
Abstract
Erythrocytes are particularly vulnerable toward age-dependent oxidative stress-mediated damage. Caloric restriction mimetics (CRMs) may provide a novel strategy for the maintenance of redox balance as well as effective treatment of age-associated diseases. Herein, we have investigated the beneficial effect of cotreatment with CRM-candidate drugs, rapamycin (an immunosuppressant drug and inhibitor of mammalian target of rapamycin) and metformin (an antidiabetic biguanide and activator of adenosine monophosphate kinase), against aging-induced oxidative stress in erythrocytes and plasma of aging rats. Male Wistar rats of age 4 (young) and 24 months (old) were coexposed to rapamycin (0.5 mg/kg body weight [b.w.]) and metformin (300 mg/kg b.w.), and data were compared with the response of rats receiving an independent exposure to these chemicals at similar doses. The exposure of individual candidate drugs significantly reversed the age-dependent alterations in the endpoints associated with oxidative stress such as reactive oxygen species, ferric reducing ability of plasma, malondialdehyde, reduced glutathione, plasma membrane redox system, plasma protein carbonyl, and acetyl cholinesterase in erythrocytes and plasma of aging rats. However, the cotreatment with rapamycin and metformin showed a significant augmented effect compared with individual drug interventions on reversal of these age-dependent biomarkers of oxidative stress, suggesting a synergistic response. Thus, the findings open up further possibilities for the design of new combinatorial therapies to prevent oxidative stress- and age-associated health problems.
Introduction
A
The erythrocytes are continuously exposed to endogenous and exogenous reactive oxygen species (ROS) under normal physiological conditions as the erythrocyte membrane is permeable to numerous oxygenated free radicals. 6 In addition, erythrocytes are fully equipped with antioxidant defense system responsible for their possible protection to a certain extent against oxidative damage. 7 Therefore, the erythrocyte represents an appropriate cellular model to study the mechanism(s) of aging and redox modulation. 8,9 Additionally, the lack of nucleus and other cell organelles in erythrocytes instinctively eases protein complexity and makes them a wonderful tool for biochemical studies. 10
The therapeutic targeting against oxidative stress may therefore provide an innovative strategy for the effective treatment of age-associated diseases. Recently, caloric restriction (CR) has emerged as a successful and consistent nonpharmacological intervention for protection against the age-associated deterioration of biological functions and improving overall health and life expectancy in several model organisms. 11 –14 Conversely, the implementation of CR in humans is difficult to adhere as a long-term strategy because of ethical issues. Thus, there is an increasing interest for search of candidate drugs that are able to mimic and elicit the same beneficial effects of CR on health, life span, and aging. Such candidate drugs, the CR mimetics (CRMs), actually share the same metabolic pathway(s) and stress responses as CR. 15
Rapamycin is categorized as a potential CRM candidate that has been suggested to be among the most promising antiaging drugs, affecting all age-related diseases and promoting healthy aging. 16 The age-delaying effect of rapamycin is mediated by inhibition of mammalian target of rapamycin (mTOR) activity and subsequent activation of autophagy. 17 Thus, rapamycin plays an important role in mTOR-dependent regulation of several cellular functions, including cell growth, protein synthesis, redox homeostasis, and autophagy during aging. 18,19 Rapamycin has been reported to bind efficiently to FK-binding proteins of erythrocytes and is highly distributed into erythrocytes. 20
Metformin, an antidiabetic biguanide, is also considered as a CRM candidate having efficient antiaging effects. 21 It acts through induction of adenosine monophosphate-activated protein kinase (AMPK)-dependent autophagy to control the aging process and extend life span. 22,23 Metformin influences metabolic and cellular processes closely associated with the development of age-related conditions such as inflammation, 24 autophagy, 25,26 and cellular senescence 27,28 and thus extends the life span of the organism. Thus, the present study was designed to examine the combinatorial effect of rapamycin and metformin against age-dependent oxidative stress in erythrocytes and plasma of rats.
Materials and Methods
Chemicals and drugs
All the chemicals and reagents used for this study were purchased from Sigma Aldrich (St. Louis, MO) unless otherwise stated. Other chemicals such as solvents were of analytical grade and procured from Merck (Germany). Rapamycin was purchased from TCI Chemicals (India) Pvt., Ltd. (India). Metformin tablets were procured from local pharmaceutical stores. Autoclaved deionized water was used in all the experiments.
Animals
The experiment was carried out with male Wistar rats of two different age groups, viz, 4 months (young) and 24 months (old), weighing 130 ± 30 g and 300 ± 30 g, respectively. Rats were bred and maintained in the animal house of our department. They were housed under good sanitary conditions and controlled environment (22°C ± 2°C and relative humidity 55% ± 15%) with a 12-h light/12-h dark cycle. All rats were fed with a normal laboratory diet of nutrient-rich pellets containing total energy as fat, protein, and carbohydrate (Pranav Agro Industries Ltd., Pune, India) and had free access to drinking water. The protocols and procedures of the study were approved by the Animal Care and Ethics Committee of University of Allahabad (Allahabad, India).
Experimental design and treatment protocol
Rats were randomly divided into following eight groups having six animals in each group.
(1) Young control: Received daily single oral administration of vehicle solution (0.9% NaCl solution containing 0.5% DMSO) for 21 days.
(2) Young rapamycin: Received daily single oral administration of rapamycin (0.5 mg/kg body weight [b.w.]) for 21 days.
(3) Young metformin: Received daily single oral administration of metformin (300 mg/kg b.w.) for 21 days.
(4) Young rapamycin + metformin: Received daily single oral administration of rapamycin (0.5 mg/kg b.w.) and metformin (300 mg/kg b.w.) for 21 days.
(5) Old control: Received daily single oral administration of vehicle solution (0.9% NaCl solution containing 0.5% DMSO) for 21 days.
(6) Old rapamycin: Received daily single oral administration of rapamycin (0.5 mg/kg b.w.) for 21 days.
(7) Old metformin: Received daily single oral administration of metformin (300 mg/kg b.w.) for 21 days.
(8) Old rapamycin + metformin: Received daily single oral administration of rapamycin (0.5 mg/kg b.w.) and metformin (300 mg/kg b.w.) for 21 days.
