
Abstract
Wu-tou decoction (WTD) is a classic traditional Chinese medicine formula and has been extensively used for the treatment of rheumatoid arthritis (RA). Previous reports indicate that WTD possesses anti-inflammatory and antinociceptive activities, and inhibits the development of arthritic joints and disease severity of collagen-induced arthritis (CIA) or adjuvant-induced rats; however, its action on angiogenesis of RA has not been clarified. This study aims to determine the anti-angiogenic activity of WTD in CIA rats and in various angiogenesis models. Our data showed that WTD (0.95, 1.9, and 3.8 g/kg) markedly reduced the immature blood vessels in synovial membrane tissues of inflamed joints from CIA rats. It also inhibited in vivo angiogenesis in chick embryo and VEGF165-induced microvessel sprout formation ex vivo. Meanwhile, WTD suppressed VEGF165-/MH7A-induced migration, invasion, adhesion, and tube formation of human umbilical vein endothelial cells (HUVECs). Moreover, WTD significantly reduced the expression of angiogenic activators, including vascular endothelial growth factor (VEGF), VEGFR2, interleukin (IL)-1β, IL-17, transforming growth factor-β, platelet-derived growth factor, placenta growth factor, angiopoietin (Ang) I and Ang II in synovium of CIA rats, and/or in HUVECs. More interestingly, WTD blocked the autophosphorylation of VEGF165-induced VEGFR2 and consequently downregulated the signaling pathways of activated AKT, ERK1/2, JNK, and p38 in VEGF165-induced HUVECs. These findings suggest for the first time that WTD possesses the anti-angiogenic effect in RA in vivo, ex vivo, and in vitro by interrupting the targeting of VEGFR2 activation.
Introduction
R
Angiogenesis is a complex process that is regulated by angiogenic mediators, including growth factors, primarily vascular endothelial growth factor (VEGF) and hypoxia-inducible factors (HIFs), as well as proinflammatory cytokines, proteases, and others. 3 Among them, VEGF is the key regulator of angiogenesis and has been implicated in various biological activities, such as stimulating endothelial cell proliferation, migration, and tube formation. 5 VEGF binds to vascular endothelial growth factor receptor 2 (VEGFR2), one of the primary receptors in angiogenesis. 6,7 VEGFR2 is upregulated at sites where VEGF is produced or increased in the synovium in response to proinflammatory cytokines. 8 –12 The binding of VEGF to VEGFR2 leads to the activation of diverse intracellular signaling molecules, including phosphoinositide 3-kinase/AKT kinase, extracellular signal-regulated kinases (ERKs), and p38 mitogen-activated protein kinase (p38-MAPK). 13,14 Therefore, targeting VEGF-induced VEGFR2-mediated signaling in ECs is important for RA treatment. 4 –7
Wu-tou decoction (WTD), a classic traditional Chinese medicine (TCM) formula from Chinese medical sage Zhongjing Zhang, has been recorded in “Jin Gui Yao Lue” for more than 1,000 years. WTD is formulated according to cold-heat theory of TCM, especially for the treatment of cold pattern of Bi Zheng, functioning as relieving pains, scattering cold pathogen, draining dampness, dispelling wind, and freeing the flow of network vessels. Accordingly, WTD is primarily used in RA patients commonly presenting with obvious pain, swelling, and dyskinesia of joint plus systematic cold signs and symptoms. WTD has been used as a complementary and alternative medicine for the treatment of RA clinically. Previous studies confirmed that WTD had the antinociceptive activity in complete Freund's adjuvant (CFA)-induced chronic inflammatory pain mice, 15 and the anti-inflammatory actions by inhibiting proinflammatory cytokines such as interleukin (IL)-1β, IL-2, IL-6, tumor necrosis factor-α (TNF-α), macrophage inflammatory protein (MIP)-1α, MIP-2, RANTES, and IP-10, and suppressing CC chemokine receptor 5 signal pathway both in CFA-induced arthritis and collagen-induced arthritis (CIA) rats. 16 –19 Besides, WTD treatments markedly reduced the expression levels of HIF-a and VEGF proteins both in the sera of CIA rats and in human fibroblast-like synoviocytes (HFLS)-RA, 17 indicating that this formula may have an anti-angiogenic effect for RA. Therefore, we investigated the anti-angiogenic activity of WTD in CIA rats and in various angiogenesis models, and its possible mechanism of action associated with VEGF-induced VEGFR2-mediated signaling pathway was also explored.
Materials and Methods
The study was approved by Research Ethics Committee of China Academy of Chinese Medical Sciences, in accordance with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. All animals were treated in accordance with the guidelines and regulations for the use and care of animals of the Center for Laboratory Animal Care, China Academy of Chinese Medical Sciences.
Water extract of WTD
WTD is a mixture of five traditional drugs, including Glycyrrhiza Radix Preparata (9 g), Ephedrae Herba (9 g), Paeoniae Radix Alba (9 g), Aconiti Radix Cocta (6 g), and Astragali Radix (9 g). The drugs were purchased from Tong-Ren-Tang Ltd. (Beijing, China), and authenticated by Professor Hu Shilin, Institute of Chinese Materia Medica, China Academy of Chinese Medical Sciences. WTD was extracted by water and then analyzed according to our previous study. 16,18,19
Induction of CIA and WTD treatment
Eighty male SD rats (8–12 week old) were obtained from Experimental Animal Center, Academy of Military Medical Sciences (production license No: SCXK 2002-001). SD rats were divided separately into five groups randomly with equal number (n = 16): the normal control group (Control), the CIA model group (Model), and CIA rats were treated with WTD at 0.95, 1.9, and 3.8 g/kg dosages, respectively. Each rat was injected emulsion, as following, to induce arthritis, except the control group:
CIA was induced as previously reported. 20 Briefly, male SD rats were injected intradermally at the base of the tail with 100 μg bovine type II collagen (Chondrex, Redmond, WA) in 0.05 M acetic acid emulsified in incomplete Freund's adjuvant (IFA; Chondrex). On day 7, rats were boosted intraperitoneally with 100 μg type II collagen in IFA.
The route of WTD delivery was oral administration intragastrically using syringe feeding. Treatment was given daily for a period of 28 days from day 1 after primary immunization. The dosage selection for WTD (0.95–3.8 g/kg/day) was based on the results of our previous study, 17 and the middle dosage of WTD for CIA rats is nearly equivalent to RA patient dosage daily. The model group and the normal rats were given an equal volume of distilled water.
Histology and histologic scoring
Rats were sacrificed by cervical dislocation on day 28 after first immunization. Both hind knees were dissected and prepared to sections for staining with hematoxylin and eosin (H&E). All sections were randomized and evaluated by two trained observers who were blinded to the treatment groups and the arthritis severity of each rat. Minor differences between observers were resolved by mutual agreement. The data were expressed as mean synovial vascularity (angiogenesis). The score was based on a scale of 0–3, as previously described. 21
Histochemical and immunohistochemical analysis
To measure blood vessel in synovial membrane tissues of inflamed joints, the polyclonal antibody (rat antibody, dilution 1:50; Abcam, Cambridge, MA) recognizing the CD31 panendothelial antigen was used for microvessel and single endothelial cell staining on 4-μm-thick paraffin-embedded sections of knee joints by immunohistochemical analysis. 22 For CD31 (Abcam) and alpha smooth muscle actin (αSMA, rabbit antibody, dilution 1:300; Abcam) immunofluorescence studies, the sections were incubated overnight in 4°C. Then, the sections were incubated for 1 h at room temperature with goat anti-rat secondary antibody (dilution 1:400; EarthOx) and goat anti-rabbit secondary antibody (dilution 1:400; EarthOx). 23,24 The results are expressed as the mean region of interest, representing the percentage of area covered with positively stained cells per image at a magnification of 400 × . The vessels were determined using Pro-Plus Image 7 after dual staining for CD31 and αSMA.
