
Abstract
Several studies have demonstrated the differentiation of human adipose tissue-derived stem cells (hADSCs) to neuronal and glial phenotypes, but directing the fate of these cells toward dopaminergic neurons has not been frequently reported. The aim of this study was to investigate dopaminergic specification of hADSCs in vitro. ADSCs were isolated from subcutaneous abdominal adipose tissue and were characterized. For dopaminergic differentiation, a cocktail of sonic hedgehog, fibroblast growth factor 8, basic fibroblast growth factor, and brain-derived neurotrophic factor were used under a low serum condition. As the control group, the ADSCs were cultured under the same low serum condition without the dopaminergic cocktail. At the end of differentiation period, the cells expressed neuron-specific markers, NES, NSE, and NEFL, and dopaminergic markers, EN1, NURR1, PITX3, VMAT2, TH, and GIRK2 genes. TH, NURR1, and EN1 mRNAs were upregulated in the dopaminergic group compared with the control group. NEFL and TH proteins were also expressed in the differentiated cells. A total of 27.9% of the cells differentiated in dopaminergic induction medium showed positive staining for TH protein. Based on reversed-phase high-performance liquid chromatography analysis, the differentiated cells released a significant amount of dopamine in response to KCl-induced depolarization. In conclusion, results of this study indicate that hADSCs can be induced by a growth factor cocktail to produce dopamine secreting cells with possible applications for future cell replacement therapy of Parkinson's disease.
Introduction
P
Mesenchymal stem cells (MSCs) have been identified as an easily accessible source of cells for clinical application. MSCs can be transplanted with no serious ethical problem and without the risk of tumorigenesis. 9 Bone marrow-derived MSCs (BM-MSCs) and adipose tissue-derived stem cells (ADSCs) are two well-known sources of MSCs with the same transcriptional profiles for stemness genes, multipotential differentiation capacity, and immunosuppressive properties. 10 Bone marrow has been the first described and the most frequently used source of MSCs. However, some investigators have proposed ADSCs as an appropriate substitute for BM-MSCs. ADSCs can be isolated from subcutaneous adipose tissue easily and in large quantities. 11 Usually 1 g of adipose tissue contains 500 times more stem cells than 1 g of aspirated bone marrow. 12
Several studies have provided evidence regarding neural differentiation of human ADSCs (hADSCs), 13 –15 but until now only one study has focused on directing the fate of these cells toward dopaminergic phenotype. 16 In this study, we examined the capability of hADSCs for dopaminergic neuronal differentiation using a growth factor cocktail containing sonic hedgehog (SHH), fibroblast growth factor 8 (FGF8), basic FGF (bFGF), and brain-derived neurotrophic factor (BDNF), which has been previously used for dopaminergic differentiation of BM-MSCs. 17 As previously shown, SHH and FGF8 signaling pathways are necessary and sufficient for the development of dopaminergic neurons in multiple sites of neural tube. 18 bFGF promotes differentiation and survival of dopaminergic grafts, and it has a neurotrophic and neuroprotective effect on embryonic dopaminergic neurons. 19 Moreover, BDNF has significant roles in the differentiation, maturation, and survival of dopaminergic neurons. 20
In this study, we showed that hADSCs can be induced by a growth factor cocktail to produce dopamine-secreting neurons. This finding may be valuable for basic research and clinical applications, especially with regard to tissue engineering, drug discovery, and cell replacement therapy of Parkinson's disease.
Materials and Methods
Isolation and characterization of hADSCs
Adipose tissue was obtained from five women who underwent elective abdominoplasty or lipoaspiration in Shariati Hospital (Tehran, Iran). The study was approved by the Ethics Committee of National Institute of Genetic Engineering and Biotechnology (7-8-93/NIGEB), and all donors gave their informed consent. These women were all 40–50 years old without any type of dietary restriction or disease background before surgery.