The route of administration and dose of rapamycin (0.5 mg/kg b.w.) were selected from our earlier report 29 and the dose of metformin (300 mg/kg b.w.) was selected from elsewhere. 30
Blood collection and separation of packed red blood cells
After completion of treatment schedule, rats were anesthetized with ketamine hydrochloride (100 mg/kg, i.p.). Blood was drawn by cardiac puncture into heparinized (10 U/mL) syringes and subjected to centrifugation at 800 g for 10 minutes at 4°C. The red blood cells (RBCs) were washed twice with cold phosphate-buffered saline (PBS) at physiological pH after removal of plasma, buffy coat, and upper 15% of the cells. The remaining packed RBCs (PRBCs) were resuspended in PBS containing 0.09% glucose. The protein content in blood plasma was measured following the method of Lowry et al. 31 and plasma was stored at −80°C for further experimentation.
Measurement of ROS
To determine the effect of rapamycin, metformin and their cotreatment on oxidative stress, ROS was measured using 2′, 7′-dichlorodihydrofluorescein diacetate (DCFH-DA) dye following the protocol earlier described by us. 21 In brief, PRBCs (100 μL) of entire experimental groups were washed in PBS and stained with DCFH-DA (20 μM) dye for 30 minutes at 37°C in the dark. Then, the stained PRBCs were washed thrice with PBS and finally resuspended in 1.0 mL PBS, and fluorescence intensity was measured using an excitation/emission wavelength of 485/528 nm, respectively, using Cary eclipse fluorescence spectrophotometer (Agilent Technologies). The results of ROS generation are expressed in percent change with respect to young control group that was set at 100%.
Furthermore, intracellular ROS generation in PRBCs was also measured using a fluorescence microscope. In brief, PRBCs (100 μL) were diluted at a ratio of 1:1 with PBS containing 0.09% glucose and incubated with DCFH-DA dye (20 μM) for 30 minutes at 37°C in the dark. The stained PRBCs were then washed with PBS, smeared and mounted on slides for microscopic observation. Images were captured by using an Olympus fluorescence microscope (Model: CX21i-TR-LED) equipped with Magnus UHCCD-USB 21.4 megapixel camera.
Measurement of total antioxidant activity by ferric reducing ability of plasma assay
The total antioxidant potential of the plasma was determined using well-known endpoint ferric reducing ability of plasma (FRAP) assay. 32 The FRAP reagent was prepared by mixing of 300 mM acetate buffer (pH 3.6), 20 mM ferric chloride, and 10 mM 2,4,6-tripyridyl-s-triazine in 40 mM hydrochloric acid at the ratio of 10:1:1, respectively. The FRAP reagent (3.0 mL) was added to 100 μL of plasma and the contents were mixed thoroughly. Then, the absorbance was read at 593 nm at the interval of 30 seconds for 4 minutes. FRAP value (μmol Fe (II)/L of plasma) was calculated using the standard calibration curve.
Measurement of lipid peroxidation
The erythrocyte malondialdehyde (MDA), an index of lipid peroxidation (LPO), was measured following the protocol of Esterbauer and Cheeseman. 33 In brief, PRBCs (0.2 mL) from the entire experimental groups were suspended in 3.0 mL PBS containing 0.5 mM glucose. The PRBC suspension (0.2 mL) was added to 1.0 mL of 10% trichloroacetic acid and 2.0 mL of 0.67% thiobarbituric acid. Then, the reaction mixture was boiled for 20 minutes at a temperature of more than 90°C, followed by subsequent cooling and centrifugation at 1000 g for 5 minutes. Further, the absorbance of mixture was read at 532 nm using Genesys 10VIS spectrophotometer (Thermo Scientific) and the concentration of MDA was calculated using extinction coefficient (ɛ = 153,000 M−1 cm−1). The value of MDA is expressed as nmol/mL PRBCs.
Determination of erythrocyte reduced glutathione
The erythrocyte reduced glutathione (GSH) was measured following the standard protocol described earlier. 34 The method was based on the ability of the -SH groups to reduce 5, 5′-dithiobis, 2-nitrobenzoic acid (DTNB) into a yellow-colored product whose absorbance was measured at 412 nm. The concentration of GSH is expressed in mg/mL PRBCs and was calculated from standard plot.
Measurement of erythrocyte plasma membrane redox system activity
The activity of the erythrocyte plasma membrane redox system (PMRS) was measured by reduction of ferricyanide as described earlier by us. 35 Briefly, PRBCs (0.2 mL) from all experimental groups were suspended in PBS containing 5 mM glucose and 1 mM freshly prepared potassium ferricyanide to a final volume of 2.0 mL. The suspensions were incubated for 30 minutes at 37°C and then centrifuged at 800 g for 10 minutes at 4°C. Then, the ferrocyanide content in supernatant was measured by using 4,7-diphenyl-1,10-phenanthroline disulfonic acid disodium salt and taking absorbance at 535 nm (ɛ = 20,500 M−1 cm−1). The values are expressed in μmol ferrocyanide/mL PRBCs/30 minutes.
Determination of plasma protein carbonyl group
The plasma protein carbonyls (PCOs) were measured following the standard procedure of Levine et al. 36 and our earlier described protocol. 21 The carbonyl content was calculated using an absorption coefficient of 22,000 M−1 cm−1 and the result is expressed as nmol/mg protein.
Measurement of acetyl cholinesterase activity
Acetyl cholinesterase (AChE) activity was assayed in erythrocytes following the earlier described method 37 and also our reported protocol. 5 The hemoglobin (Hb) was estimated by the ferricyanide/cyanide method. 34 AChE activity is expressed in IU (1 IU = μmol of acetylthiocholine iodide hydrolyzed/min) per g of Hb at 37°C. A molar absorption coefficient of 13,600 M−1 cm−1 was used for the thionitrobenzoate ion at 412 nm.
Statistical analysis
The results are expressed as mean ± standard deviation of three independent experiments. Statistical analysis was done by the software GraphPad Prism 5, version 5.01. The statistical significance of difference among various groups was calculated by two-way analysis of variance, followed by post hoc Bonferroni's test. The data with p < 0.05 were considered as significant.
Results
Effect of rapamycin, metformin, and cotreatment on ROS production
To assess the protective effect of rapamycin and metformin cotreatment in erythrocytes of old aged rats, ROS was quantified using DCFH-DA dye. Statistically, significant (p < 0.05) induction in ROS production (184.23% ± 3.52%) was observed in old aged rats when compared with young control. The treatment with rapamycin slightly induced ROS production (109.13% ± 10.53%); however, metformin treatment reduced the generation of ROS (80.4% ± 6.14%) significantly in young rats. The slight induction in ROS generation in young rats following the exposure of rapamycin might be due to intracellular self-defense mechanism. In addition, the alteration in ROS generation was found to be nonsignificant (98.16% ± 6.23%) during cotreatment with rapamycin and metformin in young rats. In contrast, the significant reduction in age-dependent increased ROS was measured following the individual treatment with rapamycin (152.78% ± 9.45%) and metformin (143.7% ± 8.3%). Moreover, the effect of metformin on ROS production was more significant than rapamycin. In addition, the effect of cotreatment with rapamycin and metformin was the most profound on the production of ROS (116.68% ± 6.39%) in old aged rats that brought the level of ROS closer to the young control (Fig. 1A). The fluorescence microscopic image analysis for intracellular ROS generation using DCFH-DA dye maintained the linearity with spectrofluorimetric data (Fig. 1B).
