated the impact of genetic S-HP imbalance due to changes in the efficiency of the SOD2 enzyme, which has been associated with several chronic age-related diseases 8 on in vitro modulation of in vitro PBMC immunosenescence. We will discuss the results obtained considering three aspects: general immunosenescence results observed in all cultures, differential results observed in PBMCs carrier's different Val16Ala-SOD2 genotypes, and the impact of these results on potential postponed aging interventions.
When PBMCs were chronically PHA activated, the following common immunosenescence patterns were observed in all cultures: higher cell proliferation rates; changes in cellular morphology; and elevation in the levels of proinflammatory cytokines and oxidative stress markers in the different cell culture passages. In contrast, S levels decreased in the older cultures when compared to younger cultures. In the 15th passage, cell cycle analysis showed arrestment in the G0/G1 phase of cells suggesting a proliferative senescence pattern. Rosette production by the cultures began to make it difficult to obtain aliquots to produce the next culture, so we resolved the issue by finishing the cell culture experiment with this passage.
When the study was conceived, we expected a shorter extension of PBMC proliferation cultures than observed in nonactivated PBMCs, which is generally maintained in primary culture conditions for approximately 15 days. Starting from 15 days of culture, a decrease in cellular proliferation was observed, indicative of an immunosenescence process. 24 Several investigations in humans and rats describe a decrease in PHA- or LPS-stimulated lymphocyte proliferation with age. 34,35 Surprisingly, PBMCs continually exposed to PHA antigen presented a proliferative hyperactivation process from intermediary and older cultures. Therefore, explaining these results could be considered a challenge.
In fact, one possible explanation of hyperactivation occurrence, as observed in this study, could be a process of promiscuous macrophage proliferation, which is observed in the presence of a subclinical chronic inflammatory process that is very common in elderly people. 1,36 Given that the initial percentage of macrophage in our cell cultures was similar among cells carrying different Val16Ala-SOD2 genotypes, it would be very hard to accept this hypothesis. However, we do not know whether the proportion of macrophages and lymphocyte cells changed along the different cell culture passages. Unfortunately, it was not possible to follow the cultures to evaluate this aspect. Despite this limitation, it is conceivable that these differences could be related to changes in the proportion of macrophages, T and B lymphocytes, and NK cells in intermediary and older cultures. However, this hypothesis deserves to be tested in complementary studies. Independent of changes in the specific PBMC proportions, the hyperactivation observed in our cultures could be considered an immunosenescence process.
As the hyperactivation process was influenced by Val16Ala-SOD2 SNP, our results also suggested that this process is modulated, at some level, by oxidative metabolism, and are according to De la Fuente's suggestion about the occurrence of involvement of oxidative changes of the immune system in aging processes. 37 In relation to the SNP studied in this study, some studies partially analyzed this question. For example, Isbir et al. 38 investigated the effect of Val17Ala-SOD2 SNP on the proinflammatory cytokine levels of patients undergoing coronary artery bypass grafting with a cardiopulmonary bypass. Although the authors did not show differences in the baseline IL-6 levels in patients carrying different SOD2 genotypes, postoperative analysis showed that V patients (VV and AV) presented higher IL levels than AA patients. Therefore, this study demonstrated previously that Val16Ala-SOD2 SNP could influence inflammatory modulation, which includes positive modulation of IL-1 proinflammatory cytokines. Paludo et al. 39 described Val16Ala-SOD2 SNP effects on the production of intracellular ROS by PBMCs with and without induction of LPSs. These studies, performed by independent research groups, reinforce the suggestion that REDOX imbalance, especially involving the S and HP levels, could have some repercussions on inflammatory response. 35
The emergence of rosette formation was another result observed in the last five-cell culture passages. Multicellular rosettes are structures formed in embryonic development and nonembryonic development contexts. Multiple modes of rosette formation have been described in adult tissues or in vitro conditions. In blood, rosettes can be formed by cell–cell interactions, mainly between macrophages and red blood cells or macrophages and T lymphocytes. 40,41 As in embryonic development, the biological relevance of rosette formation is well determined; in adult tissues, the biological role of rosettes is not very clear. In relation to immune cells, rosettes could present a mechanism related to control damaged or transformed cells into body. This hypothesis was studied by Hicks et al., 42 evaluating the action of macrophages obtained from mice with spontaneous regression of multiple types of cancer (SR/CR). The authors observed that SR/CR macrophages did not use the classical mechanism to kill tumoral cells, as increase of reactive nitrogen species and ROS production. These immune cells form rosettes with cancer cells, to unleash their cytotoxic mechanisms, which induce apoptosis. Although this was not our focus, we could speculate that rosette formation in older cultures could be a protective mechanism against some cellular internal or external age-associated stressors. As we are constantly changing the culture medium, some cell damage is expected, and accumulation of stressful molecules could cause macrophage and/or T lymphocyte membrane modifications triggering the formation of rosettes. However, this hypothesis will need to be tested by future complementary investigations.
