
Abstract
The aging female rat constitutes an interesting model of spontaneous and progressive age-related dopaminergic dysfunction as it allows assessing new therapeutic strategies for Parkinson's disease. Insulin-like growth factor I (IGF-I) is emerging as a powerful neuroprotective molecule, strongly induced in the central nervous system after different insults. We constructed a helper-dependent recombinant adenoviral vector (HDRAd-IGFI) harboring the gene for rat IGF-I. This was used to implement long-term IGF-I gene therapy in the hypothalamus of aged female rats, which display hypothalamic dopaminergic (DA) dysfunction and, as a consequence, chronic hyperprolactinemia. Rejuvenating long-term IGF-I gene therapy was implemented in young (3 months) and aged (24 months) female rats, which received a single intrahypothalamic injection of 4 × 109 viral particles of either HD-RAd-IGFI or HD-RAd-DsRed (control vector) and were sacrificed 119 days postinjection. In the young animals, neither vector modified serum prolactin (PRL) levels, but in the RAd-IGFI-injected aged rats a nearly full reversion of their hyperprolactinemic status was recorded. Morphometric analysis revealed a significant increase in the total number of tyrosine hydroxylase (TH)-positive cells in the hypothalamus of experimental compared with control aged animals (5874 ± 486 and 3390 ± 498, respectively). Our results indicate that IGF-I gene therapy in aged female rats is highly effective in rejuvenating the hypothalamic DA neuron groups.
Introduction
I
Neurotrophic factors that prevent degeneration and enhance recovery of remaining DA neurons are of clinical interest. 6 Among them, insulin-like growth factor I (IGF-I) is emerging as a powerful neuroprotective molecule, synthesized in the CNS after different insults such as ischemia, 7 cortical injury, 8,9 and injury of the spinal cord. 10 In situations involving cytotoxic damage in the hippocampus, the microglia population in this region rapidly increases the production of IGF-I and IGF-I binding protein 2, which suggests a neuroprotective role of these molecules in the CNS. 11
We have previously demonstrated that long-term IGF-I gene therapy in the medial basal hypothalamus (MBH) of regularly cycling middle-aged rats extends regular cyclicity and preserves the ovarian structure of the animals. 12 In a previous study using a first-generation adenoviral vector for IGF-I, we demonstrated that in aging female rats, short-term (17 days) hypothalamic IGF-I gene therapy improves their hyperprolactinemic condition. 13 Owing to these outcomes, we constructed a helper-dependent (HD) adenoviral vector, capable of long-term expression of rat IGF-1. This was aimed at implementing long-term (3 months) IGF-I expression in the hypothalamus of hyperprolactinemic aged female rats. The present report documents the effectiveness of the new adenovector for improving the function of the hypothalamo-lactotropic axis in senescent female rats.
Materials and Methods
Helper-dependent adenovectors
Helper-dependent adenovectors (HD-RAds) were constructed using a kit sold by Microbix Biosystems (Ontario, Canada). The kit provides the shuttle plasmid pC4HSU, the helper virus H14, and the 293 Cre4 cell line. The construction procedure followed the guidelines of the Microbix manual and those described by Oka and Chan. 14 An expression cassette containing the gene for either rat IGF-I or the red fluorescent protein DsRed, both under the control of the murine cytomegalovirus (mCMV) promoter, was cloned in pC4HSU. This plasmid consists of the ITRs for Ad 5 virus, the packaging signal, and part of the E4 adenoviral region plus a stuffer noncoding DNA of human origin, which keeps a suitable size (28–31 kbp) of the viral DNA so that it is efficiently packaged into the capsids during vector generation but bands at sufficient distance from helper virus H14 in CsCl gradients, thus minimizing the risk of contamination of the newly generated vector. The shuttle vector harboring the expression cassette of interest was transfected in 293 Cre4 cells, which were then infected with the helper Ad H14 whose packaging signal is flanked by lox P sites recognized by the Cre recombinase expressed by the 293 Cre4 cells. Therefore, the helper virus provides in trans all of the viral products necessary for generation of the desired HD-RAd. Following iterated coinfections with the HD-RAd and H14 virus, a sufficiently high concentration of HD-RAd is generated, while very low levels of H14 are produced due to the cleavage of the packaging signal of H14 by the Cre recombinase. The new adenovectors, termed HD-RAd-IGF-I and HD-RAd-DsRed (Fig. 1) as appropriate, were purified by ultracentrifugation in a CsCl gradient and titrated for adenoviral particle concentration.