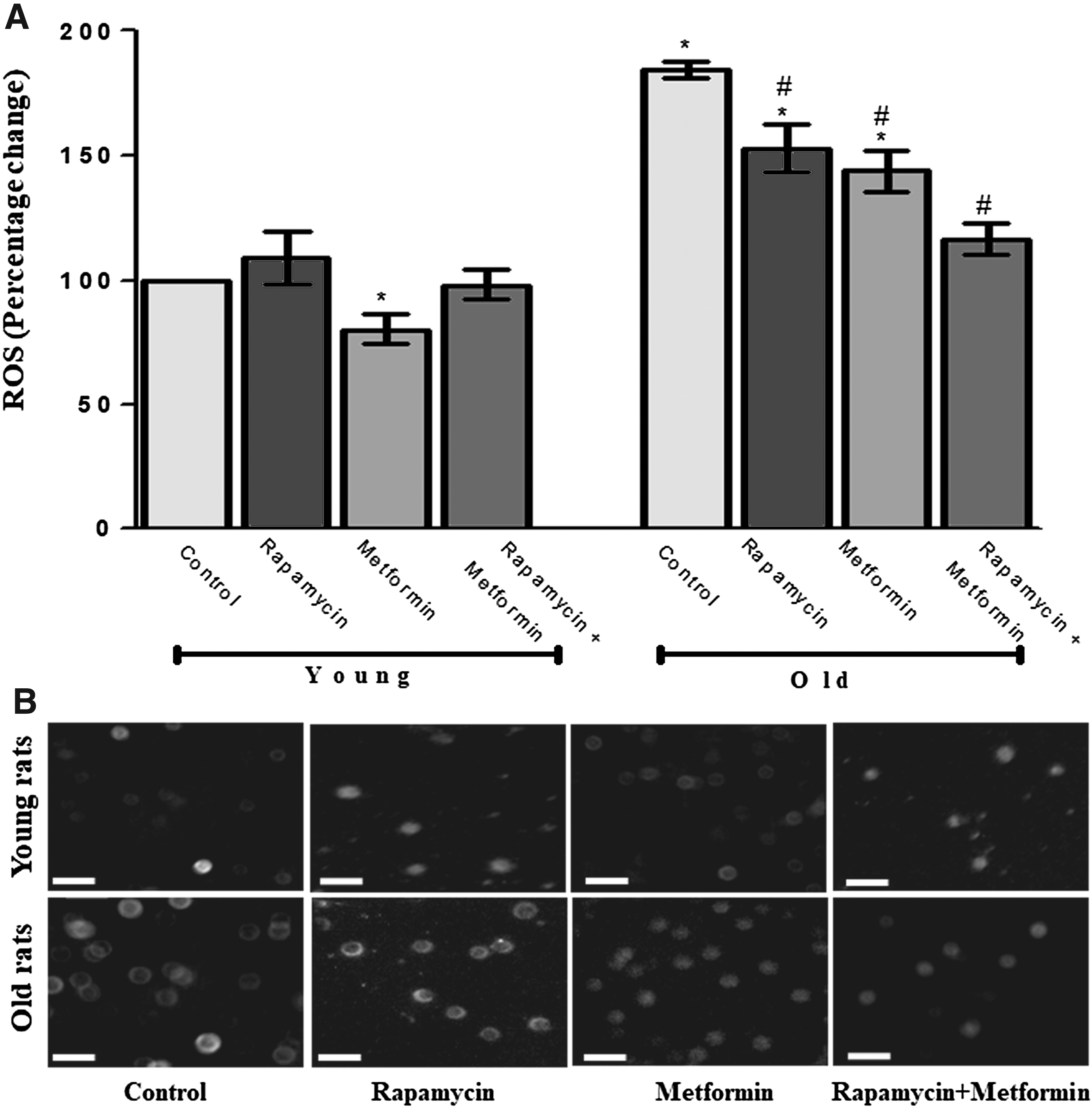
Protective effect of rapamycin, metformin, and their cotreatment on reactive oxygen species (ROS) generation in erythrocytes of young and old aged Wistar rats.
Effect of rapamycin, metformin, and cotreatment on FRAP
Next, we evaluated the protective effects of rapamycin, metformin, and their cotreatment on the total antioxidant potential of the plasma using FRAP (Fig. 2). The FRAP activity was found to be significantly (p < 0.05) reduced (20.3 ± 1.3 mol Fe(II)/L plasma) in old aged rats when compared with the young control group (30.19 ± 1.2 mol Fe(II)/L plasma). The treatment with rapamycin, metformin, and their cotreatment showed a significant protective effect by further elevating the age-dependent decreased activity of FRAP to 31.7 ± 2.7, 41.1 ± 5.4, and 45.4 ± 3.8 mol Fe(II)/L plasma, respectively. Moreover, the cotreatment group showed more pronounced restorative effect in comparison with rapamycin and metformin-treated groups in old aged rats.