Paraffin sections of joints were mounted on poly-L-lysine-coated slides. The paraffin sections were dewaxed by routine method and incubated for 10 min with 3% H2O2. The sections were placed in a 37°C, 0.1% trypsinase for 5–30 min for antigen retrieval. Each section was incubated with normal goat serum for 20 min at room temperature, and then with primary antibody against rat VEGF (rabbit antibody, dilution 1:50; Abcam) and VEGFR2 (rabbit antibody, dilution 1:50; Cell Signaling, Boston) overnight at 4°C. After incubation with Polymer Helper for 20 min at 37°C, sections were reacted with poly-HRP anti-rabbit IgG for 20 min at 37°C. The sections were then stained with 3, 3-diaminobenzidine (Sigma, St. Louis, MO) and counterstained with hematoxylin. Specimens were examined using a Leica image analyzer and analyzed by computer image analysis (Leica Microsystem Wetzlar Gmbh., Wetzlar, Germany) in a blinded manner. To localize and identify areas with positively stained cells, 10 random digital images per specimen of the synovial membrane tissues were recorded, and quantitative analysis was performed according to the color cell separation. The results are expressed as the mean region of interest, representing the percentage of area covered with positively stained cells per image at a magnification of 400 × .
In vivo chick chorioallantoic membrane assay
A chorioallantoic membrane (CAM) assay as previously described 25 was performed to determine the in vivo anti-angiogenic activity of WTD. Briefly, fertilized chick embryos were preincubated for 6 days at 37°C in 60%–80% humidity. Fifty microliters of different concentrations of WTD (0.2, 1 and 5 μg/mL) was loaded onto a 12 × 10 × 1 mm diameter Silicone ring. The ring was then applied to the CAM of a 6-day-old embryo. After incubation for 48 h at 37°C, angiogenesis in the ring was photographed with a digital camera. The area of blood vessels was counted in 0 and 48 h. We calculated the new vascular number from 48 h minus 0 h. Five eggs per group were used in each group.
Ex vivo rat aortic ring assay
This assay was carried out as previously described 26 with minor modifications. Forty eight-well plates were covered by 100 μL of Matrigel (5 mg/mL; Corning) and left to polymerize for 45 min at 37°C. Aortic rings were prepared from SD rats. Aortas were sectioned into sections 1–1.5 mm long, rinsed several times with PBS, placed on Matrigel in wells, and covered with an additional 50 μL of Matrigel for 45 min. The rings were cultured in 1 mL of H-DMEM medium with 10% fetal bovine serum (FBS) with or without VEGF165 (20 ng/mL; Peprotech) plus various concentrations of WTD (0.2, 1, and 5 μg/mL). The medium was changed every 3 days. After 9 days of incubation, the rings were fixed with 4% paraformaldehyde. The microvessel growth was photographed using phase-contrast microscopy. Numbers and lengths of the vascular branches in aortic ring were measured in Pro-Plus Image 7. All experiments were done in triplicate.
Cell culture
Human umbilical vein endothelial cells (HUVECs) were purchased from ScienCell, Inc. (Carlsbad, CA). The cells were cultured in sterile DMEM/High Glucose (H-DMEM; Hyclone) supplemented with 5% FBS, 100 U/mL penicillin, and 80 U/mL streptomycin, and were maintained at 37°C in a humidified 5% CO2 incubator. HUVECs were used at passage numbers 4–6 in this study.
MH7A, a cell line of HFLS-RA, was obtained from the Riken cell bank (Ibaraki, Japan). The cells were cultured in sterile RPMI Medium Modified supplemented with 10% FBS, 100 U/mL penicillin, and 80 U/mL streptomycin, and were maintained at 37°C in a humidified 5% CO2 incubator.
Cell viability assay
HUVECs (5 × 104 cells/mL) were seeded in 96-well plates and incubated in sterile H-DMEM supplemented with 5% FBS, 100 U/mL penicillin, and 80 U/mL streptomycin for 24 h. Cells were then incubated with different concentrations of WTD (0.2, 1, and 5 μg/mL) for 24 h. Cell viability was determined by 3-(4, 5-dimethyl-2-thiazolyl)-2, 5-diphenyl-2 H-tetrazolium bromide (MTS) method using Cell Titer 96® AQueous One Solution Cell Proliferation Assay (Promega) according to the manufacturer's instructions. The experiments were carried out thrice in triplicate measurements.
MH7A (5 × 104 cells/mL) were seeded in 96-well plates and incubated in serum-free sterile RPMI Medium Modified (supplemented with 100 U/mL penicillin and 80 U/mL streptomycin) for 24 h. Cells were then incubated in the presence of WTD (0.2, 1, and 5 μg/mL) for 24 h.
Scratch healing assay
HUVECs were seeded in a 48-well plate (5 × 104 cells in 100 μL H-DMEM per well). Cells were incubated overnight, yielding confluent monolayers for wounding. Wounds were made using a pipette tip and photographs taken immediately (time zero). After washing with PBS, 100 μL of sterile H-DMEM media (supplemented with 5% FBS, 100 U/mL penicillin, and 80 U/mL streptomycin) with or without VEGF165 (20 ng/mL) and different concentrations of WTD (0.2, 1, and 5 μg/mL) were added to the wells. Twelve hours later, photographs were taken again. The distance migrated by the cell monolayer to close the wounded area during this time was measured. Results were expressed as a migration index, that is, the distance migrated by WTD treated relative to the distance migrated by control cells. Experiments were carried out in triplicate and repeated at least thrice.
Transwell migration assay
The migration of HUVECs to MH7A was measured by the transwell chamber migration assay. For MH7A-released chemoattractant induction, MH7A cells were seeded in a 24-well plate (1 × 105 cells in 600 μL of H-DMEM media per well). The same volume of H-DMEM media without cells was added to control wells. After 24 h, inserts with a polycarbonate membrane (pore size of 8 μm) (SPL Life Sciences, Pochun, South Korea) were installed in the plate. HUVECs were seeded in the upper chambers (5 × 104 cells/mL, 200 μL/well of H-DMEM media containing WTD (0.2, 1, and 5 μg/mL). The chamber was incubated at 37°C for 20 h. Nonmigrating cells in the upper chamber were attentively removed with cotton swabs, and cells on the lower surface of the membrane were fixed with 4% paraformaldehyde and stained with crystal violet solution. The total numbers of migrated cells were then counted in five randomly selected fields for each insert (magnification × 200) using optical microscopy. All experiments were done in triplicate.
