
Abstract
The inherited polyglutamine (polyQ) expansion diseases are characterized by progressive accumulation of aggregation-prone polyQ proteins, which may provoke proteostasis imbalance and result in significant neurotoxicity. Using polyQ transgenic Caenorhabditis elegans models, we find that Kai-Xin-San (KXS), a well-known herbal formula traditionally used to treat mental disorders in China, can alleviate polyQ-mediated neuronal death and associated chemosensory deficiency. Intriguingly, KXS does not reduce polyQ aggregation in vitro as demonstrated by Thioflavin-T test, but does inhibit polyQ aggregation in C. elegans models, indicating an indirect aggregation-inhibitory mechanism. Further investigation reveals that KXS can modulate two key arms of the protein quality control system, that is, heat shock response and autophagy, to clear polyQ aggregates, but has little effect on proteasome activity. In addition, KXS is able to reduce oxidative stress, which is involved in proteostasis and neurodegeneration, but has no effect on life span or dietary restriction response. To examine potential interaction of the four component herbs of KXS, a dissection strategy was used to study the effects of differential herbal combinations in C. elegans polyQ models. While the four herbs do contribute additively to KXS function, Panax ginseng is found to be the most effective constituent. Taken together, these findings not only demonstrate the neuroprotective ability of KXS but also suggest its potential as a proteostasis regulator in protein aggregation disorders and provide an insight into the mechanism studies of traditionally used complex prescriptions and their rationality.
Introduction
The group of hereditary neurodegenerative disorders involving polyglutamine (polyQ) is principally driven by the abnormal expansion in specific genes of a trinucleotide CAG repeat, which encodes a glutamine tract. 1 Although different polyQ diseases are clinically distinguished by the particular gene in which CAG expansion occurs, they share similar clinical features due to polyQ-induced progressive neuronal dysfunction and neuronal cell death. Given the devastating consequences of polyQ diseases, the development of effective therapeutic strategies represents an urgent social and medical need.
Studies over the last few decades suggest that many neurodegenerative diseases are associated with dysregulation of the proteostasis network. 2,3 In normal cells, a combination of quality control systems and signal pathways is used to maintain cellular proteostasis. 3 –5 However, if this proteostasis network is disrupted due to excessive misfolded and aggregated proteins, aging, or metabolic/environmental stresses, it can exacerbate protein aggregation diseases. 2 For example, overexpression of an expanded polyQ tract (≥36 repeats) leads to progressive disruption of proteostasis and cellular dysfunction. 6 Therefore, pharmacological modulation of the proteostasis network, including reduction of misfolded and aggregated proteins, as well as alleviation of external and internal stresses, could be a useful strategy to delay pathogenic protein-mediated disease progression. For instance, autophagy is a fundamental mechanism in the proteostasis network to clear misfolded proteins, and downregulation of ErbB2 by its inhibitor CL-387785 can activate autophagy-mediated clearance of amyloid precursor protein C-terminal fragments and thus improve the cognitive functions in APP/presenilin-1 transgenic mice. 7
Since therapeutic benefit of clinical drugs against neurodegenerative diseases is still limited, increasing attention has been directed to exploring medicinal herbs with neuroprotective effects. In fact, traditional Chinese medicine has been widely used for centuries to treat mental and neurological disorders, such as amnesia, melancholia, anxiety, and uneasiness of mind. 8 In particular, formulae consisting of several herbs are important in traditional clinical practice because the combinations are believed to exhibit synergistic effects and reduce toxic effects of single components. 9,10 Kai-Xin-San (KXS), for instance, is such a classic formula first recorded in Bei-Ji-Qian-Jin-Yao-Fang, a medical encyclopedia by the renowned medical expert Sun Simiao of Tang Dynasty. 11 KXS is relatively simple and contains four component herbs, that is, Acorus gramineus (AG), Polygala tenuifolia (PT), Panax ginseng (PG), and Wolfiporia cocos (WC). This formula and its analogous formulae have been traditionally used to treat mental disorders, especially amnesia and depression. 11 Recent studies have also confirmed its neuroprotective effects on depression, chronic fatigue syndrome, and Alzheimer's disease. 12 –15 However, the potential of KXS against polyQ-induced neurodegeneration has not been explored.
The present study aims to investigate the protective effect of KXS against polyQ neurotoxicity using transgenic Caenorhabditis elegans models and, in particular, to elucidate its mechanisms related to proteostasis imbalance caused by polyQ aggregation. In addition, the four component herbs were also compared for their in vivo neuroprotective effects in the nematode models using a dissection approach.
Materials and Methods
Preparation of KXS and single herb extracts
The herbs, that is, A. gramineus, P. tenuifolia, P. ginseng, and W. cocos, which were produced in Sichuan, Shanxi, Jilin, and Yunnan, respectively, were purchased from Beijing Tongrentang Co., Ltd. (Beijing, China). Dried herbal materials (a mixture of 6 g AG, 6 g PT, 9 g PG, and 9 g WC following the mass ratio of a historical KXS record 11 ) were boiled twice for 1 hour each in 300 mL water. The aqueous extracts were then combined and concentrated to 300 mg/mL (weight of original herbs/volume of final solution). The extraction of individual herbs was performed in the same manner.
Nematode and bacterial strains
The C. elegans strains used in this study include N2 (wild type), HA759 {pqe-1(rt13)III; rtIs11 V[osm-10p::gfp+osm-10p::HtnQ150+dpy-20(+)]}, AM141 {rmIs133[unc-54p::Q40:: YFP]}, CL2070 {dvIs70[hsp-16.2p::gfp+rol-6(su1006)]}, and DA2123 {adIs2122[lgg-1p::gfp:: lgg-1+rol-6(su1006)]}. All C. elegans strains and Escherichia coli OP50 were obtained from the Caenorhabditis Genetics Center (University of Minnesota, Minneapolis, MN) and maintained under standard conditions. Synchronization of nematodes was performed using the standard alkaline hypochlorite method as described. 16 Briefly, gravid adults were lysed in freshly prepared bleaching solution (1% NaOCl/0.5 M NaOH), followed by a quick centrifugation to collect synchronized eggs. The eggs were washed and incubated in M9 buffer with shaking to obtain synchronous L1 nematodes.