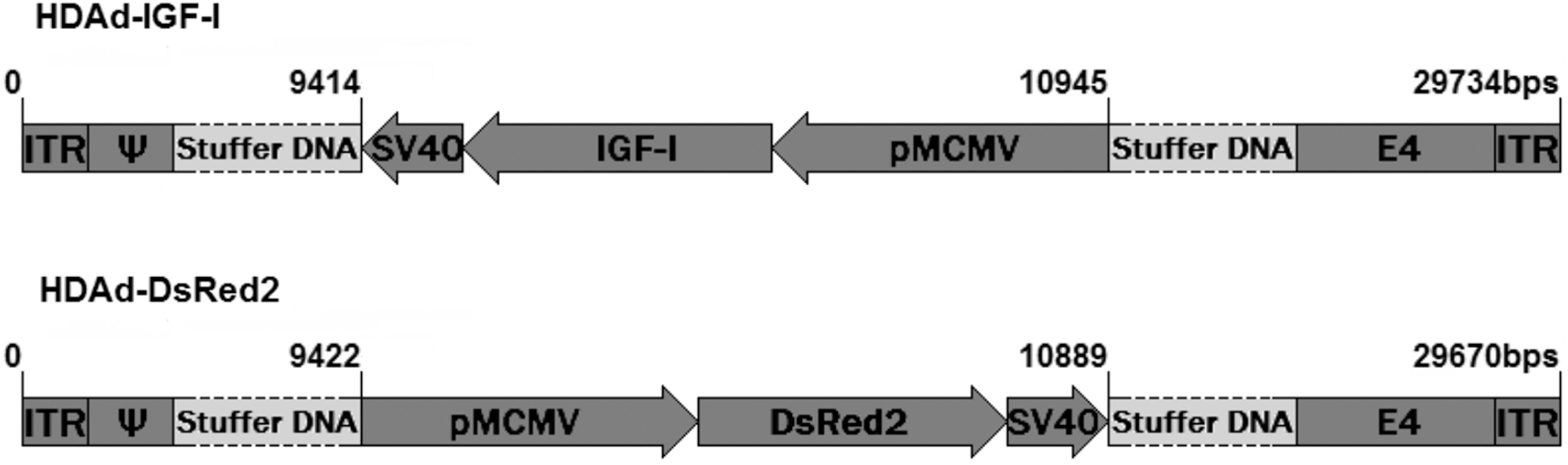
Diagrammatic representation of the genome of HD-IGF-I and HD-DsRed adenovectors. PmCMV, mouse cytomegalovirus promoter; DsRed2, reporter cDNA; IGF-I, rat IGF-I gene; SV40, simian virus 40 polyadenylation signal; ψ, packaging signal; ITR, inverted terminal repeats; E4, a portion of the E4 adenoviral gene; IGF-I, insulin-like growth factor.
RAd glial derived neurotropic factor (GDNF), a first-generation adenovector expressing the gene for rat GDNF under the control of the cytomegalovirus (CMV) promoter.
RAd-βgal, a first-generation adenovector kindly provided by Dr. Michel Perricaudet, Institut Gustave Roussy, CNRS, Paris. In this vector, the E1 genomic region has been replaced by an expression cassette containing the Escherichia coli lac Z reporter gene under the control of the CMV promoter.
The above two vectors were expanded in 293 cells, purified by CsCl gradient ultracentrifugation and titrated in 293 by plaque counting in serial virus dilutions.
Animals
Thirty-five young (3 months) and 30 aged (24 months) female Sprague-Dawley (SD) rats were used. Animals were housed in a temperature-controlled room (22°C ± 2°C) on a 12-hour light/12-hour dark cycle. Food and water were available ad libitum. All experiments with animals were performed in accordance with the Animal Welfare Guidelines of NIH (INIBIOLP's Animal Welfare Assurance No A5647-01) and approved by our Institutional IACUC (Protocol No. T09-01-2013).
Surgical procedures
Young and aged females were allotted to an intact, control or experimental group, thus forming 6 groups of 10 rats each: young intact (Y-int), young control (Y-DsRed), young experimental (Y-IGFI), aged intact (S-int), aged control (S-DsRed), and aged experimental (S-IGFI). On experimental day, −14, 14, 28, 56, 88, and 119, a small blood sample (0.3–0.4 mL) was taken from the tail veins of all rats. Serum was obtained and kept at −20°C for hormone assay.
On experimental day 0, control and experimental animals received bilateral 2-μL intrahypothalamic injections containing 4 × 109 viral particles HD-RAd-DsRed or HD-RAd-IGF-I, respectively. For this purpose, rats were anesthetized by injection of ketamine hydrochloride (40 mg/kg, i.p.) and xylazine (8 mg/kg, i.m.), and placed in a stereotaxic frame. To access the arcuate–periventricular (ARC-PeV) region, the tip of a 26-gauge needle fitted to a 10-μL syringe was brought to the following coordinates relative to the bregma: 3.0 mm posterior, 10.0 mm ventral, and 0.6 mm right and left. 15 In a separate experiment, four young rats received similar intrahypothalamic injections of either RAd-GDNF or RAd-βgal (109 plaque forming units [pfu] of each vector) and 3 days later were sacrificed for immunofluorescence (IF) observation of GDNF expression in the ARC-PeV region.