The ADSCs were isolated and characterized as described previously. 21 In brief, adipose tissue was minced and digested by 2 mg/mL collagenase I (Thermo Fisher Scientific, MA) in phosphate-buffered saline (PBS) containing 2% bovine serum albumin. After filtration through 100 μm nylon filter mesh and centrifugation, stromal vascular fraction was suspended in growth medium containing Dulbecco's modified Eagle's medium (DMEM), 20% fetal bovine serum (FBS), and 1% penicillin and streptomycin (all from Gibco, Thermo Fisher Scientific) and plated at 5 × 104 cells/mL in tissue culture flasks. The cells were passaged after reaching 80%–90% confluency.
Characterization of the ADSCs by flow cytometry
Fourth-passaged ADSCs were detached using Trypsin-EDTA and fixed in 70% cold ethanol overnight at 4°C. After washing with cold PBS, the cells were incubated with primary antibodies against CD90, CD105, CD45, and CD73 proteins (all from Abcam, Cambridge, United Kingdom) for 30 minutes. Then, the cells were stained with fluorescein isothiocyanate (FITC)-conjugated goat antimouse IgG (Sigma-Aldrich, Saint Louis) or phycoerythrin-conjugated donkey antirabbit IgG (Abcam) for 30 minutes. The cells only stained with the secondary antibodies were considered as the negative control. Flow cytometry was performed by a BD FACSCalibur™ (BD Biosciences, San Jose). Analysis of the results was performed by FlowJo vX.0.6 (Tree Star, Inc., Ashland).
Adipogenic and osteogenic differentiation of the ADSCs
For adipogenic differentiation, the ADSCs were cultured in a medium containing DMEM, 10% FBS, 1 μM dexamethasone, 100 μM indomethacin, 5 μg/mL insulin, and 500 μM isobutylmethylxanthine. Two weeks after induction, the differentiated cells were fixed with 4% paraformaldehyde and lipid accumulation was detected by Oil Red O staining. For osteogenic differentiation, the cells were cultured in a medium containing DMEM, 10% FBS, 10−8 M dexamethasone, 10−3 M glycerol phosphate, 3.7 g/L sodium bicarbonate, and 0.05 g/L ascorbic acid (all from Sigma). Two weeks later, calcium deposits were assessed by Alizarin Red S staining.
Dopaminergic differentiation of the ADSCs
Third to fifth-passaged ADSCs were cultured in 1% gelatin-coated six-well tissue culture plates with a density of 20,000 cells per well. The next day, growth medium was replaced with a medium containing DMEM, 1% FBS, 0.5% B27 (50 × ; Gibco), 250 ng/mL SHH (Sigma), 100 ng/mL FGF8 (Sigma), and 50 ng/mL bFGF (Sigma). At day 9 of differentiation period, 50 ng/mL BDNF (Sigma) was added to the culture medium and the cells were incubated for 3 more days. In the control group, the ADSCs were cultured in DMEM plus 1% FBS for 12 days. Half of the culture medium was renewed every 3 days.
Reverse transcription polymerase chain reaction and quantitative real-time PCR analyses
Total RNA of the cells was extracted using High Pure RNA Isolation Kit (Roche Applied Science, Germany). In brief, 1 μg of total RNA was reverse transcribed into cDNA using a cDNA Synthesis Kit (Thermo Fisher Scientific). The cDNA samples were subjected to polymerase chain reaction (PCR) amplification with a panel of specific primers as described in Table 1. The PCR products were size fractionated by electrophoresis on a 2% agarose gel.
Quantitative assessment of gene expression by quantitative real-time PCR (qPCR) was performed using RealQ PCR Master (Ampliqon A/S, Denmark) with green dye on a Rotor-Gene™ 6000 (Corbett Research, Australia) real-time analyzer. Relative quantifications were performed using REST 2009 (Relative Expression Software Tool; Qiagen) based on a Pair Wise Fixed Reallocation Randomization Test®. 22 At least, three biologic replicates of each group were included in the qPCR experiments, and β2 microglobulin (B2M) and β-actin (ACTB) were used for normalization of the quantitative data.