Neuronal survival assay
The HA759 strain was used to estimate polyQ-mediated neurotoxicity as described previously. 17,18 Briefly, 100 synchronized L1 larvae were seeded in liquid medium with E. coli OP50 and indicated concentrations of KXS in a final volume of 100 μL of S medium per well in 96-well plates and incubated with shaking at 15°C for 3 days. The nematodes were then collected and paralyzed with 2% NaN3 on an agarose pad (∼50 animals for each treatment). The viability of ASH neurons was evaluated by the presence or absence of GFP fluorescence using an inverted fluorescence microscope (400 × ; Olympus IX51).
Chemosensory behavior assay
The HA759 strain was used for chemosensory behavior assays as described. 19 In brief, a 9 cm NGM plate was equally designated as A and B regions, and 8 M glycerol solution was added along the border of region A and the midline of the plate. Then 2 μL of 1% butanedione was spotted as an attractant in region A (∼1 cm from the edge) to attract the nematodes, and 2 μL of 200 mM NaN3 was dropped near the attractant to paralyze approaching nematodes. The nematodes were treated with the extract at 15°C for 3 days as above in the neuronal survival assay. Before chemosensory assay, approximately 200–300 nematodes were collected, washed thrice to remove bacteria, and dropped in the middle of region B of the assay plate. After incubation at 23°C for 90 minutes, the nematodes in each region were counted and the chemosensory index was calculated as B/(A + B).
In vivo polyQ aggregation assay
The AM141 strain was used to assess polyQ aggregation in vivo as previously described. 17 Synchronized L1 larvae were added to S. medium containing E. coli suspension (final OD570 of ∼0.5) and the extract and incubated at 20°C for the indicated times. Approximately 50 nematodes in each treatment were harvested each day and mounted on an agarose pad and counted for polyQ40::YFP aggregates using an Olympus IX51 microscope (100 × ).
In vitro polyQ aggregation assay
Thioflavin-T (ThT) fluorescence assay was used to quantify polyQ aggregates in vitro as described. 20 Before the test, the GST tag of the polyQ40-GST fusion protein was removed by digestion with trypsin, which was inactivated by boiling. After centrifugation, the supernatant was collected and lyophilized. The polyQ40 powder was then dissolved in a mixture solution of 1,1,1,3,3,3-hexafluoro-2-propanol and trifluoroacetic acid (1:1) to a concentration of 2 mg/mL and treated overnight at room temperature. The polyQ peptides were then lyophilized, dissolved in 10 mM Tris-HCl (pH 7.4) to a final concentration of 20 μM, and incubated with KXS extract at 37°C. At the indicated time points, 10 μL of polyQ40 solution was mixed with 10 μL of 500 mM glycine-NaOH (pH 7.4) and 80 μL of 100 μM ThT (Sigma, St. Louis, MO) in a black 96-well plate. The ThT fluorescence was measured using a fluorescence microplate reader with an excitation of 450 nm and an emission of 580 nm (Fluoroskan Ascent FL; Thermo Scientific).
Quantitative real-time polymerase chain reaction
Approximately 1500 synchronized L1 nematode larvae were added to 3 mL of S. medium containing E. coli OP50 (final OD570 of ∼0.5) and KXS extract and incubated for 3 days. Total RNA was isolated using TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol, and complementary DNA (cDNA) was synthesized by reverse transcription. Real-time polymerase chain reaction (PCR) was performed using SYBR Green I reagent (Bio-Rad, Hercules, CA) with a MyiQ™2 real time detection system (Bio-Rad) as previously described. 21 The primers used are listed in Table 1.
Primers Used for Real-Time Polymerase Chain Reaction
Ben-Zvi et al. 50
Image quantification of HSP16.2 expression
To visualize HSP16.2 expression level, C. elegans strain CL2070 was used as described previously. 22 Synchronized L1 nematodes were added to an E. coli OP50 suspension (final OD570 of ∼0.5) containing KXS extract in a 96-well plate and incubated at 20°C for 3 days. The nematodes were then immobilized and photographed under an ImageXpress Micro system (Molecular Devices, Sunnyvale, CA). For quantification of fluorescence, the images of head and pharyngeal regions from 15 randomly selected nematodes were cut, black-to-white inverted, and measured for mean pixel density using ImageJ software (National Institutes of Health).
Determination of 26S proteasome activity
Assessment of 26S proteasome activity was performed using fluorogenic peptidases as described. 23 Approximately 2000 nematodes per group were treated as described above for the neuronal survival assay and then lysed in the proteasome activity assay buffer (50 mM Tris-HCl, pH 7.5; 250 mM sucrose; 5 mM MgCl2; 0.5 mM EDTA; 2 mM ATP; and 1 mM dithiothreitol) with a glass homogenizer. After centrifugation, 24 μg of the lysate was transferred into a 96-well plate and incubated with the specific fluorogenic substrate (50 μM). The peptides Suc-Leu-Leu-Val-Tyr-R110, Z-Leu-Leu-Glu-R110, and Ac-Arg-Leu-Arg-AMC (Enzo Life Sciences, Lausen, Switzerland) were used to determine chymotrypsin-like, caspase-like, and trypsin-like activities of the proteasome, respectively. Fluorescence (excitation 490 nm and emission 520 nm for chymotrypsin-like and caspase-like activities and excitation 355 nm and emission 460 nm for trypsin-like activity) was measured using a Thermo fluorescence microplate reader every 5 minutes for 1 hour at 25°C.
Quantification of GFP::LGG-1 expression
Assay of GFP::LGG-1 expression was performed using C. elegans strain DA2123 as described previously. 24 Synchronized L1 nematodes treated with KXS extract were incubated at 20°C for 40 hours. The nematodes were then collected, paralyzed with 2% NaN3, mounted on an agarose pad, and photographed under the ImageXpress Micro system. Fifteen nematodes from each treatment were randomly chosen, and GFP-positive puncta were counted in same-sized area per nematode. For starvation treatment, nematodes at L3 stage were washed to remove food and then incubated without food at 20°C for 10 hours.