Cell adhesive assay
HUVECs (5 × 104 cells/mL) were seeded in fibronectin (FN, 20 mg/L; Corning)- or bovine serum albumin (10 mg/mL, used as negative control)-coated 96-well plates and, respectively, incubated in sterile H-DMEM medium (supplemented with 5% FBS, 100 U/mL 1 penicillin, and 80 U/mL 1 streptomycin) for 24 h. Cells were then incubated with or without VEGF165 (20 ng/mL) and with different concentrations of WTD (0.2, 1, and 5 μg/mL) for 24 h. After treatment, cells were washed twice with PBS and 200 μL of sterile H-DMEM medium containing 5% FBS and 10% (v/v) MTS reagent was added to the cells. All absorbances at 490 nm were measured with a microplate reader. Results were expressed as cell adhesiveness. All experiments were done in triplicate.
Cell invasion assay
The cell invasion assay was performed using a transwell chamber. The upper surfaces of the transwell inserts were precoated with Matrigel (1.25 mg/mL, 20 μL/well) for 45 min at 37°C. The bottom chamber of the apparatus contained 600 μL of culture medium with or without VEGF165 (20 ng/mL for HUVECs). HUVECs (1 × 104 cells/well) were added to the upper chamber and incubated in normal growth medium with or without various concentrations of WTD (0.2, 1, and 5 μg/mL). After 24 h of incubation at 37°C and 5% CO2, noninvasive cells on the upper membrane surfaces were removed by wiping with cotton swabs. The cells were fixed and stained with crystal violet solution. Cell invasion was quantified by counting the cells on the lower surface under a phase-contrast microscope at 200 × magnification. The average number of migrating cells was counted in five random fields. Three independent assays were performed.
Tube formation assay
To examine the inhibitory effect of WTD on HUVEC tube formation, the tube formation assay was performed as described previously. 27 Matrigel (10 mg/mL) was plated in 48-well culture plates and allowed to polymerize at 37°C in 5% CO2 humidified for 30 min. HUVECs were removed from culture, trypsinized, and resuspended in sterile H-DMEM medium (supplemented with 5% FBS, 100 U/mL penicillin, and 80 U/mL streptomycin). HUVECs (6 × 104 cells/mL) were added to each chamber, followed by addition of various concentrations of WTD (0.2, 1, and 5 μg/mL) with or without VEGF165 (20 ng/mL), and then incubated for 6 h at 37°C in 5% CO2. After incubation, the capillary-like tube formation of each well in the culture plates was photographed using phase-contrast microscopy. Quantitation of the anti-angiogenic activity of WTD on tube formation was by counting the number of branch points. All experiments were done in triplicate.
Enzyme-linked immunosorbent assay
MH7A/HUVECs were treated with or without TNF-α/VEGF165 plus WTD (0.2, 1, and 5 μg/mL, respectively) for 24 h. The supernatants and cells were collected. The expression levels of VEGF, IL-1β, IL-17, transforming growth factor-β (TGF-β), platelet derived growth factor (PDGF), placenta growth factor (PlGF), angiopoietin (Ang) I, and Ang II in supernatants of MH7A, and VEGFR2 in HUVEC cells with different treatments: control—normal cultured cells; TNF-α/VEGF165—TNF-α/VEGF165 induced cells; and WTD groups—cells treated with various concentrations of WTD (0.2, 1, and 5 μg/mL)] were detected by enzyme-linked immunosorbent assay (ELISA; R&D) according to the manufacturer's protocol. All experiments were done in triplicate.
Western blot analysis
HUVECs were pretreated with WTD for 2 h and subsequently stimulated with VEGF165 (20 ng/mL) for 0.5 h. Briefly, total protein was extracted using a lysis buffer. Cell lysates (50 μg) were loaded and separated by using 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, and then blotted onto a polyvinylidene fluoride membrane. Membranes were blocked at room temperature for two hours and incubated overnight at 4°C with primary antibodies of VEGFR2, p-VEGFR2, AKT, p-AKT, ERK1/2, p-ERK1/2, JNK, p-JNK, p38, and p-p38 (1:1000; Cell Signaling Technology). Subsequently, membranes were incubated for 1 h at room temperature with secondary antibodies conjugated with horseradish peroxidase. The expression of GAPDH was used as an internal standard. All experiments were done thrice.
Statistical analysis
The software of SPSS version 11.0 for Windows (SPSS, Inc., IL) was used for statistical analysis. Continuous variables were expressed as
Results
WTD inhibits angiogenesis in synovium tissue of joints in CIA rats
Angiogenesis has been considered a critical step in the progression of chronic arthritis, as well as an early determinant in the development of RA. In our previous studies, WTD significantly decreased the arthritis score and arthritis incidence, and inhibited inflammation, pannus formation, and cartilage and bone destruction of inflamed joints in CIA and AA rats. 16,18,19 In this study, to assess the potential mechanism through which WTD exerts its anti-arthritic action, H&E staining was examined for the presence of vascular structures. As shown in Figure 1A, the extent of vascular formation was indeed inhibited by the dose of 3.8 g/kg WTD, as the score (0.5 ± 0.22) of mean synovial vascularity inside the construct decreased compared with that (2.5 ± 0.48) of model group. CD31, a marker of blood vessels, was stained in synovium tissue of joints by immunohistochemistry. As shown in Figure 1B, a significant amount of CD31 staining was present in synovium tissue of inflamed joints from model rats, and this was markedly attenuated in WTD-treated rats. In addition, WTD affected the morphology of the newly formed immature microvessels, identified by staining for CD31, which were not covered by αSMA-positive perivascular cells (Fig. 1C). Quantitative evaluation further revealed that doses of 0.95–3.8 g/kg WTD significantly reduced the number of immature CD31+/αSMA− vessels (Fig. 1D), but not mature CD31+/αSMA+ vessels (Fig. 1E), and decreased the total number of blood vessels (Fig. 1F) in synovium tissues of inflamed joints of CIA rats by immunofluorescence analysis. These results suggest that WTD has a potent anti-angiogenic activity in vivo.