On experimental day 119, five rats from each group were sacrificed by rapid decapitation, the brain was rapidly removed from the cranium, placed on a dry ice block, and the MBH dissected and homogenized for determination of IGF-I content.
The remainder of the animals were placed under deep anesthesia and perfused with phosphate-buffered formaldehyde 4% (pH 7.4) fixative. Each brain was removed and trimmed down to a block containing the whole hypothalamus. The block was then serially cut into coronal sections 40 μm thick on a vibratome (Leica, Germany).
Immunohistochemistry
In each block, one every six serial sections was selected to obtain a set of noncontiguous serial sections spanning the whole hypothalamus. Typically, a whole hypothalamus comprised about 48 coronal sections, thus yielding 6 sets of 8 noncontiguous serial sections. For counting purposes, each set was considered as representative of the whole hypothalamus. For each animal, one set of sections was immunohistochemically processed using an anti-TH monoclonal antibody (Calbiochem, Inc., La Jolla, CA). For detection, the VECTASTAIN® Universal ABC kit (Vector Laboratories, Inc., Burlingame, CA) using 3,3-diaminobenzidine tetrahydrochloride (DAB) as chromogen, was used. Sections were dehydrated, mounted, and used for image analysis.
In brain sections of some young rats, GDNF was detected by IF using a primary mouse monoclonal antibody against human GDNF (MAb212; R&D Systems, Minneapolis, MN) and a goat anti-mouse IgG antibody conjugated to Alexa 488® (Invitrogen, Argentina). Representative hypothalamic sections were photographed with a 20 × objective, whose space calibration corresponded to 0.625 μm/pixel, with a resolution of 1360 × 1024 pixels with a pixel depth of 24 bits RGB.
Image analysis
Stereological quantitation of hypothalamic TH neurons was performed as previously described by West et al.,
16
with minor modifications. Briefly, images of the ARC-PeV areas of hypothalamic sections were captured using an Olympus DP70 digital camera attached to an Olympus BX51 microscope (Tokyo, Japan). Digital images were analyzed using the ImagePro Plus (IPP™) v5.1 image analysis software (Media Cybernetics, Silver Spring, MA). To avoid double counting errors, only TH neurons showing clearly shaped nuclei were counted. The total number (N) of TH neurons per nucleus was estimated using the following equation:
where d = length (mm) of the rostrocaudal axis of the hypothalamus being assessed; n = number of slices assessed per hypothalamus; s = thickness of sections (40 μm); and x = number of TH neurons counted per slice assessed.
Hormone determination
IGF-I was extracted from the MBH by acid-ethanol cryoprecipitation and was radioimmunoassayed (RIA) using antibody UB2-495 from L. Underwood and J.J. Van Wyk, which is distributed by Dr. A. F. Parlow, Pituitary Hormones and Antisera Center, UCLA Med. Center, USA. Recombinant human IGF-I (rh IGF-I; Chiron Corp., Emeryville, CA) was used as tracer and unlabeled ligand. Intra- and interassay coefficients of variation were 7.2% and 12.8%, respectively. Serum PRL was determined by RIA using the rat materials provided by Dr. A. F. Parlow. Serum PRL was expressed in terms of rPRL RP-3. Coefficients of variation intra- and interassay for PRL, ranged between 8.4% and 12.3%, respectively.
Statistical analysis
The one-way analysis of variance was used, as appropriate, to evaluate group differences. Tukey's method was chosen as a post hoc test. Significant differences between rat groups for each hypothalamic area were defined as those with a p < 0.05. Highly significant differences were defined as those with a p value <0.01.
Results
Both HD adenovectors expressed their transgene for at least 119 days in the hypothalamus and ependymal cell layer of both young and old rats. Shortly after injection (3 days) they showed a clear preference to transduce the ependymal cell layer and the ependymal tanycytes (Fig. 2, upper panel). On experimental day 119, DsRed expression was reduced but still clearly detectable (Fig. 2, lower panel). Tissue levels of IGF-I remained higher than endogenous IGF-I in the MBH of the experimental animals after 119 days of vector injection (Inset Fig. 2). In B92, N2a, and A549 cell cultures, HD-RAd-IGF-I, but not HD-RAd-DsRed, induced high levels of IGF-I in the supernatants (data not shown).