Immunocytochemistry
The cells were fixed with 4% paraformaldehyde, permeabilized with 0.5% Triton X-100 (Sigma), and blocked with 10% goat serum (Gibco). Primary and secondary antibodies used in this study included monoclonal antibodies against neurofilament light polypeptide (NEFL), tyrosine hydroxylase (TH), antimouse FITC-conjugated IgG antibody (all from Sigma), and antimouse PE-conjugated IgG antibody (Santa Cruz Biotechnology, TX). DAPI staining was used to determine the total number of cells in TH staining fields. Preparations were examined using a fluorescence microscope (Nikon, Japan).
Western blot analysis
For protein analysis by Western blot, the differentiated cells were homogenized in ice-cold RIPA lysis buffer supplemented with protease inhibitor cocktail (Sigma) and centrifuged at 13,000 g for 15 minutes at 4°C. The supernatant was collected and the protein concentration was determined using the Coomassie Blue assay. For each sample, 50 μg of protein was separated using sodium dodecyl sulfate–polyacrylamide gel electrophoresis, transferred to polyvinylidene fluoride membranes, and blocked with 5% nonfat dried milk. After blocking, the membranes were incubated with primary antibodies against GAPDH (eBioscience, Thermo Fisher Scientific) and TH (Sigma) overnight at 4°C. Then, the membranes were incubated with goat antimouse horseradish peroxidase (HRP)-conjugated secondary IgG for 1 hour at room temperature. The immunoreactive bands were detected using an enhanced chemiluminescence (ECL) blotting Kit (CMG Co., Iran).
Flow cytometry analysis of TH protein expression
Three-week differentiated cells were detached using Trypsin-EDTA and fixed in 4% paraformaldehyde. The cells were permeabilized with 0.2% Triton X-100. After washing, the cells were incubated with primary antibody for TH and then with FITC-conjugated goat antimouse IgG (both from Sigma). The cells only stained with the secondary antibody were used as the negative control. Flow cytometry was performed by a BD FACSCalibur (BD Biosciences). Analysis of the results was performed by FlowJo vX.0.6 (Tree Star, Inc.).
Reversed-phase high-performance liquid chromatography
After differentiation of the ADSCs for 12 days, the cells were rinsed twice with Hank's balanced salt solution (HBSS) and then were incubated in HBSS containing 56 mM KCl. The buffer was collected, stabilized using orthophosphoric acid, perchloric acid, and EDTA, and stored at −70°C. Dopamine release was measured using high-performance liquid chromatography (HPLC) with electrochemical detection (ECD). Before analysis, methanol and perchloric acid were added to each sample after vortexing and centrifugation, and 10 μL of the supernatant was injected into the chromatograph. Detection was performed by an LKB Bromma 2150 solvent delivery pump (LKB, Sweden), an EICOM ECD-100 electrochemical detector at +600 mV and highest sensitivity, and data were analyzed using Clarity Lite HPLC software (Data Apex, Petrzilkova, Czech Republic). Analyses were run at a flow rate of 1.5 mL/min at ambient temperature. A mixture of dopamine and its metabolites was used as external standards.