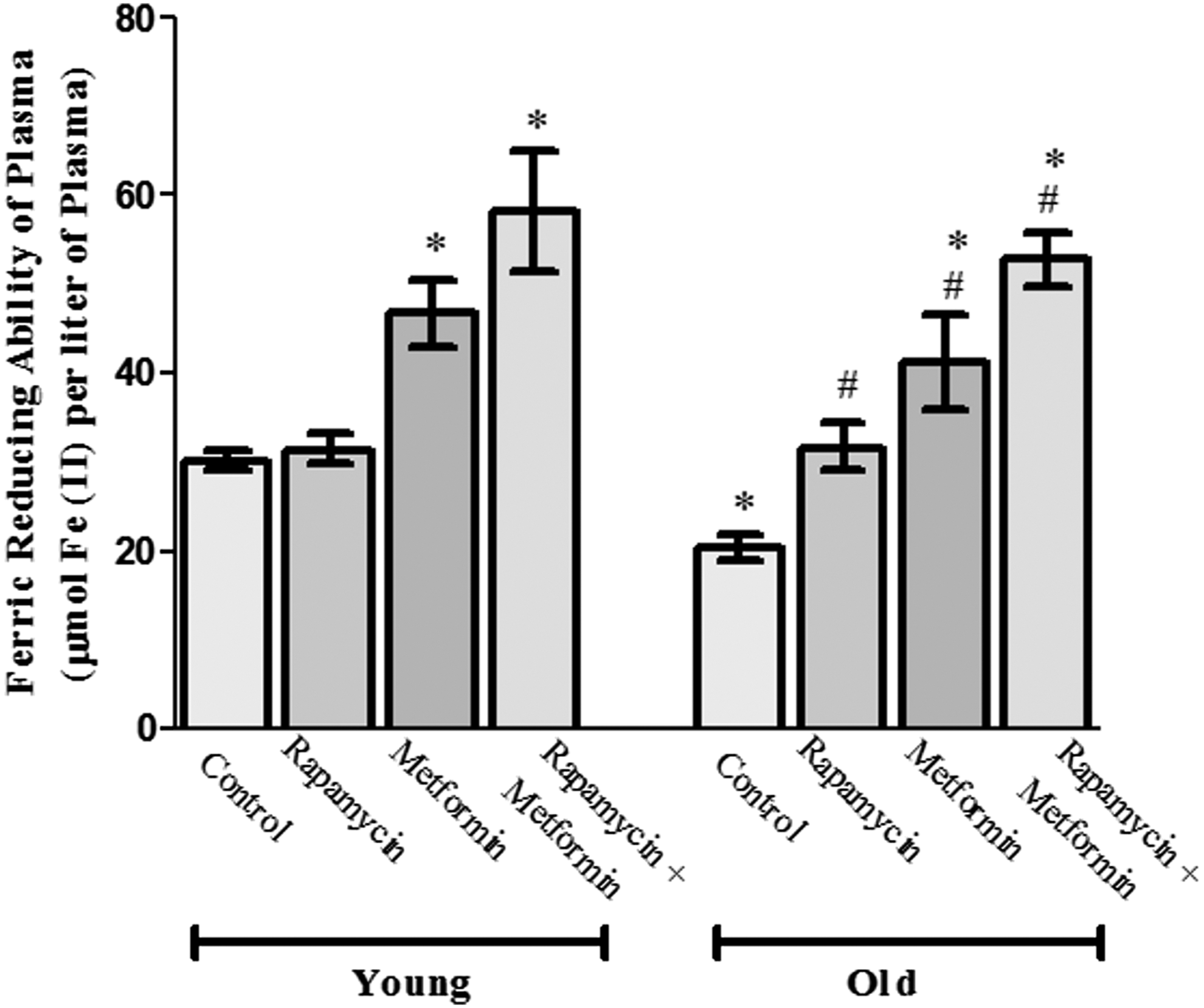
Protective effect of rapamycin, metformin, and their cotreatment on ferric reducing ability of plasma (FRAP) in young and old aged Wistar rats. Values are mean ± SD of three independent experiments and FRAP value is expressed as μmol Fe (II)/L plasma. *p < 0.05 when compared with normal young control rats and # p < 0.05 when compared with old aged control rats.
Effect of rapamycin, metformin, and cotreatment on LPO
The result of LPO in terms of its by-product (MDA) is presented in Figure 3. In the young rat group, the treatment with rapamycin showed nonsignificant response on the level of LPO when compared with respective control (102.14 ± 5.5 nmol/mL PRBCs). However, the treatment with metformin alone and cotreatment with metformin and rapamycin decreased the level of LPO significantly (p < 0.05) to 84.82 ± 3.57 and 72.1 ± 4.87 nmol/mL PRBCs, respectively, in young rats compared with the age-matched control. Moreover, a significant induction in LPO was observed for old aged rats (146.7 ± 8.7 nmol/mL PRBCs) that was brought closer to the level of control upon treatment with rapamycin (116.3 ± 4.3 nmol/mL PRBCs) and metformin (99.9 ± 6.4 nmol/mL PRBCs) in old aged rats. The cotreatment group in old aged rats showed a greater decrease in LPO (86.1 ± 8.6 nmol/mL PRBCs) with respect to the age-matched control.
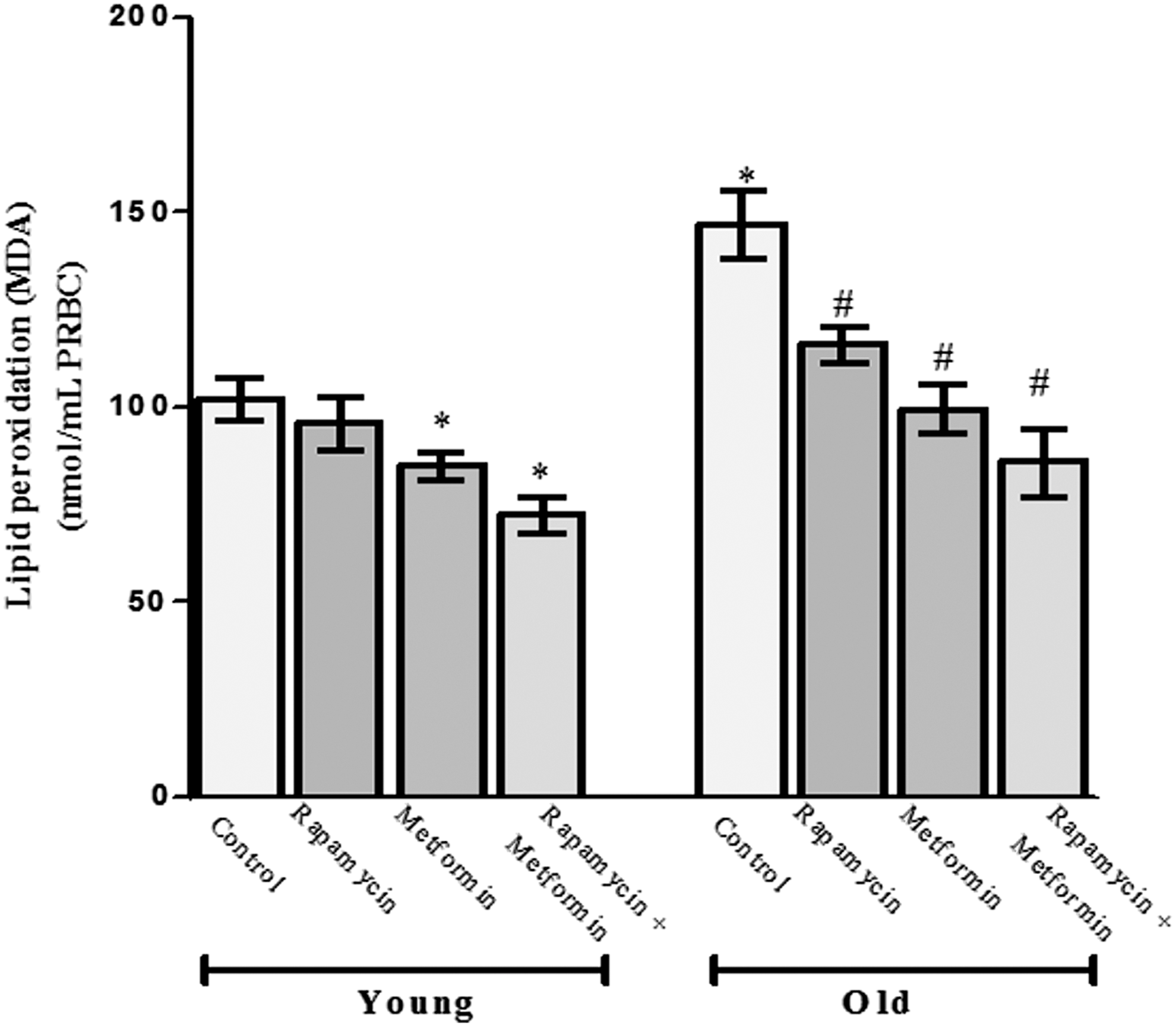
Protective effect of rapamycin, metformin, and their cotreatment on lipid peroxidation in terms of malondialdehyde (MDA) in erythrocytes of young and old aged Wistar rats. Values are mean ± SD of three independent experiments and MDA value is expressed as nmol/mL packed red blood cells (PRBCs). *p < 0.05 when compared with normal young control rats and # p < 0.05 when compared with old aged control rats.