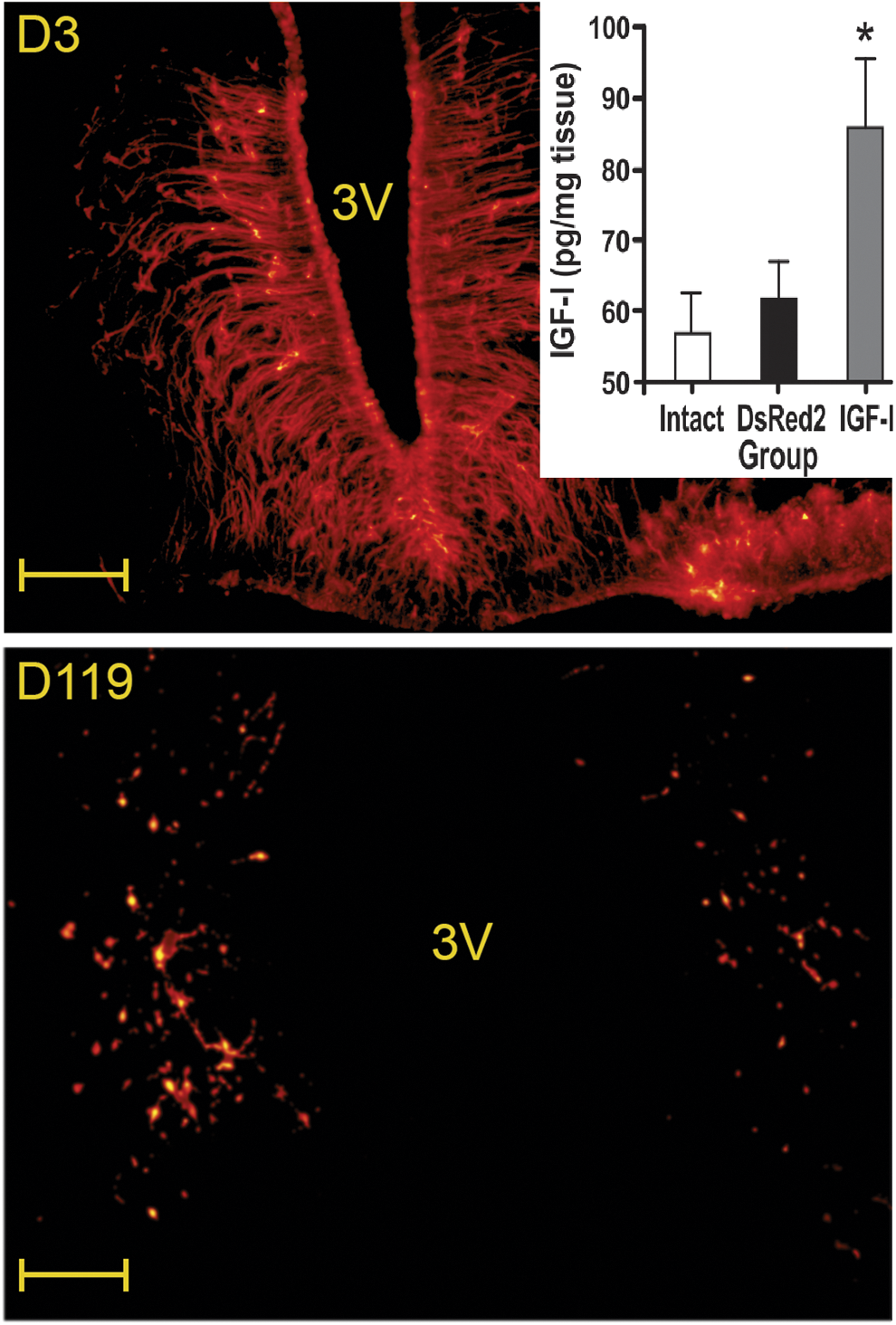
DsRed transgene expression and IGF-I levels in the MBH of DsRed and IGF-I rats. Upper and lower panels show expression levels (red fluorescence) of DsRed in the ARC-PeV region of old rats 3 and 119 days, respectively, after HD-RAd-DsRed injection. Inset shows MBH IGF-I tissue levels on Exptl. day 119 in intact, DsRed and IGF-I old animals. N was 8, 7 and 8 for the intact, DsRed and IGF-I groups, respectively. *p < 0.05 versus DsRed2. 3 V, third ventricle. Scale bars = 100 μm. ARC, arcuate-periventricular; MBH, medial basal hypothalamus. Color images are available online at
Qualitative assessment of hypothalamic sections immunostained with an anti-TH antibody revealed that the number of TH neurons was lower in the intact old rats compared with young counterparts (Fig. 3, left panels). TH immunostaining was generally fainter in the old intact rats. At the end of the experiment, old DsRed and old IGF-I rats showed a certain recovery of TH neuron immunostainability and number (Fig. 3, right panels). Quantitative immunohistochemical assessment confirmed that in the intact old rats, ARC-PeV TH neuron numbers were significantly lower than in the young counterparts (Fig. 3, inset). By contrast, TH neuron numbers in the ARC-PeV region of old DsRed and old IGF-I rats at the end of the treatment were not significantly different from their young counterparts.