From these results, we believed that PBMCs exhibited an immunosenescence pattern when submitted chronically to PHA exposure in several cell culture passages. The results also showed that the general pattern of immunosenescence was not changed by a basal S-HP imbalance associated with Val16Ala-SOD2 SNP, that is, proliferative hyperactivation, rosette formation, or an increase in the levels of oxidative and proinflammatory cytokines during cell culture passages. However, an S-HP imbalance had an influence on the speed and intensity of some immunosenescence markers, mainly at the levels of proinflammatory cytokine and oxidative stress markers. These differential responses observed in this study could be influenced by the occurrence of variations in the oxidative and inflammatory levels in nonactivated PBMCs, as described in previous studies. 14,16
In a 72-hour cell culture (zero passage), AA cells, which have a more highly efficient SOD2 enzyme, presented a faster and more intense immune response, whereas VV cells, which have lower SOD2 efficiency, presented a slower immune response. Similar results have been described in previous studies. 22,36,43 Although AA cells are more responsive to PHA activation, VV cells maintained higher cytokine levels in all the cell culture passages analyzed in this study. In vitro and in vivo investigations have reported an association between VV genotypes and high inflammatory cytokine levels. 22 –25 Moreover, differential inflammatory modulation by drugs has been reported from in vitro and in vivo studies suggesting that VV cells could be more resistant to decreases in the levels of proinflammatory and oxidative markers. 20,22 For example, Duarte et al. 23 evaluated the effect of rosuvastatin treatment (20 mg/day) in 120 hypercholesterolemics carrier's different Val16Ala-SOD2 genotypes. Rosuvastatin is a hypolipimiant drug with well-known anti-inflammatory properties. After 120 days, the authors observed that VV patients presented some resistance to lowering proinflammatory cytokine levels when compared with patients' carrier's A allele genotypes (AA and AV).
In contrast, higher oxidative stress markers were observed in AA PBMCs during cell culture passages, including lipoperoxidation and DNA damage, than in V allele PBMCs. This association is probably due to the higher basal HP levels produced from the higher SOD2 efficiency found in AA cells. As HP is membrane soluble, excess concentration of this molecule in mitochondria could migrate to cytoplasm reacting to metal ions, such as iron and zinc (Fenton reaction), which produce hydroxyl radicals and other prooxidant molecules. Hydroxyl radicals present a special high affinity with DNA molecules causing broken and chromosomal instability. Probably, the higher production of hydroxyl radicals is one of the causal mechanisms related to a higher risk of AA subjects developing some cancer types, such as prostate, breast and lung cancer. 9,10 Therefore, our results are in line with previous investigations that associated higher DNA damage with AA cells. 10,11,37
In the inflammation context, an in vitro investigation showed that AA cells that LPS activated presented a higher prooxidant state than VV cells. 44 The authors considered these results relevant in patients who have developed septicemia, which is a complication related to infectious diseases. In fact, AA patients present a high risk of developing septic shock and probably this risk is due to the overproduction of ROS and reactive nitrosative species associated with an intense inflammatory response. 12
Based on these results, it is also relevant to perform some methodological considerations about the choice of whether to perform the experiments using PBMCs and not polymorphonuclear cells, which are also involved in the inflammatory metabolism. We chose PBMCs because these cells, when exposed to antigen, triggered an inflammatory cascade. In tissues, generally, resident macrophages (M1) are stimulated by antigens or injuries inducing inflammatory response by increasing proinflammatory cytokine levels, such as IL-1, IL-6, and TNFa, as studied in this study. These processes induce the migration of other leukocytes, especially polymorphonuclear cell neutrophils, to the inflammatory site. 3 Under these terms, we opted to evaluate the influence of the S-HP imbalance on a chronic inflammatory stimulation, which is a process closely associated with aging biology that mainly involves mononuclear cells, such as macrophages. Another important question related to our experimental design regard the use of PBMCs obtained from younger healthy subjects. We chose young human PBMCs because it is very difficult to obtain PBMCs from older people with some prior homeostasis alteration. We based our choice on previous studies that showed important alterations in elderly people with healthy immune conditions compared with young adults. 45,46 A study observed higher cortisol levels in the elderly than in younger subjects, indicating an activation of the hypothalamic-pituitary-adrenal axis. Cells obtained from elderly people also showed reduced counts of CD45RCD95+ cells than younger cells. In fact, it is very hard to find immune healthy older subjects who do not present physiological alterations or have previous chronic diseases associated with chronic inflammation, and/or do not use daily medicine that could affect the experimental analysis. For example, in the studies previously cited, 41,42 the authors found just 46 noninstitutionalized healthy elderly people out of 1118 subjects. These subjects were selected in accordance with the SENIEUR protocol that defines rigorous criteria for selecting strictly healthy individuals for immunogerontological studies. Moreover, in vitro aging studies, such as that performed by Pink et al., 47 suggested that monocytes from elderly people present a higher basal production of proinflammatory cytokines, such as TNFa. Therefore, based on this evidence, we opted to evaluate the potential of in vitro senescence of PBMCs obtained just from younger subjects. Finally, another limitation in our investigation is related to the lack of analysis of antioxidant markers. In this study, enzymatic and nonenzymatic antioxidant molecules were not evaluated by logistic questions involving a limited sample volume and complex processing of the samples to perform these analyses throughout cell cultures. Therefore, complementary studies need to be performed to evaluate the role of antioxidant molecules other than SOD2 from the results obtained in this study.