Effect of rapamycin, metformin, and cotreatment on erythrocyte GSH
Next, we evaluated the age-dependent protective effect of rapamycin, metformin, and their cotreatment on antioxidant molecule GSH (Fig. 4). The level of GSH was significantly (p < 0.05) reduced (0.034 ± 0.003 mg/mL PRBCs) in old aged rat erythrocytes compared with the young control (0.075 ± 0.002 mg/mL PRBCs) rats. The treatment with rapamycin alone did not show any significant effect on GSH content in young rat erythrocytes; however, metformin alone and cotreatment with metformin and rapamycin significantly induced the level of GSH to 0.094 ± 0.004 and 0.096 ± 0.005 mg/mL PRBCs, respectively, in young rats. Moreover, rapamycin, metformin, and their cotreatment showed profound effect in old aged rats by increasing the level of GSH significantly to 0.047 ± 0.004, 0.055 ± 0.002, and 0.073 ± 0.006 mg/mL PRBCs, respectively. In addition, the cotreatment with rapamycin and metformin in old aged rats could be able to bring the level of GSH closer to young control.
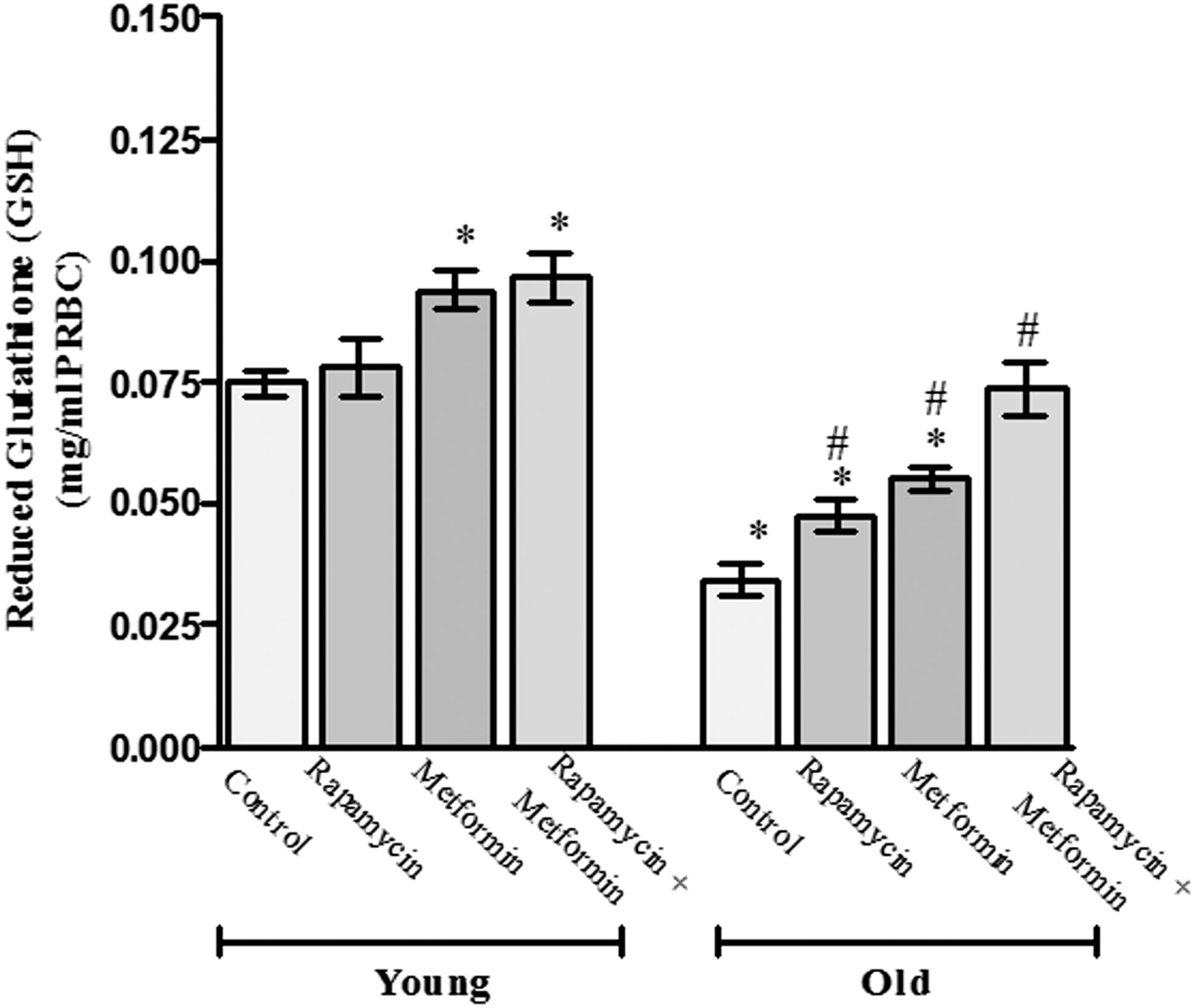
Protective effect of rapamycin, metformin, and their cotreatment on reduced glutathione (GSH) in erythrocytes of young and old aged Wistar rats. Values are mean ± SD of three independent experiments and GSH value is expressed as mg/mL PRBCs. *p < 0.05 when compared with normal young control rats and # p < 0.05 when compared with old aged control rats.