Results
Isolation and characterization of hADSCs
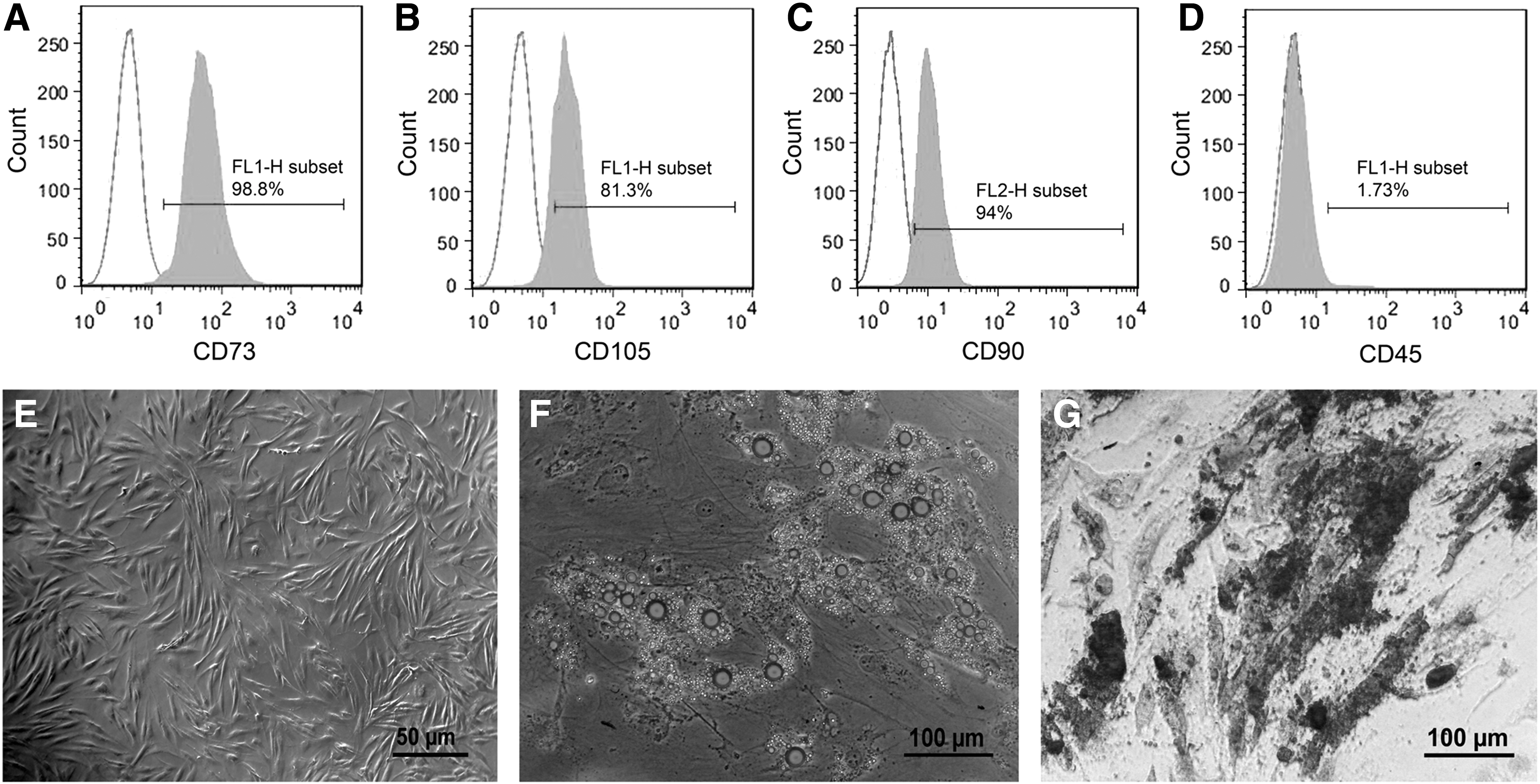
Within the first day after plating, ADSCs adhered to the plastic surfaces of tissue culture plates. ADSCs proliferated rapidly and were passaged two to three times a week. Based on flow cytometry analysis, 98.8%, 81.3%, and 94% of the fourth-passaged ADSCs were positive for the expression of MSC markers, CD73, CD105, and CD90, respectively (Fig. 1A–C). The hematopoietic marker CD45 was only expressed in 1.73% of the cells (Fig. 1D). As shown in Figure 1E, third-passaged ADSCs showed a fibroblast-like morphology. To show multipotential differentiation capability, the fourth-passaged ADSCs were differentiated into adipogenic and osteogenic lineages. After adipogenic differentiation for 3 weeks, lipid accumulation in the cells was confirmed by Oil Red O staining (Fig. 1F). Osteogenic differentiation of the 3-week differentiated ADSCs was confirmed by Alizarin Red S staining of calcium deposits (Fig. 1G). The ADSCs cultured for the same time period in 10% FBS-containing medium showed negative staining for Oil Red O and Alizarin Red S.