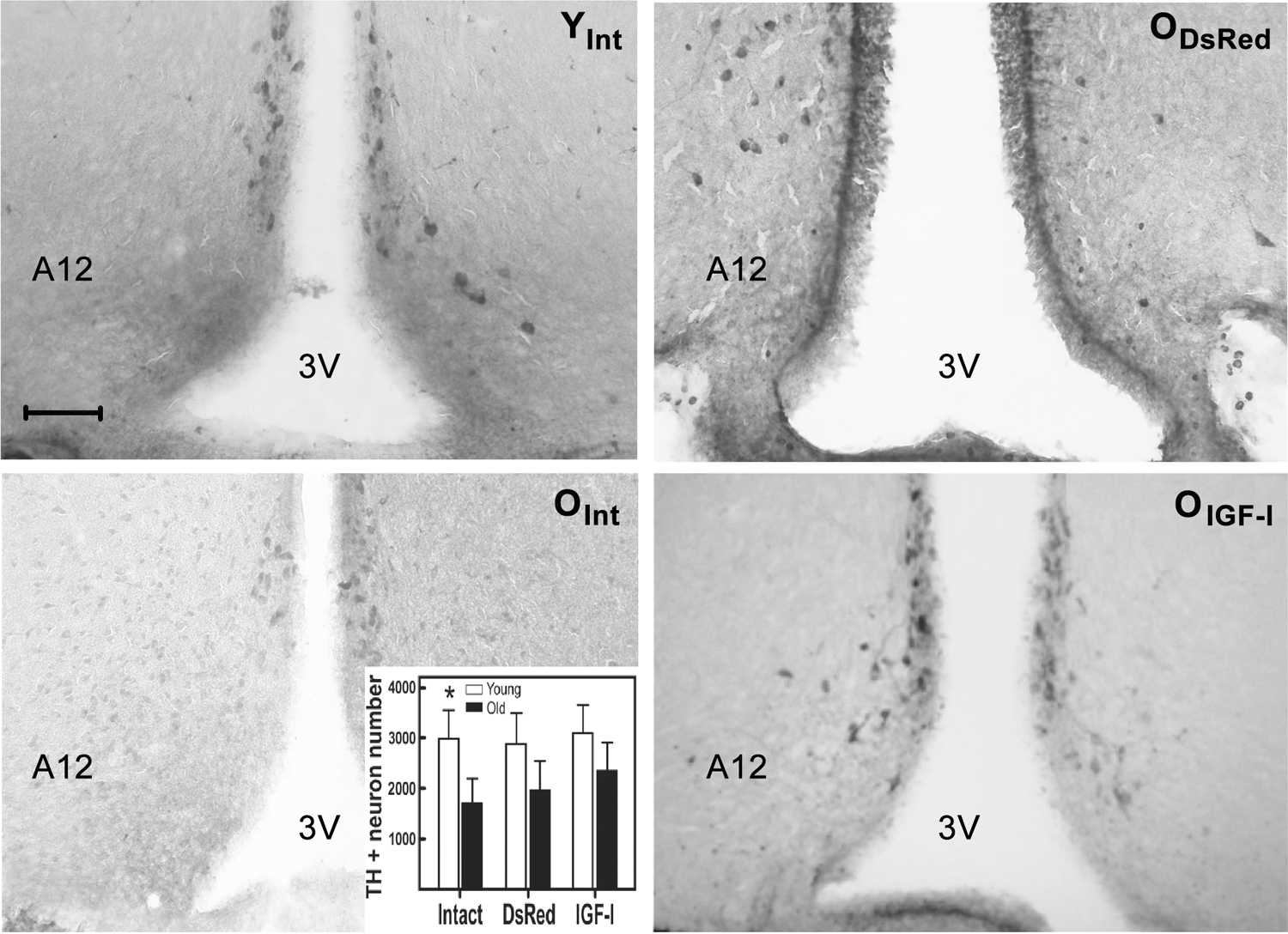
Effect of IGF-I gene therapy on the DA neurons of the ARC-PeV hypothalamic region in young and old females. The representative coronal hypothalamic sections shown pass through the medial hypothalamus and were immunolabeled with a monoclonal anti-rat TH antibody. Animals were sacrificed 119 days after the corresponding vector injection in the hypothalamus. Left panels correspond to representative young (upper) and old (lower) intact animals. Right panels correspond to representative old DsRed (upper) and IGF-I (lower) animals. Inset shows the quantitation of TH+ neurons in the ARC-PeV region in the hypothalamus. TH+ neuron counting was performed both manually and automatically using an appropriate image analysis software (see Materials and Methods section for further details). Bars over columns represent SEM values. N was 5 for all groups. Significant differences between the RAd-βgal- and RAd-IGFI-injected groups for each pair of columns are indicated by asterisks over the solid column. *p < 0.05; scale bar = 100 μm. DA, dopaminergic; SEM, standard error of the mean; TH, tyrosine hydroxylase.