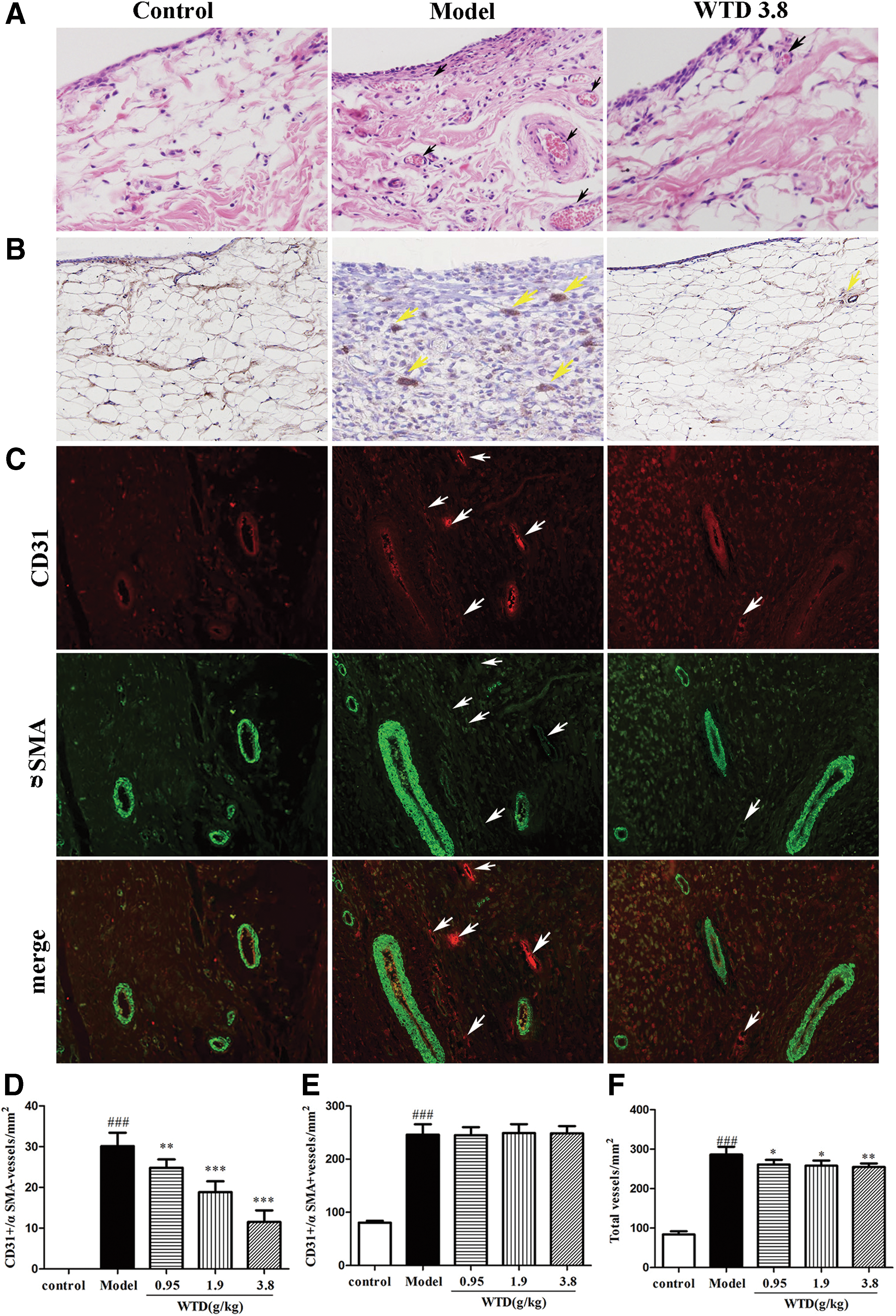
WTD decreases the vessel density in synovial membrane of inflamed joints in CIA rats.
WTD suppresses VEGF165-induced microvessel sprout formation ex vivo and in vivo angiogenesis in chick embryo
The aortic ring assay, an ex vivo assay, mimics several stages of angiogenesis, including endothelial cellular proliferation, migration, tube formation, microvessel branching, and perivascular recruitment. As shown in Figure 2A–D, VEGF165 significantly triggered endothelial cell migration and microvessel sprouting, leading to the formation of a complex microvessel network emerging from the aortic rings and growing outward; by contrast, WTD inhibited microvessel sprouting in a dose-dependent manner, suggesting that WTD suppressed VEGF165-induced microvessel sprout formation ex vivo.

WTD suppresses VEGF165-induced vessel sprout formation ex vivo and inhibits in vivo angiogenesis in chick embryo.
The anti-angiogenesis effect of WTD was investigated in vivo in CAM assay using chicken embryos. Normal vasculature pattern in the untreated CAMs with clear visualization of primary, secondary, and tertiary vessels and dendritic branching pattern is shown in Figure 2E and F (arrows). Whereas, WTD-treated (0.2, 1 and 5 μg/mL) CAM displayed distorted architecture (arrow heads) in the vasculature. The number of blood vessels was reduced drastically in WTD-treated CAMs. WTD inhibited the formation of new blood vessels significantly.
WTD inhibited the VEGF165-induced migration, invasion, adhesion, and tube formation of HUVECs
In angiogenesis, ECs migrate in response to several chemotactic factors. 28 Therefore, we attempted to elucidate whether WTD affects VEGF165-induced EC migration and MH7A cell-directed EC chemotaxis. As shown in Figure 3A and B, WTD suppressed VEGF165-induced wounding migration of HUVECs in a concentration-dependent manner. Moreover, WTD strongly inhibited the MH7A cell-directed chemotaxis of HUVECs in a dose-dependent manner, determined by using a Transwell chamber (Fig. 3C, D).
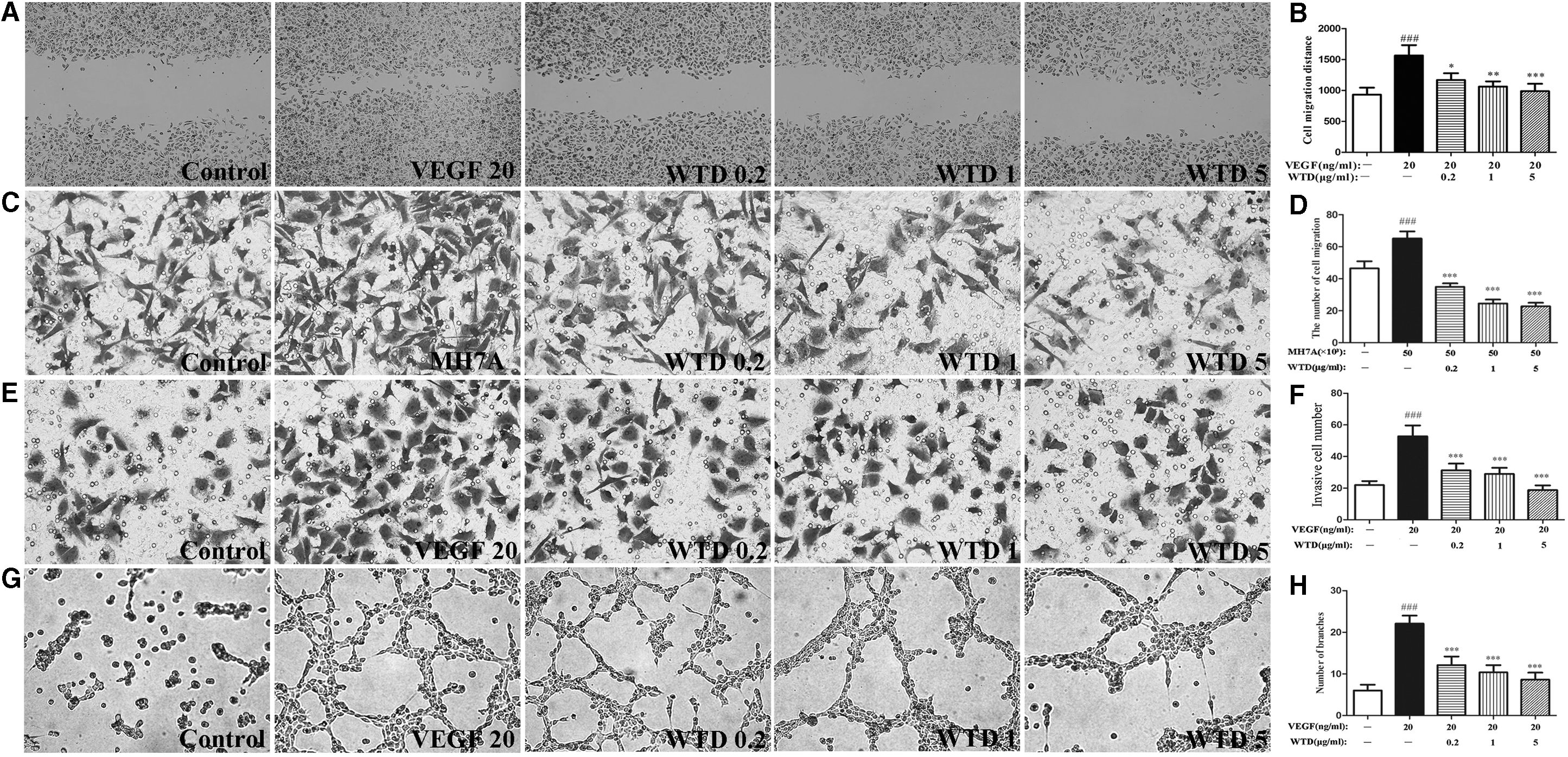
WTD inhibits VEGF165-/MH7A-induced HUVEC migration, invasion, and tube formation.