Effect of rapamycin, metformin, and cotreatment on erythrocyte PMRS activity
The result of age-dependent alteration in erythrocyte PMRS as an important regulator of antioxidant status is highlighted in Figure 5. A significant (p < 0.05) induction in PMRS activity was observed in old aged rat erythrocytes (1.64 ± 0.06 μmol ferrocyanide/mL PRBCs per 30 minutes) when compared with the young control group (0.73 ± 0.04 μmol ferrocyanide/mL PRBCs per 30 minutes). Similar to other biochemical parameters, rapamycin alone did not show any significant effect on PMRS activity in young rats. However, the administration of metformin alone and in combination with rapamycin significantly induced the PMRS activity in young as well as old aged rats. In contrast to other oxidative stress biomarkers, PMRS activity was not reduced following the supplementation of rapamycin, metformin, and their combination in erythrocytes during aging.

Protective effect of rapamycin, metformin, and their cotreatment on plasma membrane redox system (PMRS) in erythrocytes of young and old aged Wistar rats. Values are mean ± SD of three independent experiments and PMRS value is expressed as μmol ferrocyanide/mL PRBC/30 minutes. *p < 0.05 when compared with normal young control rats and # p < 0.05 when compared with old aged control rats.
Effect of rapamycin, metformin, and cotreatment on plasma PCO group
The biomarker of oxidative stress in plasma protein, that is, PCO, was measured to assess the protective effect of cotreatment with rapamycin and metformin (Fig. 6). Significant (p < 0.05) oxidation of plasma protein was observed in old aged rats (0.86 ± 0.03 nmol/mg plasma protein) when compared with untreated young rats (0.65 ± 0.05 nmol/mg plasma protein). The rapamycin, metformin, and their cotreatment showed protective effect by reducing the PCO level significantly to 0.65 ± 0.03, 0.56 ± 0.03, and 0.53 ± 0.06 nmol/mg plasma protein, respectively, in old aged rats. However, the cotreatment group showed profound protective effect in old aged rats that brought the level of PCO closer to young control group.

Protective effect of rapamycin, metformin, and their cotreatment on plasma membrane redox system plasma protein carbonyl (PCO) in young and old aged Wistar rats. Values are mean ± SD of three independent experiments and PCO value is expressed as nmol/mg protein. *p < 0.05 when compared with normal young control rats and # p < 0.05 when compared with old aged control rats.
Effect of rapamycin, metformin, and cotreatment on AChE activity
The result of erythrocyte AChE activity is highlighted in Figure 7. The AChE activity was significantly reduced in old aged rats (10.96 ± 0.32 IU/g Hb) when compared with young rats (18.4 ± 1.6 IU/g Hb). The treatment with rapamycin, metformin, and their combination did not show any significant effect on AChE activity in young erythrocytes. However, rapamycin, metformin, and their combination significantly (p < 0.05) elevated the AChE activity (18.83 ± 0.47, 20.01 ± 0.61, and 20.89 ± 0.52 IU/g Hb, respectively) when compared with old aged control group. Interestingly, the cotreatment group showed strong protection against the reduction of AChE activity in old aged rats.
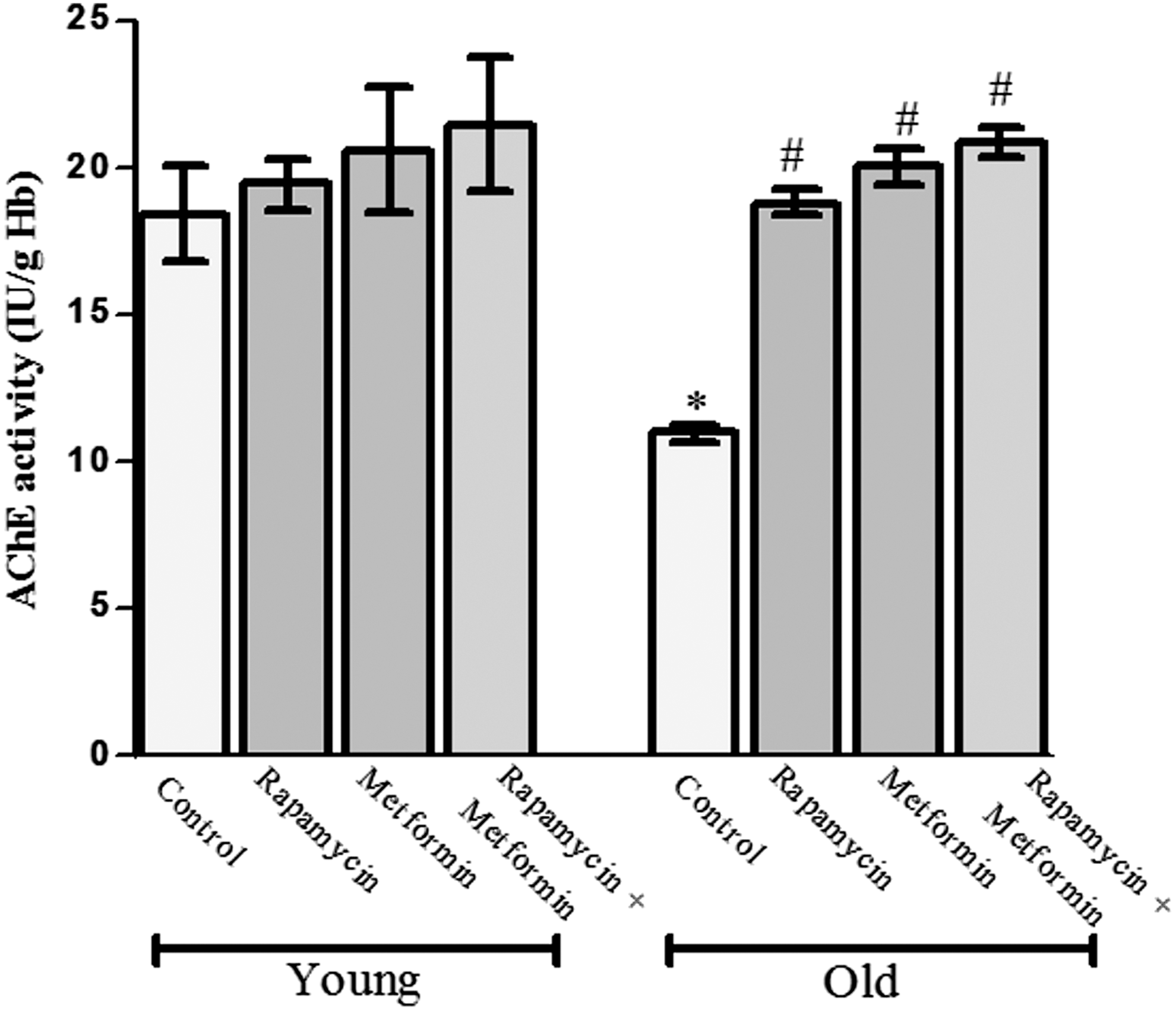
Protective effect of rapamycin, metformin, and their cotreatment on acetyl cholinesterase (AChE) activity in erythrocytes of young and old aged Wistar rats. Values are mean ± SD of three independent experiments and AChE activity is expressed as IU/g of hemoglobin. *p < 0.05 when compared with normal young control rats and # p < 0.05 when compared with old aged control rats.
