
Abstract
Advancing age is the biggest risk factor for development for the major life-threatening diseases in industrialized nations accounting for >90% of deaths. Alzheimer's dementia (AD) is among the most devastating. Currently approved therapies fail to slow progression of the disease, providing only modest improvements in memory. Recently reported work describes mechanistic studies of J147, a promising therapeutic molecule previously shown to rescue the severe cognitive deficits exhibited by aged, transgenic AD mice. Apparently, J147 targets the mitochondrial alpha-F1-ATP synthase (ATP5A). Modest inhibition of the ATP synthase modulates intracellular calcium to activate AMP-activated protein kinase to inhibit mammalian target of rapamycin, a known mechanism of lifespan extension from worms to mammals.
Introduction: Discovery of J147
O
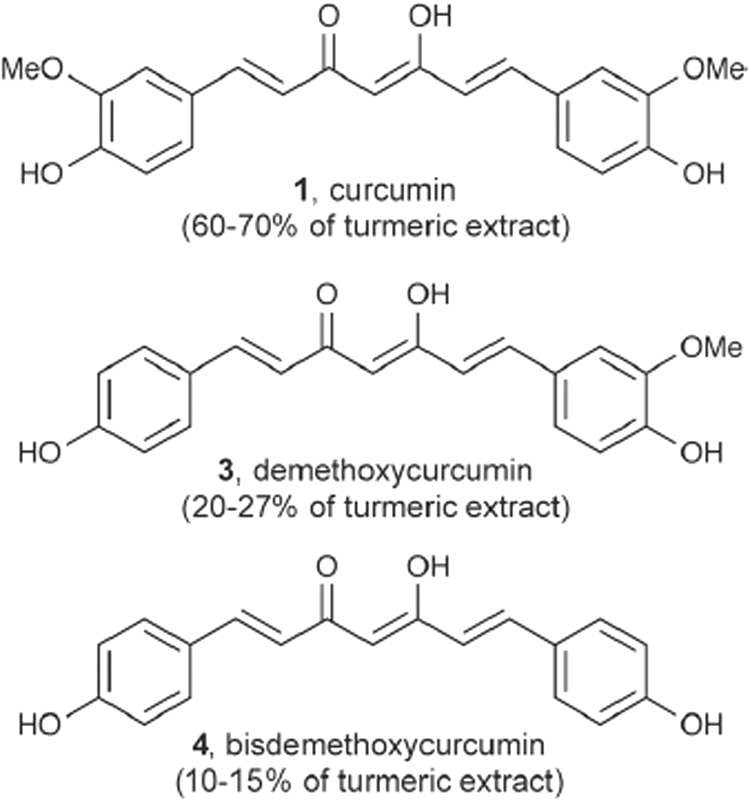
Curcumin-related constituents from extracts of Curcuma longa.
Additional medicinal chemistry produced J147 (Fig. 2), 11 a potent, orally active, neurotrophic molecule that facilitates memory in normal rodents, and prevents the loss of synaptic proteins and cognitive decline when administered preventatively to 3-month-old APP/PSEN1ΔE9 Alzheimer disease model mice for 7 months 12 or given therapeutically to 3–20-month-old aged mice. 13 Administration of J147 augmented CNS levels of brain-derived neurotrophic factor and nerve growth factor, enhanced long-term potentiation, preserved synaptic proteins, reduced markers of oxidative stress and inflammation, and reduced amyloid plaques with lower levels of soluble Abeta1-42 and Abeta1-40. However, the molecular target of J147 was elusive until now. 14
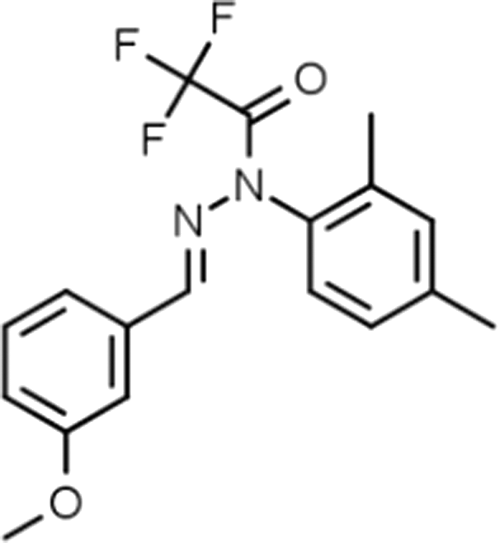
Molecular structure of J147.