Transwell chamber (precoated with Matrigel) assays were also performed to determine if WTD affects endothelial cell invasion. Numerous HUVECs migrated to the underside of the filters in the Transwell chambers after stimulation of the cells with VEGF165. However, the number of invasive cells was significantly reduced in the presence of WTD in a dose-dependent manner (Fig. 3E, F), indicating a potent inhibitory effect of WTD on VEGF165-induced endothelial cell invasion ability.
Inhibitory effects of WTD on cell adhesiveness of HUVECs were determined by adhesive assay. WTD at a concentration ranging from 0.2 to 5 μg/mL significantly suppressed the cell adhesiveness of HUVECs (data not shown).
Furthermore, the anti-angiogenic effects of WTD on HUVEC tube formation, which mimics angiogenesis, were detected by a tube formation assay. Robust and complete tubular-like structures were formed in the presence of VEGF165 when HUVECs were plated on Matrigel. WTD disrupted tube formation in a concentration-dependent manner compared with the tube formation of HUVECs induced by VEGF165 (Fig. 3G, H).
WTD reduces the expression levels of proangiogenic mediators
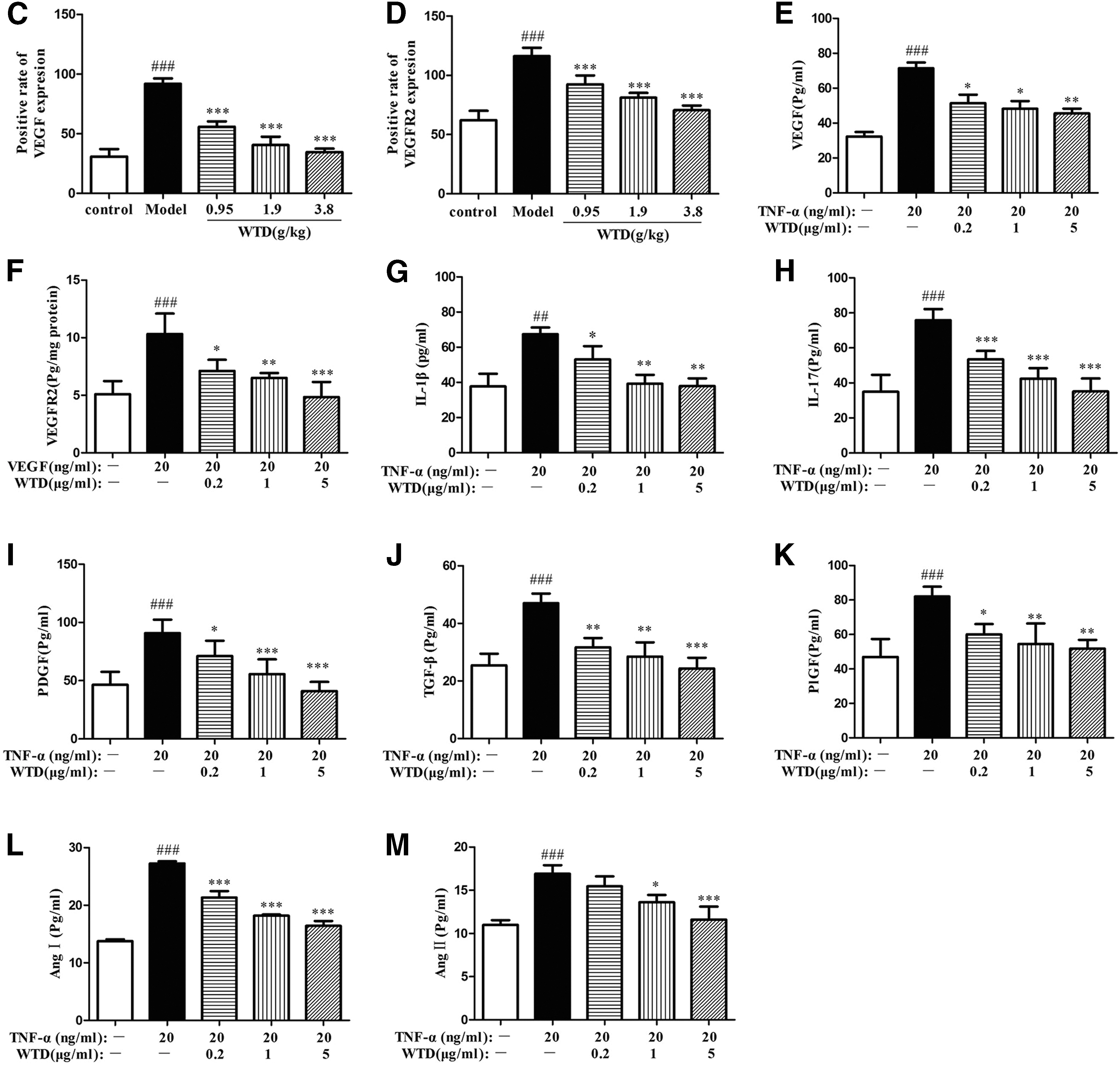
To investigate the mechanisms by which WTD suppressed the angiogenesis in RA, we detected the expression levels of angiogenic activators, including VEGF, VEGFR2 in synovium of rats by immunohistochemistry, IL-1β, IL-17, VEGF, PDGF, TGF-β, PlGF, Ang I, and Ang II in TNF-α-induced MH7A, and VEGFR2 in VEGF165-induced HUVECs by ELISA. WTD strongly reduced the VEGF (Fig. 4A, C) and VEGFR2 (Fig. 4B, D) expression in synovium of CIA rats in a dose-dependent manner. The expression levels of VEGF (Fig. 4E) in TNF-α-induced MH7A cells and VEGFR2 (Fig. 4F) in VEGF165-induced HUVECs treated with WTD were also significantly depressed. In addition, WTD significantly inhibited the expression of IL-1β (Fig. 4G), IL-17 (Fig. 4H), PDGF (Fig. 4I), TGF-β (Fig. 4J), PlGF (Fig. 4K), Ang I (Fig. 4L), and Ang II (Fig. 4M) in TNF-α-induced MH7A cells.