Lifespan assay
Liquid culture was used to determine C. elegans lifespan as described. 25 Age-synchronized nematodes in 96-well plates (∼10 nematodes per well in 100 μL and 10 wells for each sample) were grown at 20°C to young adult stage in S. medium supplemented with E. coli OP50 (final OD570 of ∼0.5) and 100 μg/mL ampicillin. Then 75 μg/mL 5-fluoro-2-deoxyuridine (final concentration) was added to block reproduction. After incubation at 20°C for 24 hours, the nematodes were treated with KXS extract. Day 0 of the lifespan assay represents the day of adding KXS extract at the beginning of adulthood. The numbers of live and dead nematodes were scored every 2 days under a microscope based on their movement.
Determination of pharyngeal pumping rate
Pharyngeal pumping assay was performed as described. 26 Synchronized L1 nematode larvae were treated with KXS extract in 96-well plates (10 nematodes per well and 10 wells per sample) as above for 3 days at 20°C. Approximately 50 nematodes in each group were transferred to 5 cm solid NGM plates seeded with E. coli OP50 and KXS. The plates were incubated at 20°C for 30 minutes and then placed under a microscope for 30 seconds to acclimate the nematodes. Pharyngeal pumping rate of the nematodes was scored by counting the contractions of the pharyngeal bulb for 10 seconds under the microscope at 100 × total magnification.
Assessment of reactive oxygen species level
Intracellular level of reactive oxygen species (ROS) was determined using 2,7-dichlorofluorescein diacetate (DCFH-DA; Sigma) as previously described. 27 Briefly, after treatment as described above for the neuronal survival assay, nematodes were harvested, washed thrice with phosphate buffered saline (PBS) to remove bacteria, and transferred in PBS to a glass homogenizer. After homogenization and centrifugation, the protein concentration of the lysate was measured by BCA method. Then, 20 μg of lysate was incubated with 50 μM DCFH-DA in a 96-well plate at 37°C for the indicated times. DCFH-DA can diffuse into cells and be hydrolyzed by intracellular esterases to DCFH, which can be rapidly converted by ROS to highly fluorescent 2,7-dichlorofluorescein (DCF). Therefore, the DCF fluorescence, representing ROS level, was measured every 5 minutes for 1 hour at 25°C using the Thermo fluorescence microplate reader with an excitation of 485 nm and an emission of 535 nm and quantified as fluorescence intensity per mg of proteins. The fluorescence levels at a specific time point were analyzed using the Student's t-test, and the difference was considered significant if the p-value was <0.05.
Oxidative stress survival assay
The oxidative stress tolerance of C. elegans was evaluated using paraquat (Sigma), a potent ROS-generating herbicide, as described previously. 28 Approximately 100 synchronized young adult nematodes were grown as described above for the lifespan assay and treated with KXS at 20°C for 4 days. The nematodes were then treated with 0.1 M paraquat, and the numbers of live and dead nematodes were scored microscopically every 12 hours based on their movement.
Statistical analysis
One-way analysis of variance or Student's t-test was performed for statistical analysis, and the differences were considered significant if p < 0.05. For C. elegans lifespan and survival assays, the data were analyzed by the Kaplan–Meier method, and the statistical significance was determined by Peto's log-rank test.