In the young animals, serum PRL profiles were only affected shortly after IGF-I or DsRed vector injection, but later PRL levels returned to normal values (Fig. 4, upper panel). In contrast, treatment with both the IGF-I and DsRed HD vectors induced a marked fall in serum PRL levels in the senescent females, thus reversing the chronic hyperprolactinemia typically present in aged female rats (Fig. 4, lower panel). In the IGF-I old rat group, serum PRL remained low for at least 119 days, whereas in the DsRed old rats PRL began to rise on experimental day 88 becoming significantly higher than PRL in the IGF-I group on day 119 but still much lower than in the intact old rats, where serum PRL showed a progressive rise with age.
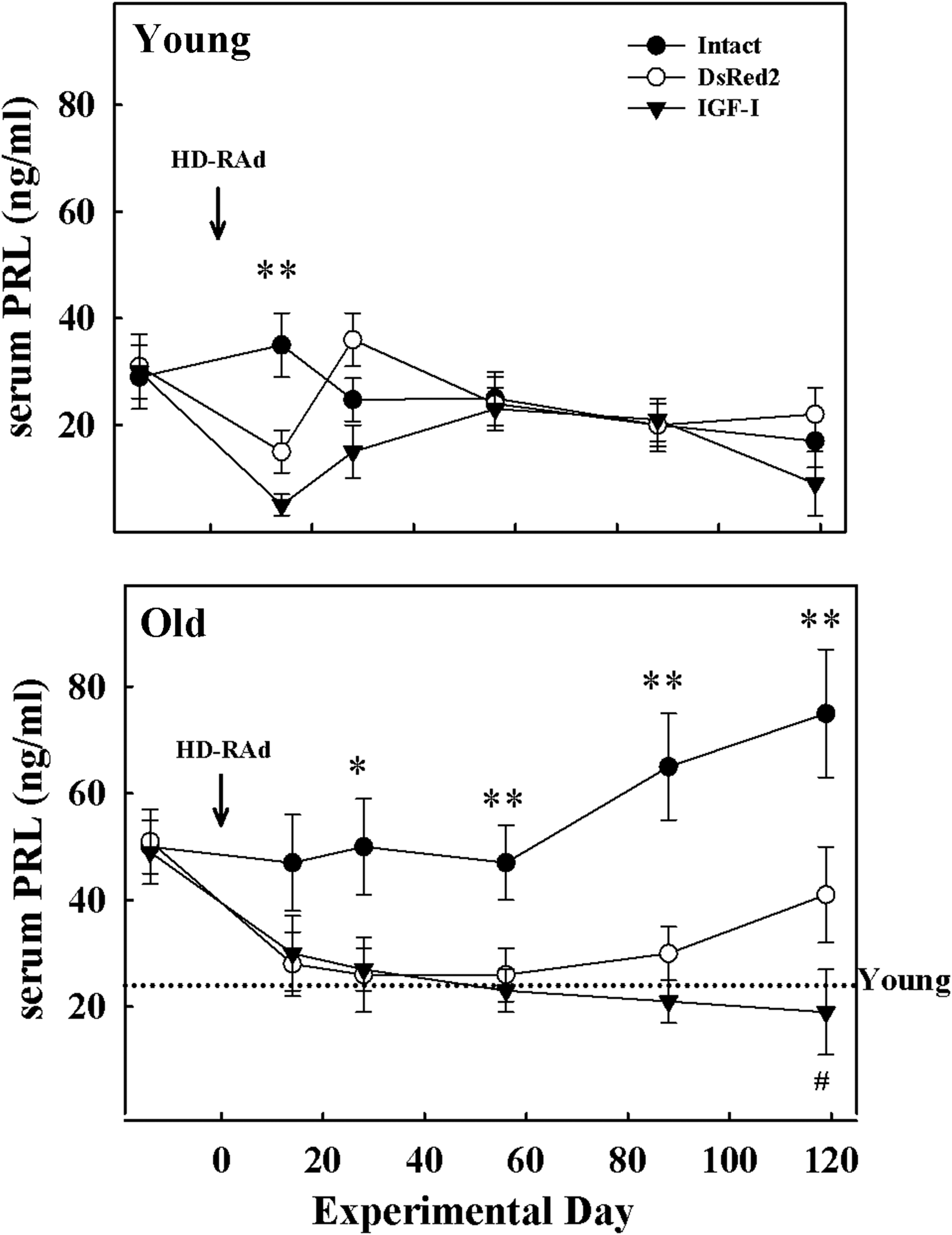
Effect of IGF-I gene therapy on serum PRL levels in young and aged female rats. The panels show the effect of intrahypothalamic injection of HD-RAd-βgal, HD-RAd-IGFI, or no injection (intact), on serum PRL profiles during a 119-day-long postinjection period. The upper and lower panels correspond to young (3 months) and aged (27 months) animals, respectively. Arrow indicates vector injection time (Exptl. day 0). Notice that at preinjection time all aged females were hyperprolactinemic compared to reference values from intact young female rats (dotted line). The number of animals in the young intact, DsRed, IGF-I, aged intact, DsRed, and IGF-I groups was 10, 9, 9, 10, 7, and 8, respectively. Bars over symbols represent SEM values. The significance of differences between intact versus the corresponding DsRed and IGF-I groups is indicated by *(p < 0.05) or **(p < 0.01) over symbols. # indicates a significant difference between IGF-I and DsRed groups on postinjection day 119. PRL, prolactin.
In a separate experiment with a separate set of young rats, intrahypothalamic injection of RAd-βgal or RAd-GDNF induced 3 days later a substantial expression of GDNF in the target area (Fig. 5, center and right panels). As expected, no GDNF expression was detectable in the hypothalamus of intact animals (Fig. 5, left panel). In B92 and N2a cell cultures, RAd-GDNF, but not RAd-βgal, induced high levels of GDNF in the supernatants (data not shown). Neither serum PRL nor IGF-I levels were determined in this complementary experiment.

Expression of GDNF in the hypothalamus of young rats. Animals were perfused and processed for GDNF IF 3 days after vector injection in the hypothalamus. Left, center, and right panels correspond to intact, RAd-βgal-, and RAd-GDNF-injected rats, respectively. N = 3 for all groups. Scale bar corresponds to 100 μm. GDNF, glial derived neurotropic factor; IF, immunofluorescence. Color images are available online at
Discussion
A number of in vivo models for Parkinson's disease have been developed for the assessment of new therapeutic strategies for this devastating disease. They include the use of neurotoxins to lesion nigral DA neurons in primates and rodents and the generation of transgenic models overexpressing α-synuclein. 17 Although each of these paradigms has provided useful information for the understanding of Parkinson's disease, they share a significant limitation namely, that the neurological lesions they study are caused by experimental manipulations rather than by aging, the only unequivocal risk factor for Parkinson's disease. 18,19 In this context, the aging female rat emerges as an interesting model of spontaneous and progressive age-related DAergic dysfunction. Besides, the functional status of TIDA neurons can be readily and humanely monitored in the animals by measuring circulating PRL levels. The advantages of the neuroendocrine system for the evaluation of gene therapy strategies in the CNS have been already demonstrated in the Brattleboro rat, a mutant lacking arginine-vasopressin (AVP), which is used as a model of diabetes insipidus. 20