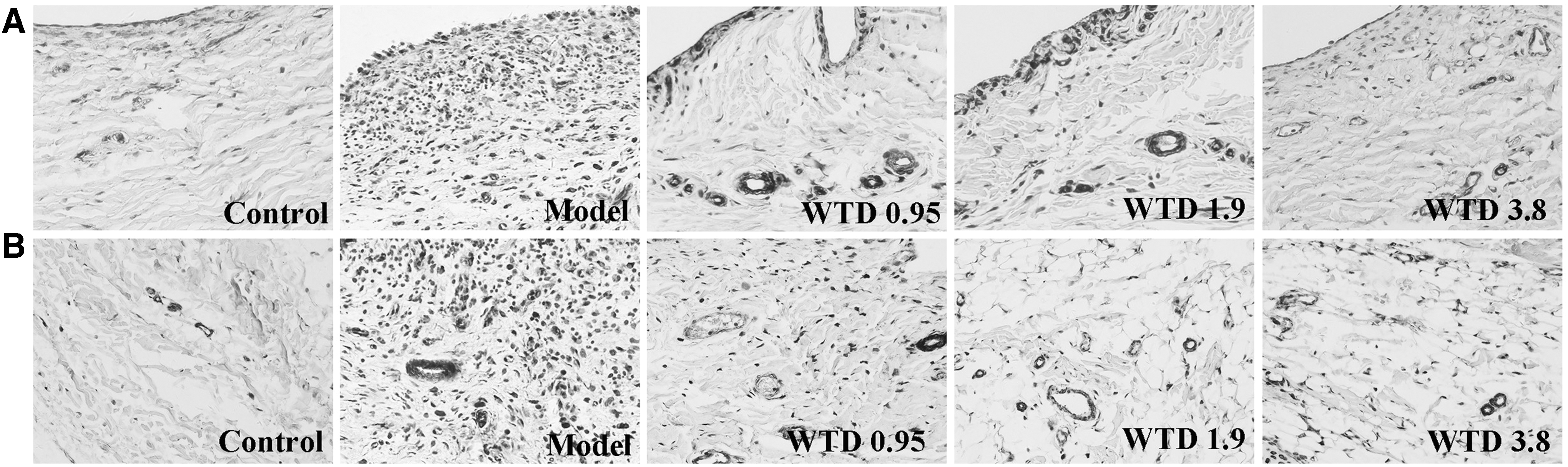
WTD reduces the expression levels of various angiogenic activators in collagen-induced rats and in MH7A/HUVECs cells. For in vivo assays, collagen-induced rats were orally administered WTD (0.95, 1.9, and 3.8 g/kg, respectively) for 28 days from the day of first immunization. At the end of the experiment, joints were obtained and stained by immunohistochemistry.
WTD decreases VEGF165-induced autophosphorylation of VEGFR2
The effect of WTD on VEGF165-induced tyrosine autophosphorylation of VEGFR2 was evaluated to examine the potential active sites of WTD in the signaling pathways. WTD decreased VEGF165-induced tyrosine autophosphorylation of VEGFR2 for 0.5 h in a dose-dependent manner, but not the total level of VEGFR2, suggesting that the prevention of VEGFR2 autophosphorylation contributed to the inhibitory effect of WTD on VEGF165-induced angiogenesis (Fig. 5A).
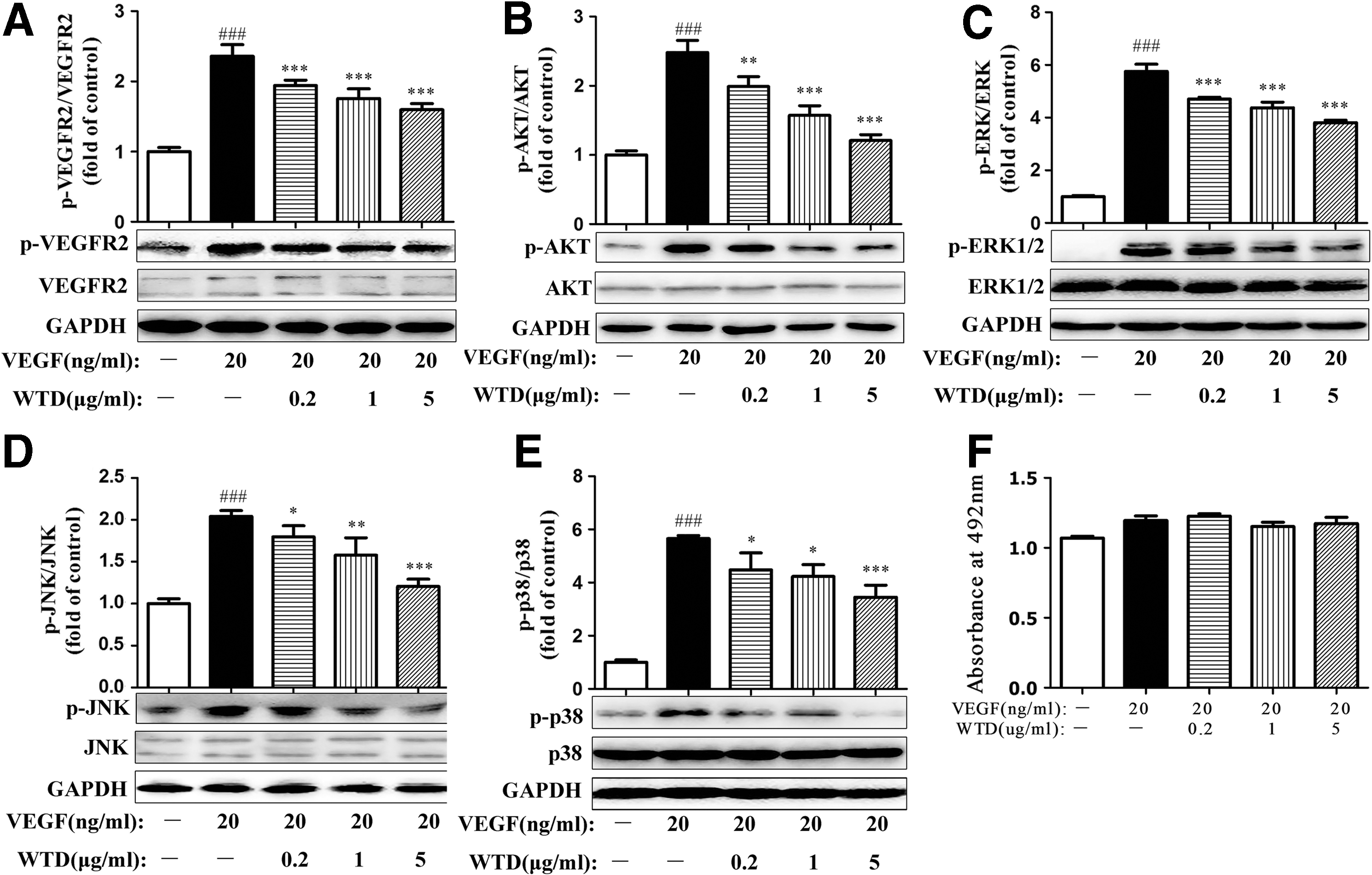
WTD decreases the autophosphorylation of VEGFR2 and the activation of key proangiogenic molecules involved in the VEGFR-2-mediated signaling pathway in VEGF165-induced HUVECs. HUVECs were starved in 10% FBS medium for 48 h, pretreated with WTD for 2 h, and then stimulated with VEGF165 (20 ng/mL) for 0.5 h before collection. WTD inhibited the phosphorylation of VEGFR2
WTD attenuates the activation of the VEGF165-induced VEGFR2-mediated signaling pathway
The expression of several key kinases involved in the VEGFR2-mediated signaling pathway was studied to further delineate the mechanism underlying the anti-angiogenic effects of WTD. HUVECs were pretreated with WTD for 2 h and subsequently stimulated with VEGF165 (20 ng/mL) for 0.5 h. As shown in Figure 5B–E, the expression levels of p-AKT, p-ERK1/2, p-JNK, and p-p38 were markedly increased by VEGF165 treatment. WTD greatly attenuated the phosphorylation of these kinases, indicating that the effect of WTD on VEGF165-induced angiogenesis was inhibited by the downregulation of VEGFR2-mediated PI3K, AKT, ERK1/2, JNK, and p38.