Neural differentiation of the ADSCs
After induction of dopaminergic differentiation, the ADSCs gradually showed a more developed neuron-like morphology with long neuronal processes. Figure 2A–E shows the ADSCs at days 1, 3, 6, 9, and 12 of differentiation, respectively. Neuronal processes are more distinguishable at higher magnification among 12-day differentiated cells (Fig. 2G). The cells differentiated in 10% and 1% FBS-containing medium without any additional factor (Fig. 2F, H, respectively) did not show neuron-like phenotype.

In the 1% FBS-containing medium with dopaminergic-inducing growth factor cocktail, the ADSCs gradually showed a more developed neuron-like morphology with long neuronal processes.
Gene expression analysis
The ADSCs that were differentiated at the presence of dopaminergic growth factors cocktail expressed NES (nestin), NSE (neuron-specific enolase), NEFL, EN1 (engrailed 1), NURR1 (nuclear receptor related 1 protein), PITX3 (pituitary homeobox 3), VMAT2 (vesicular monoamine transporter), and TH mRNAs. The undifferentiated ADSCs and the cells cultured in the control 1% FBS condition (without growth factors) expressed NES, NSE, EN1, NURR1, and TH mRNAs. Human neuroblastoma cell line SK-N-BE(2)C cells were used as the positive control for TH-expressing cells (Fig. 3).
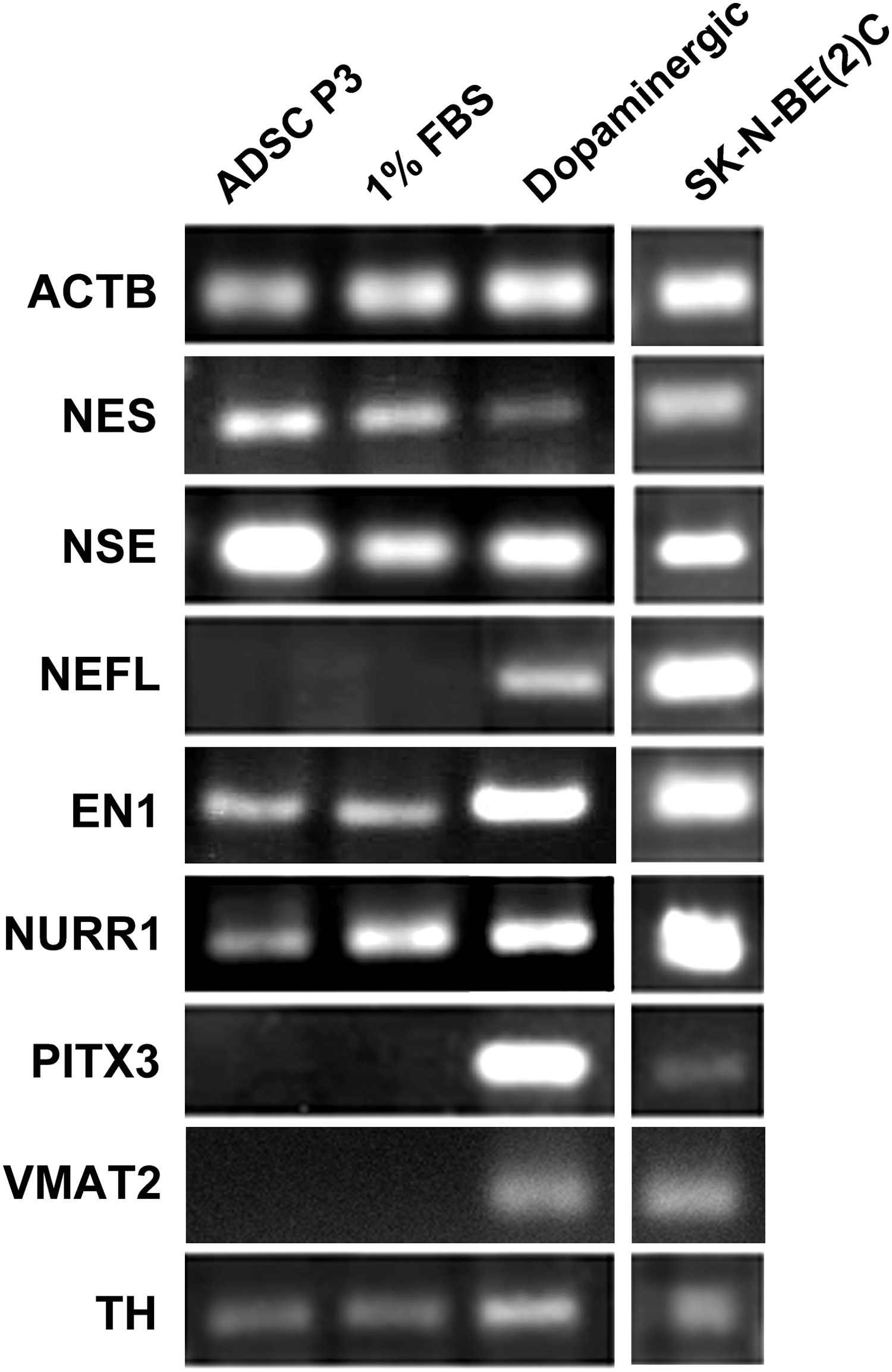
Reverse transcription polymerase chain reaction analysis for the expression of some early and late neuronal markers as well as some genes specific to dopaminergic neurons. ADSC P3: third-passaged ADSCs just before differentiation, 1% FBS: the ADSCs differentiated in the 1% FBS-containing medium without dopaminergic-inducing growth factors, dopaminergic: the ADSCs differentiated in the 1% FBS-containing medium with dopaminergic-inducing growth factor cocktail, SK-N-BE(2)C: the human neuroblastoma cell line used as the positive control.