Results
Alleviation of polyQ neurotoxicity and behavioral dysfunction by KXS
To test whether KXS exerts a neuroprotective effect on polyQ disorders, we examined the effect of KXS extract on C. elegans HA759, which expresses polyQ150 derived from human huntingtin, as well as GFP and polyQ toxicity enhancer pqe-1. The presence of GFP fluorescence is used as an indicator of ASH survival, while progressive loss of fluorescence represents neurodegeneration and neuronal death (Fig. 1A). 17 As shown in Figure 1B, the survival rate of ASH neurons was <40% in the control nematodes after incubation at 15°C for 3 days, indicating a significant neurotoxic effect of polyQ aggregation as reported previously. 17,18 Treatment with KXS, however, increased the survival rate of ASH neurons at doses >20 mg/mL, demonstrating a protective effect of KXS against polyQ-mediated neuronal death.
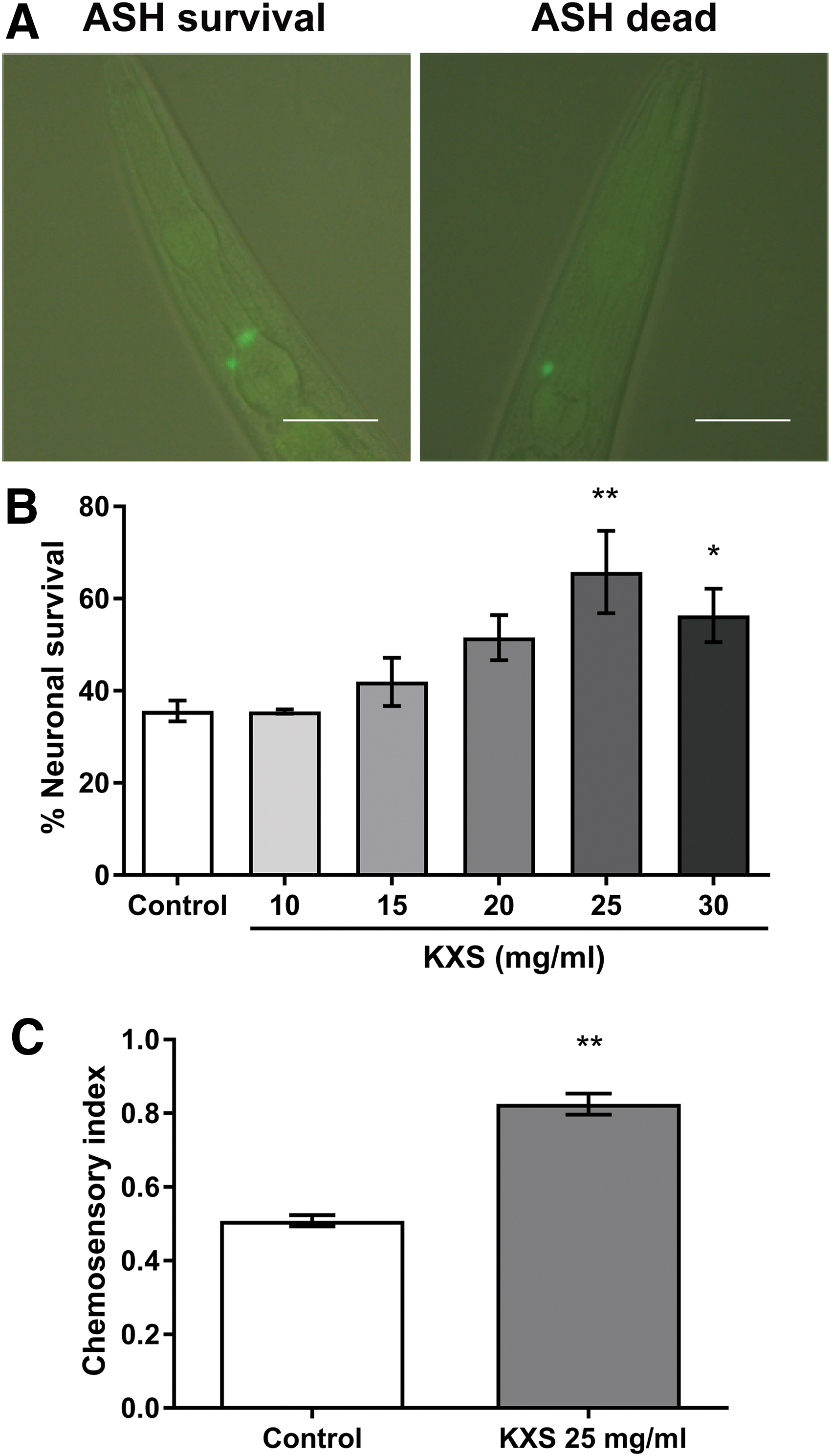
Alleviation of polyQ150-mediated neuronal death by KXS in Caenorhabditis elegans HA759.
Since behavioral impairment is a major clinical symptom in polyQ disorders, we further tested the effect of KXS on the chemosensory behavior of HA759 nematodes. 19 The nematodes were first treated with or without KXS at 15°C for 3 days as above and then incubated at 23°C for 90 minutes in the chemosensory assay; HA759 nematodes were far less active and few would move across the glycerol barrier line when they were incubated at 15°C for 90 minutes. As shown in Figure 1C, after 90 minutes at 23°C, approximately half of the control nematodes failed to avoid the glycerol barrier due to the loss of ASH neurons, which are required for avoidance of hyperosmotic chemicals in C. elegans. 29 Interestingly, when the nematodes were treated with 25 mg/mL KXS, their avoidance capability was significantly improved. Collectively, these results suggest that KXS is capable of protecting C. elegans against polyQ-induced neurotoxicity.
Inhibition of in vivo, but not in vitro, polyQ aggregation by KXS
Since reduction in the number of abnormal protein aggregates is known to help maintain proteostasis and suppress proteotoxicity, 30 we used the transgenic C. elegans strain AM141 to investigate the effect of KXS on polyQ aggregation. As AM141 nematodes grow, the polyQ40::YFP expressed in their body wall muscle cells will gradually aggregate and form fluorescent foci, which are bright and easy to distinguish in early stages (Fig. 2A). In control nematodes, the average number of polyQ40::YFP aggregates was 11 and 35 after 48 and 72 hours, respectively (Fig. 2B). When the nematodes were treated with 25 mg/mL KXS, however, the average numbers of aggregates were decreased to 6 and 29 at 48 and 72 hours, respectively, suggesting that KXS can inhibit polyQ aggregation in vivo. Then we examined whether KXS reduces aggregation through direct interaction with polyQ proteins. Using an in vitro ThT fluorescence method, we found that polyQ proteins aggregated over time as ThT-positive fibrillar structures regardless of whether KXS was present or not; in the same experiment, EGCG, a potent anti-aggregation compound able to interact with amyloid proteins, 31 decreased ThT fluorescence (p < 0.05) (Fig. 2C). Together, these results indicate that KXS is likely to reduce polyQ aggregate formation in vivo by an indirect mechanism.