Next, we examined whether the above suppressive effect of WTD was due to its cytotoxicity. When confluent HUVECs were treated with WTD and/or VEGF165 for 24 h, the cytotoxicity was monitored by the MTS assay. Our results showed that WTD did not exert any cytotoxic effects on HUVECs (Fig. 5F) under the experimental conditions used in this study, suggesting that WTD might specifically suppress the above function of HUVECs.
Discussion
We have previously demonstrated that WTD is effective in the treatment of CIA and AA, with the decrease of arthritis score and arthritis incidence, and inhibition of inflammation, pannus formation, and cartilage and bone destruction of inflamed joints. 16,18,19 It is of great importance to understand its mode of actions and potential drug targets to effectively use WTD for the therapy of RA. To further clarify the mechanisms of WTD acting on RA, we, in this study, discovered and demonstrated the anti-angiogenic effect of WTD in vivo, ex vivo, and in vitro.
Angiogenesis as a critical component of disease progression in RA involves in the pannus formation and maintenances the infiltration of synovial membrane. RA synovium contains a significant fraction of immature blood vessels. 23 Progression of the disease increases the presence and density of immature, but not mature vessels and only immature vessels are depleted in response to anti-TNF-a therapy. 23,24 In the study, our results indicated that the immature blood vessels, but not mature vessels in synovial membrane tissues of arthritic joints of CIA rats treated with WTD were significantly inhibited. In addition, WTD demonstrated potent inhibitory effect on sprouting of microvessel from rat aorta and neovascularization in fertilized chicken embryo. Therefore, these results indicate that WTD has potent anti-angiogenic activity both in vivo and ex vivo. Glycyrrhiza Radix Preparata, Ephedrae Herba, Paeoniae Radix Alba, Aconiti Radix Cocta, and Astragali Radix are the compositions of WTD. It has been reported that Astragali Radix promotes angiogenesis in vitro using HUVECs, 29 in rat animal models of wound healing, 30 or in zebrafish models, 31,32 while others suggest that Astragali Radix has an anti-angiogenic effect 33 or decreases VEGF levels. 34 Besides, the effect of each of other four herbs of WTD on angiogenesis is still unknown, too. Therefore, how the five herbs contribute to the anti-angiogenic effect of WTD in RA is the next aim to investigate.
New vessel formation (angiogenesis) involves multiple steps, including endothelial cell migration, invasion, adhesiveness, tube assembly, and remodeling. 35 In this study, we systematically investigated the potential effects of WTD on these key processes of VEGF165-induced endothelial cell viability. During the process of angiogenesis, a single layer of endothelial cells lining the inside of blood vessels divides and breaks off from the vessel membrane, forming the tubes, and eventually become new capillaries. 36 WTD effectively inhibited the key aspects of angiogenesis in endothelial cells, such as migration and invasion. After cells migrate into the perivascular space in blood vessels, HUVECs undergo differentiation and the cells assume a characteristic shape to facilitate the adherence among the cells to form a lumen or tube-like structure. WTD remarkably inhibited the formation of such tube-like structures in a dose-dependent manner. As the process of tube formation highly relied on cell-cell adhesion, 37 we also found that the cell adhesiveness of HUVECs could be significantly reduced in the presence of WTD. Therefore, these results indicate that WTD inhibits angiogenesis by inhibiting the key angiogenic processes in vitro. Further investigation revealed that, the anti-angiogenic efficacy of WTD was shown to be mediated by interfering with the endothelial cell function.
In RA, synovial angiogenesis is closely associated with chronic inflammation. Moreover, the hypertrophy, migration, invasion, and adhesion of synoviocytes are critical for inflammation, pannus, and joint destruction of RA. Thus, the effect of WTD on the hypertrophy, migration, invasion, and adhesion of RA fibroblast-like synoviocytes (RA-FLS) is also observed in the study. The results were similar with that of WTD on HUVECs (data not shown). It suggests that WTD may inhibit RA inflammation by intervening in these processes, and the anti-angiogenic effect of WTD might be related to the anti-inflammation. On the other hand, WTD consists of five Chinese medicinal herbs: Glycyrrhiza Radix Preparata, Ephedrae Herba, Paeoniae Radix Alba, Aconiti Radix Cocta, and Astragali Radix. Each herb of WTD has been reported to play an anti-inflammatory role separately. 38 –44 Taken the five herbs together, the anti-inflammatory effect of them may be contributable to the anti-angiogenic effect of WTD in RA too.
A great number of proangiogenic factors, including VEGF, HIFs, TNF-α, IL-1β, IL-17, TGF-β, PDGF, PIGF, Ang I, Ang II, and MMPs, govern angiogenesis in RA. 9 Among these, VEGF is the most potent angiogenic regulator, which is produced in the synovium in response to proinflammatory cytokines such as TNF-α, IL-1β, IL-17, HIFα, and TGF-β. VEGF acts in angiogenesis by inducing EC proliferation, migration, and tube formation. 11,45 –47 VEGF mediates endothelial proliferation, while inhibiting pericyte and SMC development, a process instead dependent on PDGF signaling. 48 –50 Other factors like Ang I and Ang II play a crucial role in the control of vessel stabilization and regression. 51 In this study, to observe the mechanism of the anti-angiogenic effect of WTD, these angiogenic mediators were investigated. Our results demonstrated that WTD significantly reduces the expression level of VEGF in synovium of CIA rats, and the TNF-α-induced upregulation of VEGF, IL-1β, IL-17, PDGF, TGF-β, PIGF, Ang I, and Ang II in MH7A. These findings were consistent with our previous results, when WTD markedly inhibited the expression of VEGF, IL-1β, and HIFα in sera of CIA rats and in HFLS-RA, and MMPs-1 and -13 in the joint of CIA rats and HFLS-RA. 16 Taken together, WTD inhibits angiogenesis by downregulating proangiogenic factors, including VEGF, IL-1β, IL-17, PDGF and TGF-β, PIGF, Ang I, and Ang II. Besides, it is thought that synovial tissue hypoxia mediated by HIFα promotes angiogenesis, in part, through an increased expression of proangiogenic factors such as VEGF, while prolyl hydroxylase domain enzyme (PHD-2) and glucose-6-phosphate isomerase (G6PI) can regulate hypoxic responses in RA. 52,53 We will investigate the effects of WTD on PHD-2 and G6PI in the next study.