qPCR analysis showed that the expression of EN1 in the dopaminergic group was 2.02-fold (p < 0.001) higher than the control (1% FBS without dopaminergic cocktail) group (Fig. 4A). The expression of NURR1 was upregulated by 2.32-fold (p = 0.014) with the addition of dopaminergic cocktail (Fig. 4B). TH expression in the dopaminergic group was 3.935 (p = 0.009) folds higher than the control (1% FBS) group (Fig. 4C).

qPCR and RT-PCR analyses for the expression of some dopaminergic genes.
We also assessed the expression of GIRK2 (G-protein-regulated inward-rectifier potassium channel 2) and CALB1 genes that are expressed in ventral mesencephalic dopaminergic neurons. 23 Both genes were weakly expressed in the undifferentiated ADSCs. GIRK2 was also expressed weakly in the ADSCs after differentiation in 1% FBS condition. However, the ADSCs differentiated in dopaminergic induction medium showed an upregulated GIRK2 expression, but did not express CALB1 gene (Fig. 4D).
Immunocytochemistry, Western blot, and flow cytometry analyses
As revealed by immunocytochemistry and Western blot analyses, the ADSCs that were differentiated in the dopaminergic induction medium expressed NEFL (Fig. 5A, B) and TH (Fig. 5E, F, and I) proteins, whereas these proteins were not expressed in the control 1% FBS condition (Fig. 5C, D for NEFL and Fig. 5G–I for TH proteins).

Immunocytochemistry, Western blot, and flow cytometry analyses for the expression of some dopaminergic markers.
Based on flow cytometry analysis, about 0.6% of the ADSCs that were differentiated in 1% FBS condition (Fig. 5J) and 27.9% of the ADSCs differentiated in the dopaminergic induction medium showed positive staining for TH protein (Fig. 5K).
HPLC analysis
To confirm generation of functional dopaminergic neurons, dopamine release from the differentiated ADSCs was examined by reverse phase HPLC. The ADSCs that were induced by a dopaminergic growth factor cocktail released a significant amount of dopamine (505 ± 95 pg/mL) in response to the depolarizing stimulus of 56 mM K+ (Fig. 6C, D). Dopamine release was not detected for the control group (Fig. 6B).