When an adenoviral vector encoding the rat AVP cDNA was stereotaxically injected into the supraoptic nucleus of Brattleboro rats, a substantial expression of AVP in magnocellular cells as well as the presence of immunohistochemically detectable AVP in their axons projecting to the posterior pituitary was detected. Measurement of urine output and urine osmolality showed that the symptoms of diabetes insipidus in the Brattleboro rats were significantly reduced for up to 4 months after injection of the viral vector. 21
In a previous study, using a bicistronic adeno-associated viral vector (AAV) expressing IGF-I and DsRed, injected in the anterior hypothalamus of regularly cycling adult female rats, we showed that the treatment extended the period of regular cyclicity and preserved ovarian integrity compared with intact or AAV-DsRed-treated animals. The latter vector had a weak regular cyclicity prolonging effect. 12 As indicated above, in an earlier study we found that short-term intrahypothalamic IGF-I gene therapy in the MBH of aged female rats restored the DA neuron population of the ARC-PeV nuclei. The typical hyperprolactinemia of aged rats was also significantly reduced. 13 That study, although promising, used first-generation adenovectors, which are immunogenic and therefore not suitable for clinical use. Besides, due to the immunogenicity of these vectors, transgene expression is usually short lived as transduced cells are readily destroyed by the immune system. 22 Since we wanted to assess the long-term stability of the restorative effect of IGF-I gene therapy in the hypothalamus of aged females, we constructed HD adenovectors as gene delivery vehicles. In these adenovectors, essentially all viral genes have been deleted, thus they are devoid of any significant immunogenicity and are suitable for clinical use. Furthermore, they allow long-term expression of transgenes. 23
The present results confirm that the number of TIDA neurons decreases significantly with age in female rats 24 and suggest that intrahypothalamic long-term IGF-I gene therapy increases TH neuron number in the hypothalamus of aged females. It is likely that this increase is accounted for by a trophic effect of transgenic IGF-I on preexisting dysfunctional DA neurons expressing low levels of TH, which would be rendered TH+ by the treatment. Nevertheless, our findings are also consistent with a neurogenic effect of IGF-I in the hypothalamus of the aged animals. In fact, the neurogenic and astrogenic activity of IGF-I in the hypothalamus has been previously demonstrated in vivo in adult rats. 25 Interestingly, the HD DsRed adenovector also seems to restore, although to a lesser extent, TIDA neuron number in old rats (see further comments about this below). This finding is consistent with the PRL data in aged rats. Clearly, both the IGF-I and the DsRed adenovectors significantly reduced serum PRL levels in the old animals. In the young counterparts, this effect was detected on experimental day 14 with PRL levels returning to normal afterward. In the old