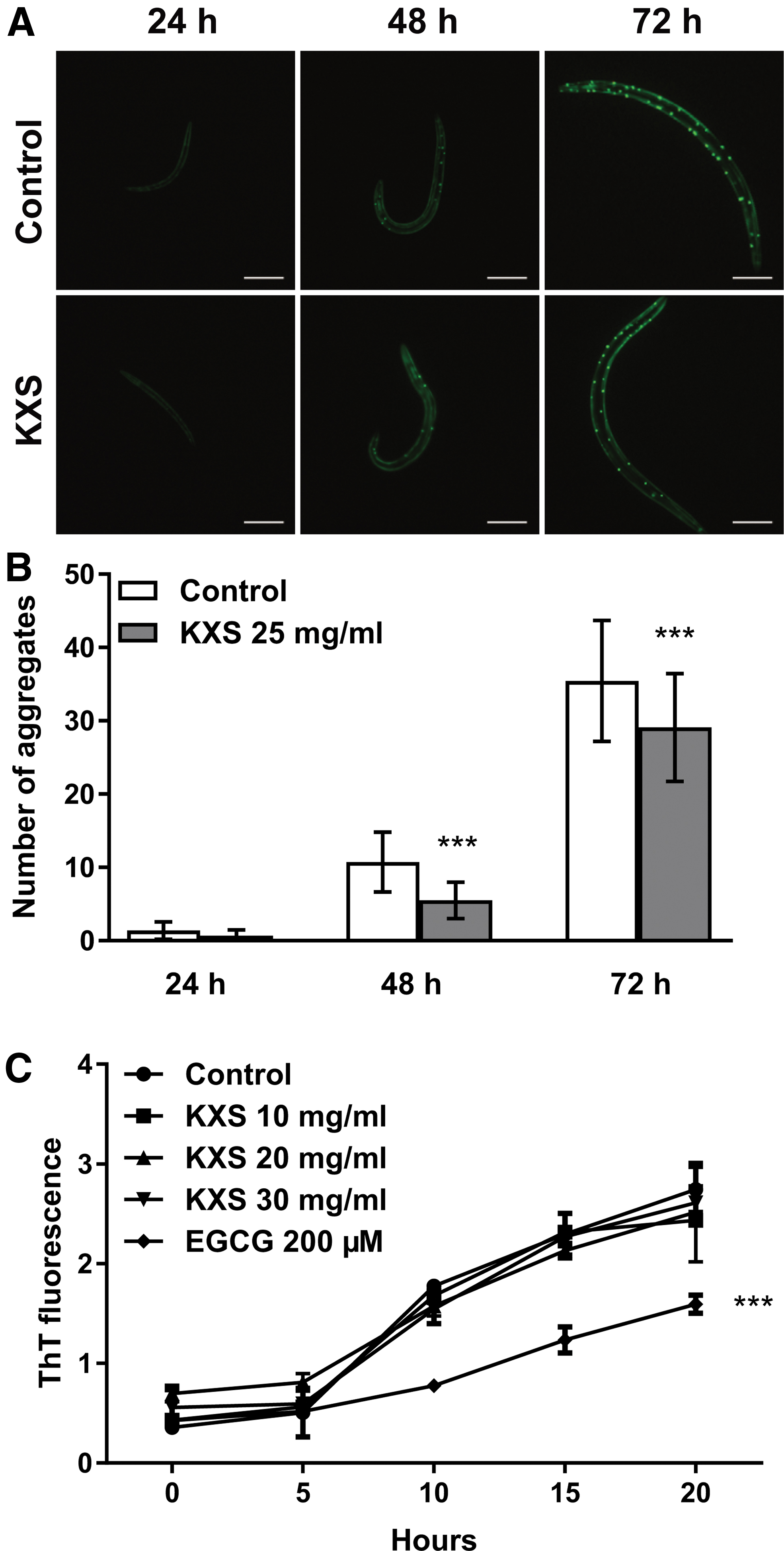
Prevention of polyQ aggregation by KXS in vivo but not in vitro.
Regulation of heat shock protein expression by KXS
As KXS probably inhibits polyQ aggregation indirectly, we then asked whether it functions through the cellular protein quality control system. First, we investigated its effect on the expression of a set of heat shock proteins (HSPs), the crucial machinery in protein quality control. 32 Real-time PCR analysis revealed that, after KXS treatment, hsp16.2 expression was strongly upregulated in wild type (N2), as well as transgenic polyQ nematodes HA759 and AM141 (Fig. 3A–C). However, the expression of the HSP70 (hsp1) was not changed significantly in all three nematode strains. HSP90 gene (daf-21) expression was downregulated in HA759 nematodes, but was not changed significantly in AM141 and N2. These results strongly suggest that KXS acts as an inducer for HSP16.2. Therefore, we used a transgenic strain CL2070 carrying a hsp-16.2p::gfp reporter gene to confirm this finding. As shown in Figure 3D and E, the mean pixel density of GFP fluorescence increased significantly after heat shock or treatment with 25 mg/mL KXS, demonstrating a positive effect of KXS on hsp16.2 gene expression.

Modulation of HSPs in C. elegans by KXS.
Involvement of autophagy, but not proteasome, in the inhibition of polyQ aggregation by KXS
Apart from molecular chaperones, cells also use the ubiquitin-proteasome system and the autophagy-lysosome pathway to remove misfolded or aggregated proteins. 33 We therefore first examined the effect of KXS on 26S proteasome activity using fluorogenic peptide substrates specific for its chymotrypsin-like, caspase-like, and trypsin-like protease activities, which were monitored by increase in fluorescence over a 60-minute time course. 23 As shown in Supplementary Figure S1, the fluorescence in all the samples of nematodes treated either with or without 25 mg/mL KXS was increased in the 60-minute period, while that of the reactions without nematode lysates stayed unchanged during the 60-minute reaction (data not shown). KXS, however, showed no effect on the activities of the three proteases in all nematode strains tested (N2, HA759, and AM141) as the fluorescence remained almost at the same level between respective KXS-treated and nontreated nematodes at all the time points of reaction, suggesting that KXS does not regulate the proteasome. Then we tested the expression of key autophagy (otherwise known as macroautophagy) gene lgg-1 (the nematode ortholog of mammalian LC3) to study the influence of KXS on this clearance pathway. As shown in Figure 4A, the expression of lgg-1 was upregulated by KXS in all N2, HA759, and AM141 strains. Furthermore, we used the transgenic DA2123 strain, expressing the gfp::lgg-1 reporter gene, to confirm the effect of KXS on the lgg-1 gene, and found that 25 mg/mL KXS also resulted in an increase in the mean number of GFP-positive puncta per area in DA2123 nematodes (Fig. 4B, C). Since lgg-1 is a widely accepted marker for autophagy in C. elegans, 34 these results demonstrate that KXS is able to induce autophagy.