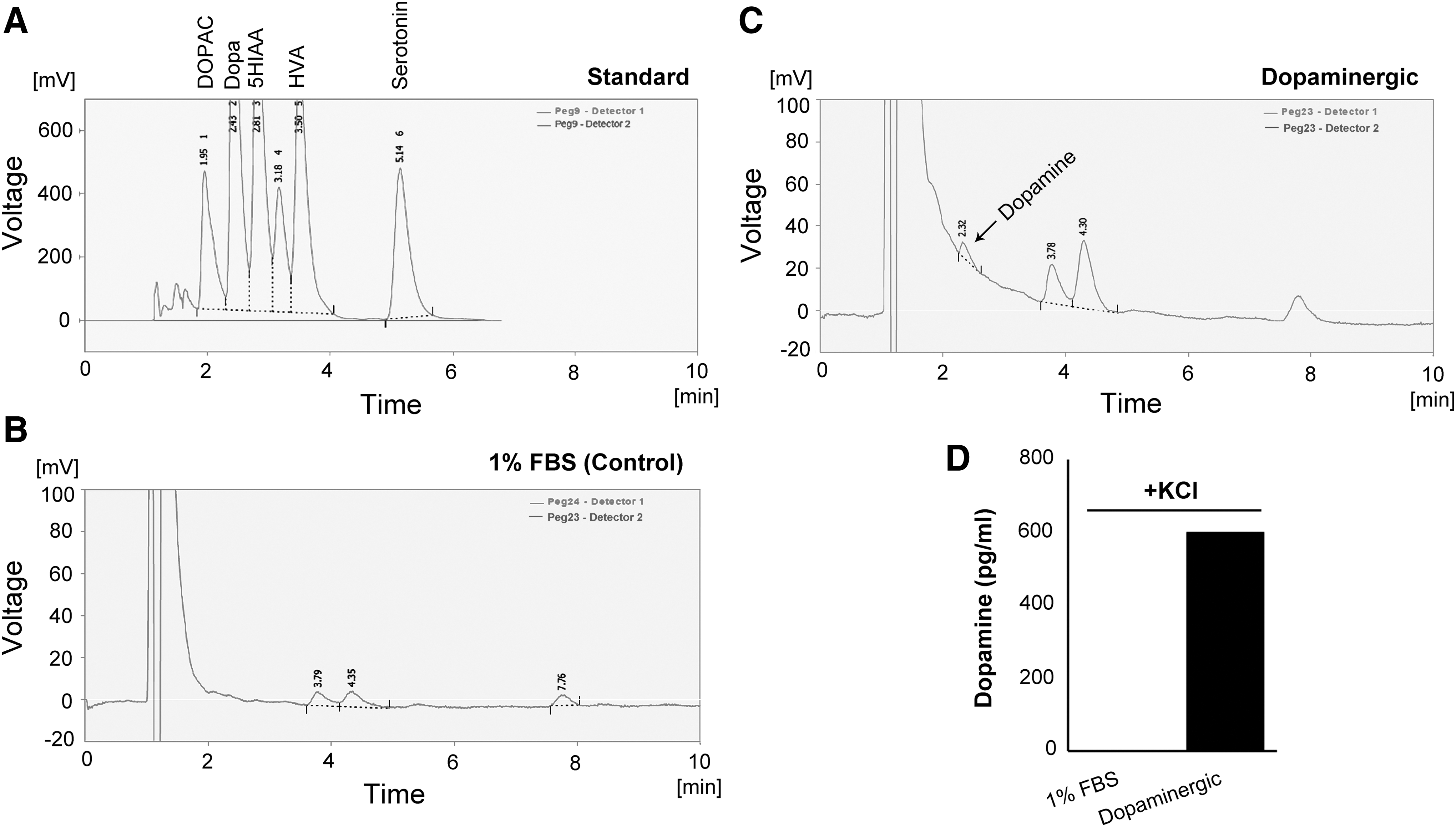
HPLC quantification of dopamine release by the ADSC-derived dopaminergic neurons after KCl-induced depolarization for 15 minutes. A mixture of dopamine and its metabolites was used as the external standards.