VEGF elicits biological effects by cooperation with the transmembrane receptor VEGFR2, 54 and the high-affinity combination of the two molecules regulates endothelial cell proliferation, migration, and tube formation. 55 To explore the molecular mechanism of the anti-angiogenic effect of WTD, we assessed the ability of WTD on the expression and activation of VEGFR2. Our data showed that WTD reduces the expression level of VEGFR2 in synovium of CIA rats and VEGF165-induced upregulation of VEGFR2 in HUVECs for 24 h. Meanwhile, WTD significantly blocked the kinase activity of VEGFR2 by downregulating VEGF-induced autophosphorylation of VEGFR2 expression in HUVECs for 0.5 h dose dependently. These results suggest the inhibitory effect of WTD on angiogenesis by targeting VEGFR2.
To further elucidate the anti-angiogenic mechanism of WTD, we examined the downstream signaling mediators of VEGFR2 related to angiogenesis. The activation of VEGFR2 by VEGF165 stimulation subsequently results in the activation of several downstream pathways, including PI3K/AKT, ERK-MAPK, p38-MAPK, and JNK-MAPK, which can positively regulate proangiogenic signals. 56,57 The results of our study suggest that WTD may downregulate the VEGF165-induced phosphorylation of multiple downstream signaling components of VEGFR2, AKT, ERK, JNK, and p38. Such downregulation of these protein molecules facilitates the inhibition of migration, invasion, and adhesion of VEGF165-induced endothelial cells in vitro. The latest study has shown that other downstream signals such as nocanonical nuclear factor-κB and Notch-1 also mediate VEGF-induced angiogenesis. 58,59 In the next study, we will further investigate the effect of WTD on these two pathways.
In conclusion, the above findings are summarized in Figure 6. Our results indicate that WTD significantly reduced synovial angiogenesis in CIA rats, and inhibited sprouting of microvessel in rat aorta and neovascularization in chick embryo. This reduction may be attributable to the inhibition of endothelial cell migration, adhesion, invasion, and tube formation. Furthermore, WTD exerts anti-angiogenic effects by downregulating angiogenic activators and suppressing the VEGFR2-mediated signaling pathway, which plays multiple roles in the regulation of angiogenesis. These findings suggest for the first time that WTD possesses the anti-angiogenic effect in RA in vivo, ex vivo, and in vitro by interrupting the targeting of VEGFR2 activation. Therefore, WTD may act as a potential therapeutic agent for RA treatment through its anti-angiogenic effects in the synovium.
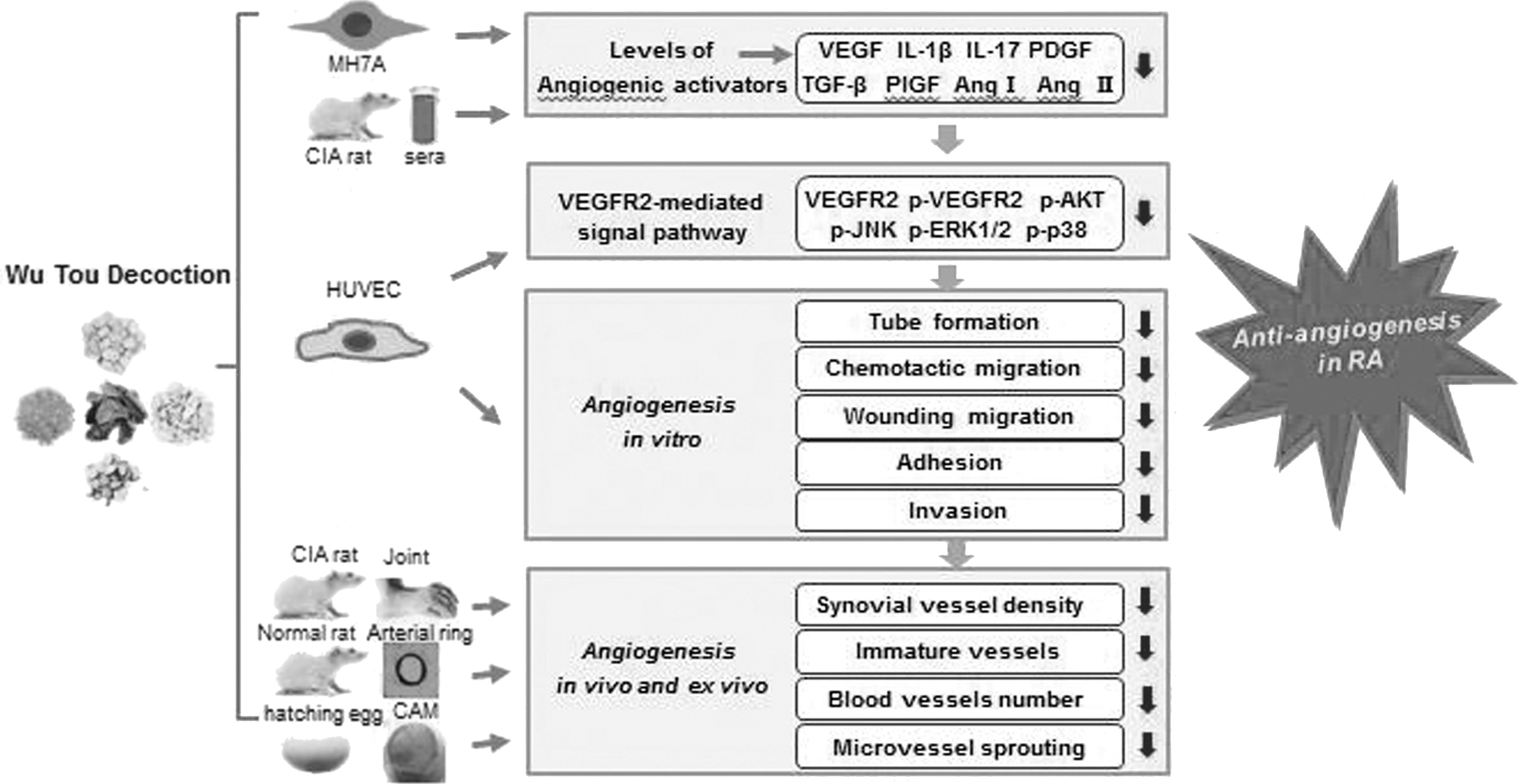
Model depicting the proposed mechanism by which WTD exerts anti-angiogenic effect in experimental arthritis. Target-derived inflammatory cytokines such as IL-1β, IL-17, PDGF, TGF-β, PlGF, Ang I, and Ang II could be secreted out from human fibroblast-like synoviocytes-RA induced by TNF-α, then stimulating VEGF secretion, subsequently activated VEGFR2 and the downstream of AKT and MAPK signal pathway in HUVECs. Consequently, the activated VEGFR2-mediated signal pathway can lead to HUVEC migration, adhesion, invasion, and tube formation. Black arrows indicate that an inhibition or a downregulation was observed. CAM, chorioallantoic membrane; CIA, collagen-induced arthritis.
Footnotes
Acknowledgment
This work was supported by grants from the National Major Scientific and Technological Special Project for “Significant New Drugs Development” (No. 2018ZX09731019-012) and the National Natural Science Foundation of China (No. 81630107, 81573878, 81374068).
Author Disclosure Statement
No competing financial interests exist.