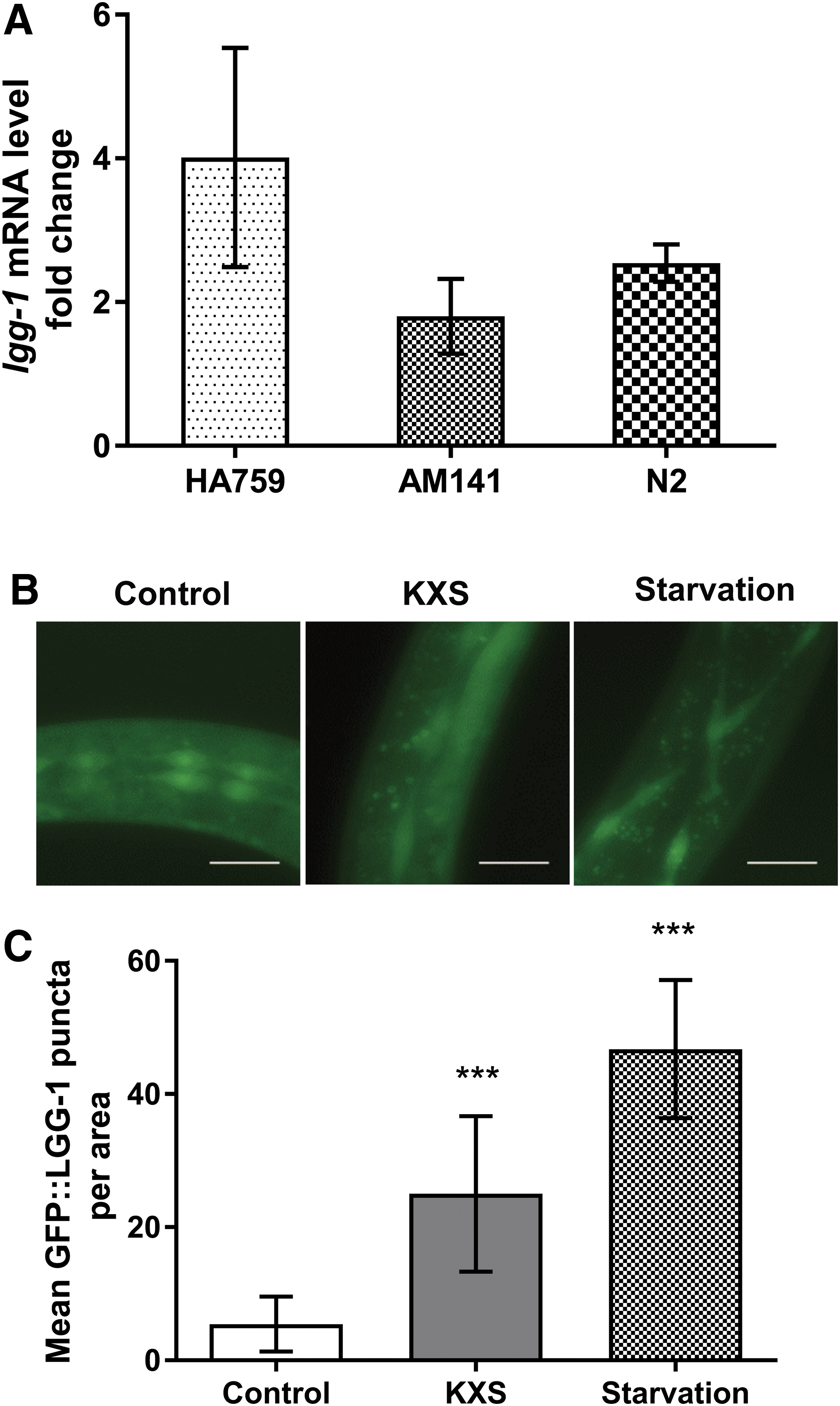
Induction of autophagy by KXS in C. elegans.
In summary, two key arms of the protein quality system, that is, the heat shock response and autophagy, are involved in KXS-mediated reduction of polyQ aggregation in C. elegans.
Association of oxidative stress, but not aging stress and dietary restriction, with the neuroprotective effect of KXS
Signal pathways that respond to stresses such as aging, dietary restriction, and oxidative stress, are known to impact the proteostasis network. 5 As aging plays an important role in late-onset neurodegeneration, we first determined the effect of KXS on C. elegans lifespan and found that treatment with 25 mg/mL KXS did not affect the lifespan of either wild-type or transgenic polyQ nematodes (Supplementary Fig. S2). Then we investigated the influence of KXS on pharyngeal pumping rate of the nematodes as decreased pharyngeal pumping may cause reduced food intake and induce dietary restriction, an intervention that retards aging and suppresses protein aggregation, and found that the pharyngeal pumping rate of KXS-treated nematodes was essentially unchanged compared to untreated animals (Supplementary Fig. S3). Oxidative stress has been implicated in the pathophysiology of many neurodegenerative diseases, 35 and thus, we further examined the ROS level in nematodes using a DCFH-DA fluorescence method. As shown in Figure 5A–C, the ROS level was significantly reduced by KXS treatment in N2, HA759, and AM141 strains. Next, a paraquat assay was used to measure the effect of KXS on survival rate of nematodes under conditions of oxidative stress. As shown in Figure 5D, treatment with KXS increased the survival time of N2 nematodes exposed to paraquat. Collectively, these findings indicate that KXS does not delay aging or induce a response similar to that caused by dietary restriction, but instead confers protection against polyQ toxicity through an antioxidant mechanism.