Discussion
The present study deals with the investigation of age-dependent alterations in biomarkers of oxidative stress in the erythrocyte and plasma of aging rats following cotreatment with rapamycin and metformin. It has been well documented that the redox homeostasis is generally impaired by an imbalanced situation between pro-oxidant and antioxidant biomarkers during progression of age. 38,39 Disruption of the organizational structure during oxidative stress represents a fundamental mechanism in system failure and disease. 40 Erythrocytes are the best suitable model to study the mechanism of redox homeostasis as these cells are fully equipped with antioxidant defense system to scavenge pro-oxidants. 41 Moreover, the absence of protein synthesis machinery due to enucleation of erythrocytes opens different windows of vulnerability toward oxidative stress-mediated damage and therefore erythrocytes are known to act as oxidative sinks. 6
ROS, released from neutrophils, macrophages, and endothelial cells, are taken up by erythrocytes. In addition, endogenous ROS are continuously generated by the slow autoxidation of hemoglobin (Hb). The ROS generated on the RBC membrane through Hb autoxidation are ideally located to react with membrane lipids and proteins producing LPO and modified membrane proteins that can affect the membrane structure. 42 Erythrocytes possess redundant and overlapping mechanisms for protection against oxygen free radicals, including enzymes such as catalase, superoxide dismutase (SOD), GSH peroxidases and glutathione reductase, and low-molecular-weight antioxidants either produced intracellularly (GSH and NADH/NADPH) or taken up by cells (α-tocopherol, ascorbate, bioflavinoids, and selenium). 43 The age-associated deterioration of cellular structural and functional activities is mainly attributed to the enhanced production of ROS and subsequent ROS-mediated oxidative damage, as well as accumulation of macromolecules. 44
The reduction of food intake without malnutrition has been shown to impose positive effects on health and life span. CRMs that mimic the biological effects of CR reduce the age-associated accumulation of oxidatively damaged cellular macromolecules and prolong life span of organisms. 45 The most likely CRM candidates are sirtuin-1 activators such as resveratrol, 46 mTOR inhibitors such as rapamycin, 47 and insulin pathway and AMPK activators such as metformin. 48 In the present study, we have selected metformin based on our previous report that demonstrated the maintenance of erythrocyte redox balance 21 and induction of protective autophagy pathway. 49 Although the antiaging effect of rapamycin has been reported in various cell types, 50,51 we are probably for the first time reporting the protective effect of rapamycin and its cotreatment with metformin on the age-dependent impairment of redox status in rat erythrocytes and plasma. Earlier, we have shown that rapamycin protects rat erythrocytes against induced oxidative stress 52 and degenerating rat hippocampal neurons against amyloid-β-intoxication. 29
Rapamycin could be administered in association with other drugs such as resveratrol that controls some of its adverse effects. 53 Moreover, metformin in combination with other compounds such as cysteine is known to ameliorate oxidative stress and insulin resistance in streptozotocin-induced type 2 diabetes in rats. 30 Therefore, the present study provided comparative insights on the role of rapamycin and metformin alone or in combination to regulate various biomarkers of oxidative stress. Herein, we have demonstrated that the levels of ROS, LPO, and protein carbonyls (PCO) were decreased, while the levels of FRAP, GSH, and AChE were increased following the administration of rapamycin and metformin, and the most profound effect was observed with their combinatorial treatment.
The erythrocytes are highly susceptible to ROS that oxidatively damage the macromolecules and ultimately lead to aging and cell death. 42 The maintenance of redox homeostasis is essential for survival of cells, and extensive generation of ROS often induces alteration in conformation of cytoskeleton proteins and hemolysis of erythrocytes. 54 Rapamycin is a complex molecule that acts as an immunosuppressant as well as autophagy inducer through inhibition of mTOR, a central integrator of various prosurvival pathways. Autophagy plays a very important role during red cell maturation, and loss of autophagy in erythroid cells leads to defective removal of mitochondria and severe anemia in vivo. 55 Furthermore, the age-associated defective autophagy process fails to clear the accumulated macromolecules and contributes to the development of several age-dependent diseases. 56 In young age, the autophagy process is known to be active at a basal level, while in old age, this process becomes defective and contributes to increased oxidative stress, accumulation of damaged cell organelles, and oxidative stress-mediated protein aggregation. The differential effect of rapamycin on ROS generation in erythrocytes from young and old rats could be due to change in the redox status. The observed significant reduction in ROS level after rapamycin treatment of old rats may also reflect the involvement of various prosurvival signaling pathways, including autophagy, which have been well established to scavenge ROS overproduction in various cell types. 29,52 George et al. have shown that erythrocytes contain membrane-bound NADH oxidases that also contribute to the production of endogenous ROS. 57 Recently, an alteration in the expression of NADH oxidase has been reported with the advancing of age. 58 Moreover, rapamycin has been suggested to increase NADPH oxidase-mediated production of superoxide and ROS. 59 Additionally, we have previously reported a similar response of rapamycin in inducing a nonsignificant level of ROS in young rat erythrocytes. 52 Harrison et al. have shown that rapamycin extends the life span of genetically heterogeneous mice in middle and old age rather than young age, which also supports our findings of a pronounced rapamycin-mediated effect in all the studied parameters in old aged rats. 60
The combinatorial exposure of rapamycin and metformin affected all the studied parameters in erythrocytes of young as well as old aged rat erythrocytes, except ROS production in young rats. In young rats, rapamycin shows insignificant change in ROS production, while metformin significantly reduced ROS production, and their combination also reduced ROS production. Moreover, the combinatorial exposure of rapamycin and metformin reduced the level of ROS in comparison to the rapamycin-treated young group that might be due to the strong antioxidant activity of metformin.