J147 Targets Mitochondrial ATP Synthase
Goldberg et al. 14 determined the protein target of J147 using drug affinity responsive target stability (DARTS), a method that relies on the protection against proteolysis conferred on the target protein by interaction with a small molecule. 15 Separately, Goldberg et al. performed a pull-down experiment in which a biotinylated derivative of J147, BJ147, is bound to cell extracts, interacting peptides are captured with streptavidin, and these are analyzed by LC/MS/MS to identify coprecipitating proteins. The most highly enriched protein in both the DARTS and affinity precipitation experiments was ATP5A, the alpha subunit of the mitochondrial ATP synthase (Fig. 3). J147 incubated with mitochondria isolated from bovine heart exhibited potent (EC50 @ 20 nM) although only partial modest inhibition (∼24%) of ATP synthase activity. 14
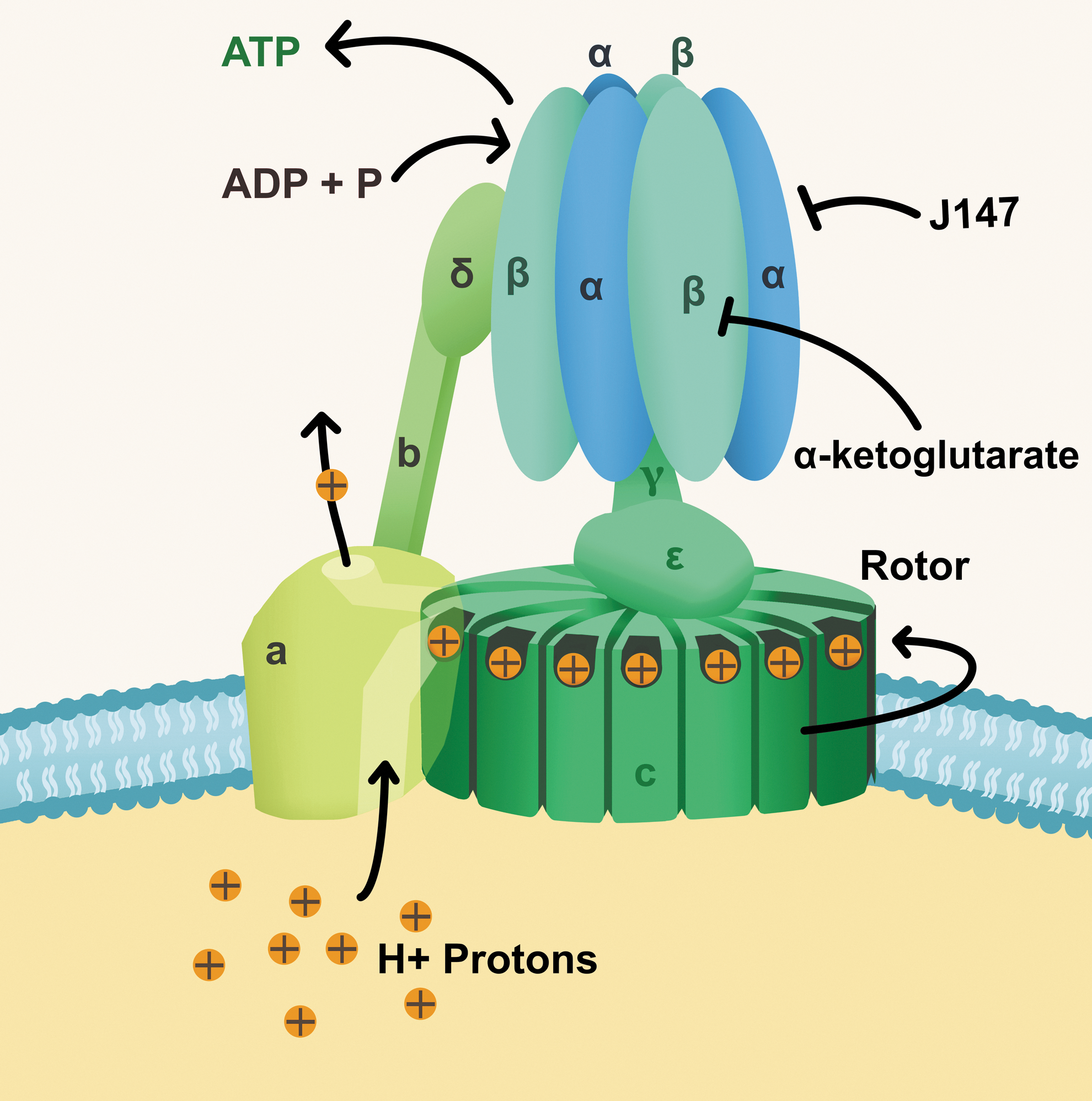
Schematic of ATP synthase, Complex V. The ATP synthase comprises two “rotary nanomotors,” each powered by a different fuel. The membrane-embedded motor, termed F0, is powered by the flow of hydrogen ions across the membrane. As the protons flow through the motor, (a) they turn a circular rotor (c) (shown in green). This rotor is connected to the second motor, termed F1. The F1 motor is a chemical motor, powered by ATP. The two motors are connected together by a stator (b) (left). The F0 motor uses the power from the proton gradient generated by other complexes of the respiratory chain to force the F1 motor to generate ATP. The vacuolar ATPase working in reverse, uses an ATP-driven motor to pump protons across a membrane. Work of Goldberg et al. demonstrated that J147 is bound to the alpha subunit of the ATP synthase. Earlier work of Chin et al. demonstrated that alpha-ketoglutarate mediated ATP synthase inhibition through the beta subunit.