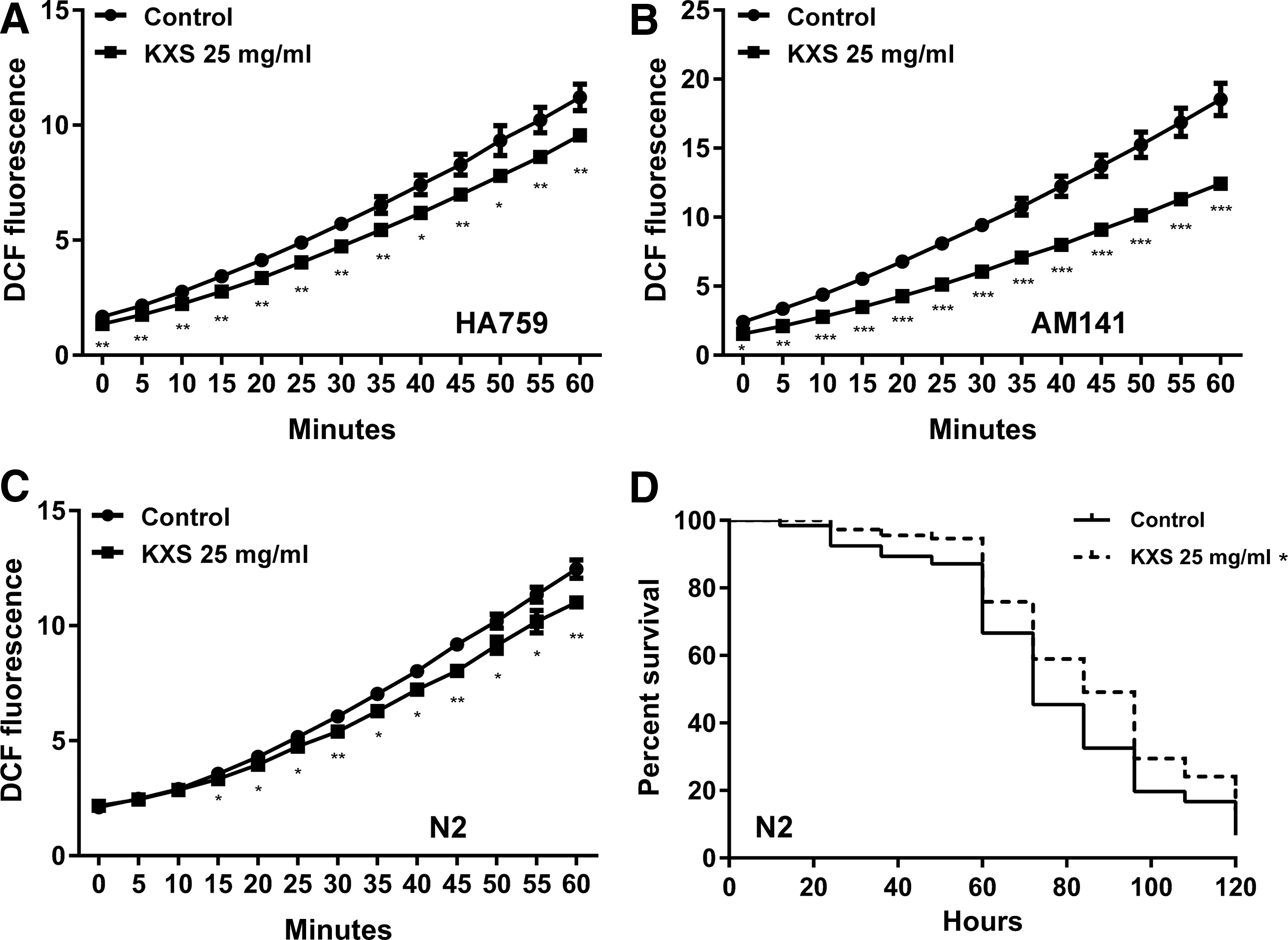
Reduction of oxidative stress by KXS in C. elegans.
Additive effect of KXS component herbs against polyQ neurotoxicity
To study potential interaction of the component herbs of KXS, we used a “dissection” approach, which is an effective strategy for understanding the design of traditional formulae, 36 with equivalent concentrations of individual extracts in the formula. As shown in Figure 6A, treatment with A. gramineus extract (AG; 5 mg/mL), P. ginseng extract (PG; 7.5 mg/mL), and KXS (25 mg/mL) showed protection against polyQ-mediated neurotoxicity in HA759 nematodes, while no significant effect was detected with P. tenuifolia extract (PT; 5 mg/mL) and W. cocos extract (WC; 7.5 mg/mL) treatment, suggesting that AG and PG play important neuroprotective roles in the KXS formula. Then, each component herb was excluded from the KXS formula, one at a time, and the neuroprotective effects of the dissected formulae were investigated. Interestingly, removing PG leads to a relatively weak but still effective protection compared to the full KXS formula, while removing the other herbs individually does not affect the overall protective effect of the formula (Fig. 6B). Taken together, these results suggest that at least some of the single herbs in the KXS formula act in an additive, or perhaps synergistic, way; P. ginseng may be the principal component, and other herbs may represent adjuvant components.
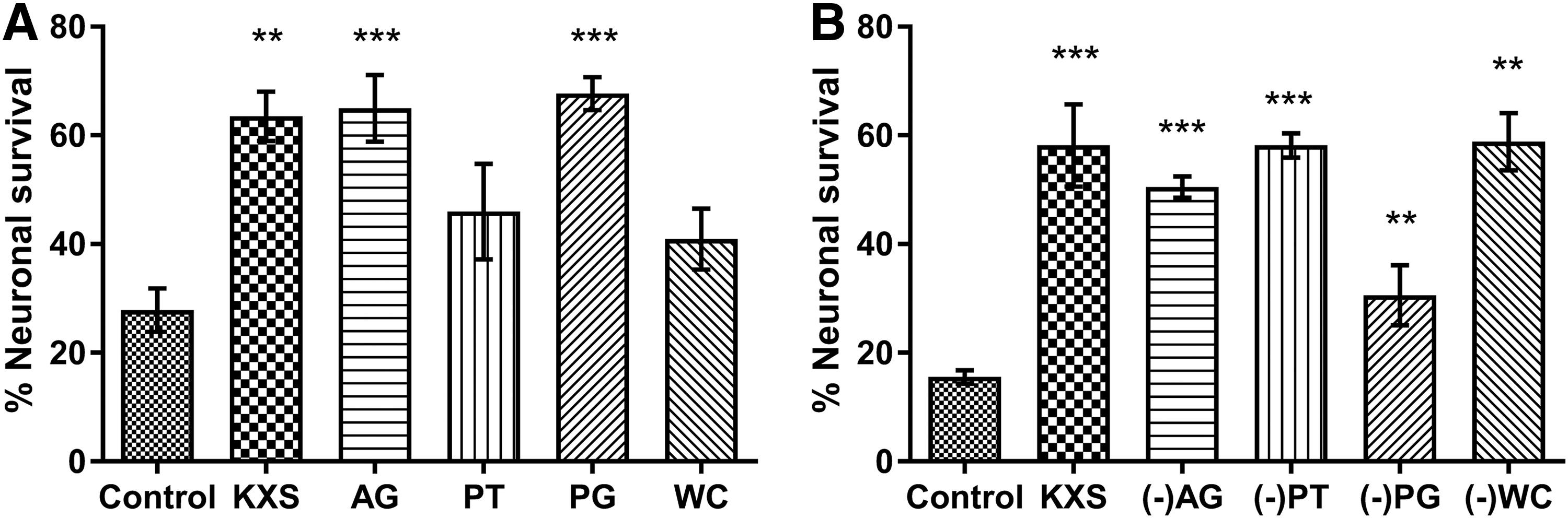
Neuroprotective effect of KXS and its component herbs in C. elegans.