ROS-mediated oxidative stress affects erythrocyte membrane by inducing LPO that leads to the loss of membrane integrity and cell death. 61 It is also evident from several studies that MDA level is increased in rat erythrocytes and other tissues with advancing of age. 38,62 Erythrocyte band-3-associated enzymes such as phosphofructokinase and glyceraldehyde-3-phosphate dehydrogenase are also affected by the accumulation of MDA. 63 In addition, reduction of MDA content following the administration of rapamycin has been reported in several studies. 64,65 In line with these observations, we also found reduction in MDA level after treatment with rapamycin and metformin in young and old aged rats; however, the cotreatment showed the most pronounced effect. FRAP is also an extensively studied biomarker used to evaluate the extent of oxidative stress and serves as a putative index of antioxidant capacity. 66 The ability of antioxidants found in plasma or tissues to reduce ferric to ferrous ion has been employed for estimation of the antioxidant capacity of plasma to predict oxidative stress during aging. An elevated antioxidant capacity in plasma and tissues of young and old aged rats has already been reported after metformin supplementation. 21,49
GSH is an intracellular nonenzymatic and thiol-containing antioxidant compound that detoxifies free radicals and protects membrane proteins and lipids from free radical-mediated oxidation. 67 Our earlier report has provided evidence of decreased erythrocyte and brain GSH level as a function of aging in rats. This age-related decline in GSH results from reduced expression of proteins involved in glutathione synthesis and its reduction. 68 However, there are some conflicting reports suggesting that GSH remains unaffected during adult aging in humans. 69 In our study, metformin reverses the decreased level of GSH in old aged rat erythrocytes, which is in concurrence with a previous report. 21 Furthermore, the cotreatment with rapamycin and metformin showed more pronounced improvement in the GSH level during aging. This improvement through rapamycin by reducing the oxidative stress in vivo 70 and metformin was presumably by enhancing the pentose phosphate pathway-related formation of NADP that can be used in GSH regeneration. 71
Erythrocytes contain PMRS, which transfers electrons from intracellular donors to extracellular acceptors outside the cell. PMRS enables cells to effectively counteract the oxidative damage. An elevated activity of erythrocyte PMRS has been reported during aging in humans 35,72 and rats. 62 CRMs modulate PMRS activity by increasing enzyme-dependent antioxidant protection of plasma membranes, thus playing an important role in the regulation of intracellular redox homeostasis in aged cells. 73 Erythrocyte PMRS provides protection against oxidative stress during aging and an increased PMRS activity following the treatment with rapamycin and metformin further substantiates the protective role of CRMs against free radicals in old age. Furthermore, there was no significant change observed in our study after rapamycin supplementation to young rats, whereas it has a synergistic effect with metformin in young and old groups.
Plasma PCO is a crucial marker to measure oxidative damage and protein oxidation. The carbonylation of a particular protein is linked to the loss of its function. Earlier, it has been reported that PCO level in plasma, brain, and membrane was increased significantly in old aged rats when compared with those of young rats. 49,74,75 Metformin is known to ameliorate the protein oxidation in plasma and brain of young as well as old aged rats. 21,49 In our study, rapamycin did not produce any pronounced effect in young rats, but significantly decreased the PCO level in old aged rats. However, we observed marked alteration in the level of PCO in young and old aged rats when rapamycin was supplemented in combination with metformin. In general, oxidized proteins (PCO) are usually removed by autophagy-mediated lysosomal degradation pathway, and autophagy becomes defective with advancing of age that further contributes to the elevation of PCO level and accumulation of oxidized proteins in the cells. In young rats, rapamycin decreased the PCO level nonsignificantly, which might be due to the fact that rapamycin effectively restores the age-associated defective autophagy in old aged rats. In our previous study, we have shown slight decrease in the PCO level by rapamycin in young rat erythrocytes. 52 Autophagy has also been shown to regulate erythropoiesis and remove nucleus, mitochondria, and other cell organelles during maturation of erythrocytes. 76
AChE in erythrocytes is one of the major markers of cholinergic metabolism that rapidly hydrolyzes acetylcholine to acetate and choline at the synaptic cleft and plays an essential role in neurotransmission. 77 Several reports have shown that AChE is much lower in erythrocytes and brain of old aged rats compared with young rats. 5,78 During aging, there is an increased level of oxidative damage and LPO in erythrocytes, which may lead to reduction in AChE activity. In our study, we observed nonsignificant change in AChE activity after rapamycin and metformin supplementation either alone or in combination in young rats. However, these drugs augmented the reduced AChE activity in old aged rats with greater response when given in combination. AChE is prone to oxidative stress-mediated dysfunction during aging, and the cotreatment with rapamycin and metformin, through their free radical scavenging activity, preserves the enzyme's catalytic triad and increases its activity. Taken together, the study demonstrated the synergistic effect of rapamycin and metformin cotreatment against the age-dependent oxidative stress in rat erythrocytes.
Conclusion
The treatment of old aged rats with rapamycin and metformin in combination offered the best protection against oxidative damage by effectively restoring the normal architecture of erythrocytes. As the erythrocyte is particularly susceptible toward oxidative stress-mediated damage, there is also an age-dependent variation in vulnerability against oxidative stress as well as levels of biomarkers of oxidative stress. Rapamycin has been found to be effective to maintain redox balance during aging, enhancing the ability of erythrocytes to improve antioxidant status in combination with metformin. The cotreatment has shown a synergistic effect that might provide an innovative strategy to develop and design a suitable combination therapy for the effective management of oxidative damage during aging.
Footnotes
Acknowledgments
A.K. Singh would like to acknowledge the Dr. D.S. Kothari Post Doctoral Fellowship scheme of University Grants Commission, New Delhi, India, for providing financial support (F.4-2/2006(BSR)/BL/14-15/0326) and fellowship. Department of Biochemistry is supported by an FIST grant of DST-SERB, Government of India.
Author Disclosure Statement
No competing financial interests exist.