Moreover, despite methodological concerns related to in vitro investigations, all the results suggested that a basal S-HP imbalance had an impact on the modulation of some aspects of immunosenescence, although the general pattern of this immune aging process triggered by PHA activation was conservative.
The two imbalanced SOD2 genotypes seem to have a negative impact on immune response cell carriers, whereas the high-SOD2 efficiency genotype (AA) produced higher levels of oxidative stress markers and the low-SOD2 efficiency genotype (VV) produced higher proinflammatory cytokine levels than AV cells. S-HP imbalance found in both homozygous genotypes has been associated with diseases related to chronic inflammatory patterns, such as cancer, and to cardiovascular and metabolic disorders. 8
The role of an S-HP imbalance in proliferative cytokines and oxidative molecules from PBMC culture passages could be related to the role of S and HP in inflammatory modulation. A large number of studies have suggested the relevance of oxidative metabolism in inflammatory response. First, tissue injury leads to the production of ROS, including S and HP molecules generating oxidative stress. 48 Second, the elevation of some types of ROS molecules has also been associated with chronic regeneration of wounds. This is the case with HP molecules, whose rapid production in response to wounding is considered essential in minimizing infection risk, and in activating the epithelial signaling pathways necessary for triggering epidermal regeneration. Moreover, the increase in HP levels has a role in recruiting neutrophils and other immune cells to the wound site. 49 On the other hand, phagocytes also generate ROS molecules. Macrophages, in particular, produce high levels of NO associated with its phagocytic activity. As is well known, at low concentrations, NO is considered an important signaling molecule. In contrast, high NO concentrations produced by macrophages at the injury site interfere directly, since it is highly reactive with S, and this reaction generates peroxynitrite, which can cause extensive membrane lipoperoxidation. 50 As AA causes increases in basal HP levels, whereas VV causes increases in S basal levels, this condition could potentially affect the inflammatory response of PBMCs continually exposed to PHA. Although the mechanisms related to the S-HP imbalance in chronic inflammatory induction need to be clarified, the results described in this study corroborate the theory, which preconizes the involvement of the immune system in the aging process of the organism by alterations in the oxi-inflammatory balance. 49
Finally, it is important to discuss the potential relevance of our findings for preventing chronic morbidities associated with an oxi-inflammatory imbalance and postponing aging processes associated with body dysfunctions. As postulated previously by De la Fuente and Miguel, 51 the confirmation of the central role of oxi-inflamm-aging could be through the administration of adequate dietary richest in antioxidant molecules. 21 It is important to point out that several foods that present antioxidant properties also have anti-inflammatory effects [x]. Previous results obtained from Val16Ala-SOD2 epidemiological studies could also be used to corroborate indirectly the oxi-inflamm-aging theory. This is the case with investigations involving cancer risk with Val16Ala-SOD2 and dietary pattern interactions. 52 –54
Although an in vitro investigation performed by Capeleto et al. 24 suggested that the interaction between the genetic imbalance caused by Val16Ala-SOD2 SNP and resveratrol, a powerful molecule associated with lifespan extension, could modulate differentially inflammatory patterns; strategies aimed at improving the redox state of the immune system 55 could be considered essential for minimizing degenerative aging processes.
Footnotes
Acknowledgments
The study described was supported by grants and fellowships from Brazilian research agencies (CNPq and CAPES). We wish to express our thanks to Beatriz da Rosa Bonadiman, Charles Elias Assmann, Dianni Capeleto, and Cibele Ferreira Teixeira for their help with cell culture management. F.B., V.F.A., and M.M.M.F.D.: cell culture, analysis, and data interpretation. F.B., E.E.R., and I.B.M.C.: study concept, design, data interpretation, and article writing.
Author Disclosure Statement
No competing financial interests exist.