It should be noted that several other mitochondrial-associated proteins, VDAC, Slc25a, and IP3R3, were also identified in the BJ147 pull-down experiments. These proteins 16,17 along with ATP5A comprise the mitochondria permeability transition (mPT) pore responsible for executing cell death programs during lethal conditions of stress through mitochondrial Ca2+ efflux and are probably activated during neurocytotoxicity. 18 How these molecules figure in the activities of J147 is presently not known, although it would be tempting to hypothesize that J147 may directly inhibit the mPT in cells experiencing neurotoxicity.
J147 Modulates Mitochondrial Bioenergetics
The mitochondrial membrane potential (Δψm) is a key measure of cellular vitality. Mitochondrial dysfunction characterized by lower Δψm correlates with reduced levels of ATP in aging and Alzheimer's dementia (AD). 19,20 Using a cationic dye assay. 21 J147 was shown to increase the Δψm, which is consistent with slower dissipation of the proton gradient by ATP synthase. Similar results were found when ATP5A levels were reduced by specific siRNA treatment. The reduced ATP synthase activity was accompanied by augmented production of superoxide, which typically occurs upstream of ATP synthase (Complex V) at Complexes I and III. 14 These findings are consistent with the “retrograde, ROS-mediated prosurvival response” previously described by Formentini et al. 22
Perhaps most importantly, cellular levels of ATP increased by 4 hours post-treatment with J147 although with maximal effect at a concentration of 100 nM. 14 Why do ATP levels increase, when a key component of ATP production is inhibited? Increased ATP levels are associated with cell survival. Although not discussed by Goldberg et al., one possibility is that another capability of ATP synthase is involved. ATP synthase is fully capable of the hydrolyzing ATP as well as creating ATP. Interestingly, it has been reported that embryonic neuronal cell types, unlike adult neurons, have ATP synthase activity that tends to consume rather than create ATP. 23 In fact, the HT22 cell line used by Goldberg et al. to measure ATP levels is known to be resistant to glutamate-induced oxidative stress because of reverse activity of ATP synthase. 24 It should be noted that Goldberg et al. also did observe increase of ATP in drosophila heads treated with J147. The combination of unimpeded ATP production by glycolysis, increased ATP synthesis by AMP-activated protein kinase (AMPK)-related pathways, reduction of ATP hydrolysis by ATP synthase, and cell type specificity may explain the apparent anomaly and raise questions about how generally does J147 increase ATP levels.
J147 Modulates Resting Ca2+ Homeostasis to Activate the AMPK/Mammalian Target of Rapamycin Axis
J147 treatment augments cytosolic Ca2+ levels with subsequent activation of Ca2+/Calmodulin-mediated activation of CamKinaseKinase 2. This contributes to activation of AMPK in the brain 25 by J147 despite the increased ATP levels, which typically would inhibit AMPK. Studies with inhibitors demonstrate that CA+ signaling is required for the observed AMPK activation by J147. This calcium flux changes with the mitochondrial membrane potential (Δψm) that is maintained by the proton flux and activity of the ATP synthase. 26 Goldberg et al. 14 do not understand how J147 inhibition or siRNA knockdown of ATP5A leads to compartmental changes in the observed levels of calcium.