Discussion
In this study, we used a low-serum condition (1% FBS) and a cocktail of growth factors, containing SHH, bFGF, FGF8, and BDNF to induce dopaminergic differentiation of hADSCs. This dopaminergic-inducing cocktail has been previously used for differentiation of human BM-MSCs toward dopaminergic neurons. 17
Undifferentiated ADSCs expressed NES, NSE, NURR1, and TH genes, which indicated the capability of ADSCs for dopaminergic differentiation. After induction with dopaminergic cocktail of growth factors, the ADSCs expressed NES, NSE, and NEFL mRNAs. NES is a marker of NSCs and neural precursors, 24 and NSE and NEFL are early and mature neuronal markers, respectively. 25,26 The differentiated cells also expressed EN1, NURR1, PITX3, and TH that are important for development of postmitotic dopaminergic neurons 27 and VMAT2 that packages intracellular dopamine into small synaptic and dense core vesicles for subsequent release from dopaminergic neurons. 28,29 In our study, PITX3 and VMAT2 were only expressed in the dopaminergic induction medium. Moreover, the expression of EN1, NURR1, and TH was significantly upregulated in the dopaminergic induction medium compared with the control 1% FBS condition. These findings are in agreement with some previous reports indicating that FGF8 and SHH can induce the expression of EN1 and NURR1 genes, respectively. 30 NURR1 is a transcription activator of TH and appears to regulate dopamine neurotransmitter synthesis. 31 EN1 is involved in survival and maintenance of mesodiencephalic dopaminergic neurons, 32 and PITX3 is essential for the transition from postmitotic dopaminergic progenitors to TH-expressing neurons. 33 TH is the first and rate-limiting enzyme of dopamine biosynthesis. 34 The expression of TH begins in the differentiating dopaminergic neurons right after cessation of proliferation. 33 During development, a delicate crosstalk between EN1 and PITX3 is essential for development of distinct subsets of mesodiencephalic dopaminergic neurons. 35
We also assessed the expression of GIRK2 and CALB1 genes in the naive and differentiated ADSCs. GIRK2 and calbindin (protein coded by CALB1 gene) are two known markers of ventral mesencephalic dopaminergic neuron, 23 which are differentially expressed in the A9 neurons of the substantia nigra pars compacta and the A10 neurons of the ventral tegmental area, respectively. 23 Surprisingly, in contrast to findings in naive BM-MSCs of cynomolgus monkeys, 36 both GIRK2 and CALB1 genes were weakly expressed in the undifferentiated hADSCs. The ADSCs differentiated in dopaminergic induction medium showed an upregulated GIRK2 expression but did not express CALB1 mRNA. This finding may show the stimulatory role of SHH, bFGF, FGF8, and BDNF cocktail in the development of the A9 dopaminergic neurons from hADSCs.
As detected by immunocytochemistry and Western blot analyses, the ADSCs differentiated in the dopaminergic induction medium expressed NEFL and TH proteins, and flow cytometric analysis showed expression of TH protein in a significant proportion of the cells. These cells also released a significant amount of dopamine in response to KCl-induced depolarization, which was greater than the amount previously reported for BM-MSC-derived dopaminergic neurons. 37 –39 Recently, Boroujeni et al. 16 induced dopaminergic differentiation of hADSCs with a combination of choroid plexus epithelial cells-conditioned medium and knockout serum replacement, and demonstrated a similar level of dopamine secretion (560 pg/mL). Altogether, these findings show generation of functional dopamine-secreting neurons from hADSCs.
In conclusion, this study successfully showed the differentiation of hADSCs toward dopamine-secreting neurons using a cocktail of growth factors. ADSC-derived dopaminergic neurons may be a valuable source for future basic and clinical applications, especially with regard to tissue engineering, drug discovery, and cell replacement therapy of Parkinson's disease.
Footnotes
Acknowledgment
This work was supported by Iran National Science Foundation (grant no. 78040521).
Author Disclosure Statement
No competing financial interests exist.