rats, the PRL lowering effect of transgenic IGF-I was long lasting (at least 119 days), whereas in the DsRed counterparts, serum PRL levels progressively increased after postinjection day 56, which suggests that the effect of HD RAd-DsRed was shorter lived. We believe that if we had prolonged the experiment for 2 further months, serum PRL levels in the old DsRed rats would have risen to those of intact rats, while serum PRL might have remained low in the old IGF-I animals. Our results show that even though the treatment only partially restored TIDA neuron number in the old rats, this recovery was enough to fully reverse their chronic hyperprolactinemia.
Neither HD-RAd-DsRed nor RAd-βgal increase endogenous IGF-I production in cell cultures, which demonstrates that these reporter genes per se are unable to induce production of the peptide. The observation that injection of RAd-βgal in the MBH of young rats induced expression of GDNF (an endogenous neuroprotective molecule) suggests that this is an unspecific effect, possibly related to the tissue injury caused by the needle plus some effect of the vector per se. We have even observed that saline injection in the MBH of young rats induces some GDNF expression (Morel et al., unpublished data). Intracerebroventricular injection of HD-RAd-DsRed in old rats induced a minor increase in IGF-I levels in the cerebrospinal fluid. 26 We hypothesize that as tissue injury heals, the endogenous production of endogenous neuroprotective factors, including but not limited, to IGF-I, progressively declines. This may happen faster in young than in old rats thus accounting for the very transient effect of DsRed in young rats and a more protracted effect in old counterparts. Perhaps the availability of transgenic rats harboring a Tet-On regulatable construct specifically expressing IGF-I in the hypothalamus when doxycycline is added to the drinking water of the animals might provide an injury-free overexpression of hypothalamic IGF-I in old rats. This way the specific protective effect of IGF-I on TIDA neurons could be clarified.
Conclusions
The present study shows that long-term overexpression of IGF-I in the hypothalamus of old rats has a sustained restorative effect on TIDA neurons. It is likely that with age TIDA neurons become dysfunctional (TH negative) but remain alive and can be restored by IGF-I and perhaps other neuroprotective peptides. This hypothesis is in line with a previous study reporting that in young rats, estrogen administration fully inhibits TIDA neurons but that the effect is reversible. 27 The results also suggest that HD-adenovectors are suitable gene delivery tools for long-term treatments and may constitute a safe system for implementing long-term gene therapy in clinical trials.
Footnotes
Acknowledgments
The authors are indebted to Ms. Yolanda E. Sosa, Ms. Araceli Bigres, and Mr. Oscar Vercellini for technical assistance. E.S., G.R.M., O.A.B., and R.G.G. are career researchers of the Argentine Research Council (CONICET). G.M.C. is a career researcher of the Buenos Aires Research Commission (CIC-PBA). M.L.L. and J.I.S. are recipients of CONICET doctoral fellowships. This study was supported by grants #PICT15-0817 and #PICT13-1590 from the Argentine Agency for the promotion of science to R.G.G. and grant #FPREDM052015 from the Fondation pour la Recherche en Endocrinologie, Diabetologie et Metabolisme to E.S.
Author Disclosure Statement
No competing financial interests exist.