Common Mechanisms: J147, Alpha-Ketoglutarate, and ATP5A Modulate AMPK/Mammalian Target of Rapamycin Signaling
AMPK activation is known to lead to mammalian target of rapamycin (mTOR) inhibition. The mTOR is the catalytic subunit of two structurally distinct complexes: mTORC1 and mTORC2 that integrate both intracellular and extracellular signals of cell metabolism, growth, proliferation, and survival. mTORC1 is composed of mTOR, regulatory-associated protein of mTOR (Raptor), mammalian lethal with SEC13 protein 8 (mLST8), and the noncore components PRAS40 and DEPTOR. This complex functions as a nutrient/energy/redox sensor and controls protein synthesis. 27 The activity of mTORC1 is regulated by insulin, growth factors, phosphatidic acid, certain amino acids, and their derivatives (e.g., l-leucine and β-hydroxy β-methylbutyric acid), mechanical stimuli, and oxidative stress. Rapamycin inhibition of mTORC1 appears to provide most of the beneficial effects of the drug (including life-span extension in animal studies). In contrast, mTORC2 inhibition produces reduced glucose tolerance and insensitivity to insulin.
AMPK is activated through phosphorylation of threonine 172 on the alpha-subunit. This lowers activity of some ATP-consuming pathways while promoting ATP synthesis through others such as fatty acid oxidation. Goldberg et al. showed that J147 mediates a time-dependent activation of AMPK followed by increased phosphorylation of raptor at Ser792, known to be critical for inhibition of mTOR. Then evidence was presented that downstream of mTOR/raptor two important lifespan extension pathways were engaged: (1) S6 kinase with reduced protein translation and (2) acetyl-CoA carboxylase (ACC1) with increased beta-oxidation of fatty acids, which should lead to overall ATP production, and less fatty acid synthesis.
14
ATP Synthase Inhibition Is Neuroprotective
Formentini et al. generated a conditional mouse model expressing a mutant form of human ATPase inhibitory factor 1 (hIF1). They reported that reduced ATPase activity in these mice protected CNS neurons from excitotoxic damage. 28 Goldberg et al. corroborated these results, showing that hIF1-mediated protection in cell culture neurotoxicity assays with the HT22 transfected with a constitutively activated hIF1 and that ATP5A siRNA-mediated knockdown is neuroprotective. 14
Aging: Lifespan Effects
Although it may be counter-intuitive that reduced ATP synthase activity can promote lifespan extension, several studies support this finding. Although complete loss of mitochondrial function is detrimental, partial suppression of the electron transport chain has been shown to extend the lifespan of C. elegans. 29 –32 Furthermore, Chin et al. 33 demonstrated that the tricarboxylic acid (TCA) metabolite alpha-ketoglutarate (α-KG) extends the lifespan of worms by inhibiting the beta subunit of ATP synthase and TOR, the worm equivalent of mTOR, although in their experiments ATP levels decrease, rather than increase, perhaps suggesting that ATP levels are less relevant than what Goldberg et al. hypothesize. Goldberg et al. show that J147 extended median lifespan of drosophila up to 12.5%, 14 a result similar to that reported by Sun et al. for women with normal TOR activity. 34 It should be noted that a 12.5% increase in drosophila lifespan is modest and that lifespan increase was not explored in any other model system, including mice.
Senescence-accelerated mouse-prone 8 (SAMP8) mice are a spontaneous mutant inbred strain that display a phenotype of accelerated aging and AD-like neurological changes with age. 35 The key genetic changes that lead to accelerated aging have not yet been identified in this model. The method of Rangaraju et al. 36 measuring transcriptional drift associated with aging in C. elegans was used to examine gene expression in the hippocampus of old SAMP8 mice. Patterns suggestive of a youthful genotype were more common in the J147-treated animals, suggesting that J147 may delay or inhibit epigenetic drift. It would be extremely useful to know whether this effect occurs in normal mice and other tissues.
Medical Implications
Because age is the greatest risk factor for development of AD, an intervention that slows aging or extends the health span is expected to delay onset of disease, providing major benefits to patients. 37,38
Health span and aging are closely linked to metabolism. Dietary restriction (DR) or calorie restriction are the “gold standards” for life extension and delay of age-related diseases across all species studied. 39 –41 A limited number of pathways controlling nutrient or energy metabolism also have longevity benefits. 42,43 Various metabolites can modulate aging 43 –45 and the TCA cycle intermediate α-KG (Fig. 3) was recently shown to extend the lifespan of adult C. elegans through inhibition of the ATP synthase beta catalytic subunit that seems similar to that described for J147 that inhibits the allosteric regulating alpha subunit. 33
Starvation-activated anaplerotic gluconeogenesis increases glutamate-linked transaminases in the liver to provide carbon derived from amino acid catabolism with elevation of α-KG levels.