Discussion
Pathogenic protein aggregates play a key role in proteostasis imbalance and neurodegeneration, and great efforts have been made to seek therapeutics capable of suppressing proteotoxicity and delaying disease onset. 37 In this study, we find that KXS, a traditional formula for the treatment of mental disorders, can significantly inhibit the neurotoxicity mediated by polyQ aggregation; for example, KXS ameliorates the impairment by polyQ aggregation of avoidance behavior in C. elegans (Fig. 1). These findings not only demonstrate the neuroprotective ability of KXS but also suggest that it may act as a proteostasis regulator in protein aggregation disorders.
A number of studies have shown that modulation of the protein quality control system, the central component of the proteostasis network, is an effective strategy for reducing proteotoxicity in many model organisms. 38 Using a transgenic C. elegans strain expressing polyQ40 tracts, KXS was found to inhibit polyQ aggregation (Fig. 2), suggesting an ability to delay disease onset. However, further analysis showed that KXS had no influence on polyQ oligomer or fibril formation in vitro. This is similar to the effect of another traditional formula, Liuwei Dihuang, which alleviates amyloid Aβ-induced paralysis without directly affecting Aβ aggregation. 39 These results suggest that KXS may act through the cellular protein quality control system instead of targeting the aggregating protein species itself.
HSPs, which have several roles in protein biosynthesis, such as maintaining proteins in soluble stable conformations and refolding misfolded proteins, are key actors in the proteostasis network. 32 For example, both genetic and pharmacologic approaches targeting HSP70 can promote polyQ androgen receptor clearance and alleviate neurotoxicity in a Drosophila model. 40 Since activation of the molecular chaperone machinery provides a promising strategy for reducing protein aggregates, we tested the effect of KXS on gene expression of HSPs and found that KXS can significantly upregulate hsp16.2 expression in wild-type N2 and two polyQ strains (Fig. 3). This effect was confirmed in transgenic strain CL2070, which carries an hsp-16.2p::gfp fusion gene. In summary, our results uncover an important role of hsp16.2 in the protective effect of KXS.
Under normal conditions, protein degradation is critical for the removal of protein aggregates and damaged proteins, but excessive levels of aggregation may compromise cellular degradation systems, leading to a chronic imbalance in proteostasis. 41 Therefore, enhancement of the cellular degradation machinery provides an effective means of improving proteostasis and may delay disease progression. While KXS has little effect on proteasome activity, it can modulate the expression of the autophagy gene lgg-1, a marker gene for autophagosomes. Induction of autophagy by KXS was confirmed in the gfp::lgg-1 transgenic strain (Fig. 4). Since inactivation of autophagy genes has been demonstrated to accelerate the accumulation of polyQ aggregates in C. elegans, 42 these findings suggest a positive connection between the regulation of autophagy by KXS and its neuroprotective effect.
Signal pathways that respond to stresses such as aging, dietary restriction, and oxidative stress, have been shown to impact the proteostasis network and limit the progression of neurodegeneration. 5 For example, dietary restriction can also suppress polyQ and Aβ proteotoxicity and increase longevity in C. elegans. 43 Unexpectedly, our data suggest that KXS, at a concentration of 25 mg/mL, did not alter the lifespan or pharyngeal pumping in wild type and polyQ strains, suggesting that its action is independent of the longevity and dietary restriction mechanisms, such as the IIS pathway. 17 In contrast, KXS does offer protection against oxidative stress, another detrimental factor in polyQ pathogenesis. 35 Antioxidants may improve proteostasis and reduce proteotoxicity, for example, a formula containing certain common antioxidants can significantly reduce oxidative stress and improve mental state in AD patients. 44 Our study reveals that KXS is able to decrease ROS level and extend the survival time under paraquat stress in C. elegans; this significantly contributes to the reduction of polyQ-mediated neurotoxicity. Previous studies also showed that KXS can improve memory deficiency induced by hindlimb suspension majorly related to its antioxidant activities. 45 These findings are consistent with a beneficial effect of KXS on cellular antioxidant defense systems. In addition, given the positive relationship between polyQ and oxidative stress, clearance of polyQ aggregates by KXS may also alleviate oxidative damage.
Although many studies have demonstrated a therapeutic role for traditional formulae in treating neurodegenerative diseases, in most cases the active components remain to be identified. In the present study, we dissected the KXS formula to explore the active components and showed that, in C. elegans at least, P. ginseng has the most pronounced neuroprotective effect of the KXS constituents. This is in line with ancient records, in which ginseng was originally described as the principal component of KXS. Moreover, we found that A. gramineus, a neuroprotective herb well known for its volatile oil and asarone, 46 also protected ASH neurons against polyQ toxicity (Fig. 6). However, P. tenuifolia and W. cocos did not exhibit beneficial effects in HA759 nematodes (Fig. 6). These findings indicate that P. ginseng makes the major contribution in the neuroprotective effect of KXS. Interestingly, a variety of compounds from P. ginseng and A. gramineus are shown to have protective effects against neurodegenerative diseases. For example, ginsenosides Rb1, Rc, and Rg5 from P. ginseng are capable of protecting medium spiny neurons from glutamate-induced apoptosis in YAC128 HD mouse model. 47 Ginsenosides are also shown to induce autophagy, which helps to explain the reduced Aβ deposits and pathological behaviors in transgenic C. elegans models. 48 Furthermore, β-asarone, a major bioactive component of A. gramineus, can also inhibit neuronal apoptosis through the CaMKII/CREB/Bcl-2 pathway both in vitro and in AβPP/PS1 mice. 49 Therefore, these active compounds of the component herbs may contribute to the protective effect of KXS against polyQ-mediated neurotoxicity.
Footnotes
Author Disclosure Statement
The authors declare no conflicts of interest relevant to this study.
Funding Information
This work was supported by the 111 Project (B17018), Guangdong Yangfan Innovative & Entepreneurial Research Team Program, Special Funds of the Central Finance to Support the Development of Local Universities and Colleges, Guangdong Province Department of Education (2015KGJHZ022 and 2016KQNCX084), Natural Science Foundation of Guangdong Province (2017A030313864), and China Postdoctoral Science Foundation (2019M653300).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.