α-KG levels are increased upon starvation and additional α-KG does not extend the lifespan of DR animals, indicating that α-KG is a key metabolite that mediates longevity by DR. However, there are subtle differences among these modulators of the ATP synthase. Unlike J147, α-KG inhibition of the ATP synthase leads to reduced ATP content, decreased oxygen consumption, and increased autophagy in both C. elegans and mammalian cells. However, α-KG and J147 treatment ultimately inhibits the canonical mTOR pathway.
Recent studies have highlighted a role for ATP synthase in the regulation of mTOR and lifespan extension in flies and worms 33,34 and inhibition of mTOR/TOR through activation of AMPK is a canonical longevity-associated pathway. 46 It is possible that inhibition of mTOR/TOR by J147 explains the modest increase in lifespan observed in flies and worms.
Shabalina et al. describe a mitochondria-targeted superantioxidant SkQ1. 47 Dramatic effects on various eye disorders from dry eye to glaucoma and on wound healing are documented with a recent report showing that SkQ1-treated mice live significantly longer (335 vs. 290 days). SkQ1 also delays an AD-like phenotype in a rat model of AD. 48 Shabalina et al. achieved antiaging and apparent neuroprotective effects by delivering a potent antioxidant to the mitochondrial inner space to reduce ROS. In contrast, J147 appears to increase ROS. Together these suggest that ROS may either play a dual role in neuroprotection depending on context.
J147 inhibits the ATP synthase only about 20%, and the superoxide increases do not seem all that great (the measurement appears to be mitochondria specific, as it uses fluorescent indicator MitoSox M36008 from molecular probes). The differences seem modest and the apparent contradiction of inhibiting the synthase while simultaneously increasing ATP levels and in generating more ROS but preventing age-related pathologies are not as great as they appear at first glance. Clearly, although J147 is binding to the alpha subunit of ATP synthase, it appears to be doing far more than just inhibiting the synthase. It may not be so much an inhibitor as altering the allosteric activity of the alpha subunit of ATP synthase. Perhaps some of the other J147 targets are important in this regard, such as the three other proteins involved in mTP that Goldberg detected in their pull-down, but not DART assays.
Improvements and Future Directions
Based on the demonstrated neurogenic potential of J147 in mouse models, the Schubert laboratory developed a human neuronal stem cell screening assay to optimize derivatives of J147 for human neurogenesis. The best derivative of J147, CAD-031, maintains the neuroprotective and memory-enhancing properties of J147, yet is more active in the human neural stem cell assays. 13 Which, if any, of these molecules is heading into human clinical trials remains an open question.
A talented Chinese medicinal chemistry group headed by Professor Yuqiang Wang (Jinan University, Guangzhou, PRC) have published and patented a promising compound based on the J147 story. A lead candidate T-006 in which the methoxyphenyl group of J147 was replaced by tetramethylpyrazine has been selected for preclinical development. 49
The ability of J147 to delay epigenic drift, a potentially critical hallmark of aging, is significant and deserves exploration in subsequent experiments. Such studies may lead to greater understanding of the mechanisms that underlie genetic drift.
J147 and related molecules impact many of the pathways contributing to neurodegenerative conditions. For example, Daugherty et al. reported that J147 slowed many of the markers of diabetic neuropathy in the streptozotocin-induced mouse model of type 1 diabetes. 50 In any event, significant follow-up in normally aging mice is required to confirm the antiaging of J147 effects in a mammalian system. Although J147 does demonstrate neuroprotective efficacy in a mouse model of AD, several other mouse models of AD should be investigated as well, including tauopathy models, because of the limited ability of mouse AD models to predict efficacy in humans. If such studies are performed, it would be very interesting indeed to compare J147 with its parental compound curcumin simultaneously in the same model system. In general, the multitargeting curcumin tends to inhibit ROS unlike J147, is neuroprotective in AD mouse models, 7 and also extends lifespan in flies, 51 –53 and in addition extends C. elegans lifespan, 54 which has not yet been reported for J147. Apparently curcumin is not as strongly neuroprotective as J147, 11 although a direct comparison in animal models of AD is warranted. It is important to note that there is opinion suggesting that despite the apparent problems with curcumin, there are enough statistically significant data to not dismiss its potential clinical potential. 55 In any case, translation of this work into human clinical trials for AD is eagerly awaited.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
*